
Abstract
Indoleamine 2,3-dioxygenase (IDO1) and tryptophan 2,3-dioxygenase (TDO) are two structurally different enzymes that have a different tissue distribution and physiological roles, but both catalyze the conversion of tryptophan to N-formylkynurenine (NFK). IDO1 has been clinically validated as a small-molecule drug target for cancer, while preclinical studies indicate that TDO may be a target for cancer immunotherapy and neurodegenerative disease. We have developed a high-throughput screening assay for IDO1 and TDO based on a novel chemical probe, NFK Green, that reacts specifically with NFK to form a green fluorescent molecule with an excitation wavelength of 400 nm and an emission wavelength of 510 nm. We provide the first side-by-side comparison of a number of published inhibitors of IDO1 and TDO and reveal that the preclinical IDO1 inhibitor Compound 5l shows significant cross-reactivity with TDO, while the relative selectivity of other published inhibitors was confirmed. The suitability for high-throughput screening of the assays was demonstrated by screening a library of 87,000 chemical substances in 384- or 1536-well format. Finally, we demonstrate that the assay can also be used to measure the capacity of cells to metabolize tryptophan and to measure the cellular potency of IDO1 and TDO inhibitors.
Keywords
Introduction
Tryptophan is an essential amino acid that is used for protein synthesis and the production of neurotransmitters that regulate behavior, such as serotonin and melatonin. Tryptophan is also a regulator of the local immune response by T lymphocytes. Aberrant tryptophan catabolism has been implicated in a number of diseases, including cancer, and neurodegenerative diseases. The first and rate-limiting enzymes in the degradation of tryptophan are indoleamine 2,3-dioxygenase (IDO1) (EC 1.13.11.42) and tryptophan 2,3-dioxygenase (TDO) (EC 1.13.11.11). Both enzymes have a different tissue distribution and different physiological roles. IDO1 is broadly expressed, is regulated by interferon-γ (IFN-γ), and acts locally. IDO1 expressed in the placenta protects the fetus from allogeneic rejection. 1 In contrast, TDO is constitutively expressed at high levels in the liver, where it regulates systemic tryptophan levels. 2 Despite catalyzing the same reaction, the two enzymes have little structural similarity.
Many human tumors constitutively express IDO1, 3 and high expression of IDO1 correlates with poor prognosis in colorectal and endometrial cancer.4,5 Expression of IDO1 by immunogenic mouse tumor cells prevented their rejection by preimmunized mice. 3 The effect was accompanied by a lack of accumulation of specific T cells and could be reverted by systemic application of 1-methyl-L-tryptophan (1-MT), a low-potency inhibitor of IDO1.1,6
In addition, 1-MT increased the antitumor activity of various chemotherapeutics and cancer vaccines in mouse tumor models.7,8 INCB024360 is a potent IDO1 inhibitor developed by Incyte Corporation (Wilmington, DE). 9 INCB024360 showed reduction of tumor volume in immunocompetent mice 9 and a dose-dependent reduction of kynurenine levels in patients with cancer in a phase I study. 10 INCB024360 is currently evaluated in phase II clinical trials as a single agent in ovarian cancer and in combination with the CTLA-4 antagonist ipilimumab in metastatic melanoma, clearly showing the potential of IDO1 inhibitors as a treatment option for cancer. 10
Similar to IDO1, expression of TDO by immunogenic mouse tumor cells prevented their rejection by preimmunized mice, and the effect could be reverted with an inhibitor. 11 Moreover, TDO is overexpressed in brain tumors, where it supports tumor growth and immune evasion by maintaining intratumoral kynurenine levels that activate the aryl hydrocarbon receptor and promote the generation of immune-suppressive T cells. 12 Besides cancer, there is evidence that TDO activity contributes to neurodegenerative disease. 13 Patients with Alzheimer disease (AD) have low levels of serum tryptophan and high levels of serum kynurenine, and these changes correlate with the level of cognitive decline. 14 TDO was found to be overexpressed in brain tissue of patients with AD. 15 Brain levels of quinolinic acid, kynurenic acid, and 3-hydroxykynurenine have been reported to be elevated in early phases of Huntington disease, in line with findings in mouse models. 16 Recently, TDO was identified as a metabolic regulator of age-related α-synuclein toxicity in a Caenorhabditis elegans model. 17 Depletion of TDO suppressed toxicity of amyloid-β and polyglutamine and increased life span. The same effect could be obtained by addition of dietary L-tryptophan. 17 These studies support the idea that restoring the balance in tryptophan metabolism may provide therapeutic benefit in patients with neurodegenerative disease.
Despite the substantial genetic and pharmacological evidence of the importance of IDO1 and TDO as small-molecule drug targets, only a handful of different chemical scaffolds have been reported. Most inhibitors reported for TDO contain the same indole pharmacophore.11,18 For IDO1, INCB024360 is advanced to phase II, albeit that its oral bioavailability in humans seems to be hampered by a short half-life (~0.5 h in mice), 9 explaining the high doses, administered twice daily, to patients. 10
The lack of chemical matter is explained by the lack of robust assays for high-throughput screening (HTS). Both IDO1 and TDO are heme-containing enzymes that use oxygen and L-tryptophan to produce N-formylkynurenine (NFK). In activity assays, NFK can be directly detected (e.g., by high-performance liquid chromatography [HPLC] methods or by its intrinsic fluorescence). However, fluorescence detection is not preferable for screening, because the low wavelength absorbance of NFK (i.e., 312 nm) makes direct detection prone to interference and quenching by compounds in screening libraries. Many laboratories have therefore searched for alternative readouts for screening. One often-used approach is based on deformylation of NFK to kynurenine, followed by reaction with 4-(dimethylamino)benzaldehyde (pDMAB).19–21 Unfortunately, pDMAB reacts with all compounds containing primary amines.
21
In one recent alternative assay format, kynurenine was linked to a chemosensor, such as sensor-1, to produce a new fluorophore.
22
NFK can also be directly detected by reaction with piperidine to produce a fluorophore.
23
However, while both these assays have better selectivity than pDMAB,
22
they have one or more steps at extremely acidic pH (pH 1.0) or extremely basic pH (pH 11.0) and high temperature (55–65 °C). Furthermore, piperidine is a toxic chemical and has a nasty smell, hampering its use in large HTS campaigns. Therefore, neither of these assay formats is well suited for HTS. Here, we describe the development and validation of a new assay format. In this assay, NFK is directly converted into a fluorophore under mild conditions, which is detected by fluorescence at long wavelengths (510 nm), where screening compound interference is minimal (see
Materials and Methods
Assay Chemicals
NFK was synthesized according to the procedure described by Todorovski et al. 24 NFK Green was prepared in house and is commercially available at Netherlands Translational Research Center B.V. (NTRC) (www.ntrc.nl). Ascorbic acid was purchased from Acros (cat. no. 352681000; Geel, Belgium), catalase from Sigma Aldrich (cat. no. C40; St. Louis, MO), methylene blue from Janssen Chimica (cat. no. 1267565; Geel, Belgium), and L-tryptophan from Sigma Aldrich (cat. no. T0254).
Inhibitors and Compound Libraries
The IDO1 reference inhibitor NTRC 0820-0 and the TDO reference inhibitors NTRC 0672-0 and NTRC 0989-0 were synthesized in house. NTRC 0820-0 is identical to example 35 from Röhrig et al. 19 NTRC 0672-0 is identical to example 4 from Madge et al. 18 NTRC 0989-0 is identical to example 25 from Dolušić et al. 25 The IDO1 reference inhibitors 1-MT 5 and norharmane 26 were purchased from Sigma. Compound 5l9 and S7111 27 were purchased from, respectively, MedKoo, Inc. (Chapel Hill, NC), and Selleck/Absource GmbH (Munich, Germany). The compound library screened at Pivot Park (Oss, The Netherlands) was composed by selection of a diverse set of 87,000 lead-like compounds from a commercial supplier (Specs, Delft, The Netherlands). Before the compounds were purchased by Pivot Park, all structures were filtered for optimal physical chemical properties and structural alerts. Furthermore, all structures were visually inspected by NTRC chemists to remove remaining promiscuous, nonspecific, or reactive compounds.
Protein Expression
Full-length human IDO1 (Uniprot ID 14902) was expressed with an N-terminal hexahistidine tag. 28 For TDO (Uniprot ID 48775), a fragment comprising amino acids 18 to 388 with a C-terminal hexahistidine tag was used. The proteins were expressed in Escherichia coli grown in medium containing extra hemin and purified by affinity chromatography using Ni-NTA agarose beads (Qiagen, Venlo, The Netherlands) to >95% purity, as determined by polyacrylamide gel electrophoresis and scanning of the gel. The proteins were desalted with a PD10 column (GE Healthcare Bio-Sciences AB, Uppsala, Sweden) and stored at −80 °C in aliquots in buffer containing 10% glycerol (cat. no. 18.469.39; Janssen Chimica). Enzymatic activity was characterized in assays measuring the conversion in time of L-tryptophan (Sigma-Aldrich) by monitoring the increase in UV absorption of NFK at 325 nm. 20 Michaelis-Menten parameters (KM, Trp and kcat) were determined to characterize the enzyme preparations. For IDO1, a KM, Trp of 6 µM and kcat of 4.6 s−1 were measured. For TDO, KM, Trp was 190 µM and kcat was 1.1 s−1. These values are consistent with those from literature.21,24
IDO1 Assay
Compounds were dissolved and diluted in DMSO and finally in IDO1 reaction buffer, consisting of 50 mM NaH2PO4, pH 6.5, supplemented with 0.05% Tween-20 (cat. no. P7949; Sigma Aldrich) and 1% glycerol. A fresh IDO1 solution was prepared from a frozen batch on the day of use. Enzyme solution was kept on ice and used within 3 h to start an assay. All assay components were diluted in IDO1 reaction buffer. Then, 10 µL of compound solution, 20 µL of a 50-nM enzyme solution in phosphate buffer supplemented with 20 mM ascorbic acid, 20 µg/mL catalase, and 20 µM methylene blue were combined in the well of a black 384-well plate (cat. no. 3573; Corning, Corning, NY) using an AquaMax dispenser (Molecular Devices, Sunnyvale, CA) and incubated for 30 min at room temperature. Subsequently, 10 µL of 0.4 mM of the substrate L-tryptophan was added by using a Multidrop dispenser (Thermo Scientific, Waltham, MA), and incubation was continued for 60 min at room temperature. Then, 10 µL of NFK Green was added by a FlexDrop dispenser, the plate was sealed, and the reaction was developed for 4 h at 37 °C. NFK Green was dissolved in DMSO, which stopped the reaction. To determine the production of NFK, the seal was removed and fluorescence was read on an EnVision multimode reader (PerkinElmer, Waltham, MA) (excitation, 400/25; emission, 510/20). For HTS, plates were covered with a lid (cat. no. 3099; Corning) and set in a tissue culture incubator at 37 °C, 95% humidity, and normal atmospheric CO2 (0.03%). Assay quality was determined by signal-to-background (S/B) and calculation of Z′ factor. Eighteen wells of a 384-well plate were used as background or control wells (enzyme reaction mixture without L-tryptophan) and 18 wells to determine the maximum signal (all components of the reaction, including L-tryptophan). The same wells were used as control wells to calculate the Z′ factor. In routine profiling experiments and in HTS, robustness of the assay and stability of assay reagents were further monitored by determining 10-point dose-response curves of a reference inhibitor (NTRC 0820-0). Dose-response curves were fitted to a four-parameter logistics equation in XLfit (ID Business Solutions, Ltd., Guildford, UK). pIC50 values were used to calculate average potencies of the inhibitors and variation, as summarized in Table 1 . pIC50 is –log IC50. For convenience, inhibitory potencies are described as IC50 values in the text and the figure legends. For the article, dose-response curves were redrawn in Prism (GraphPad Software, La Jolla, CA).
Inhibitory Potency a of IDO1 and TDO Reference Inhibitors in Biochemical NFK Green Assays.
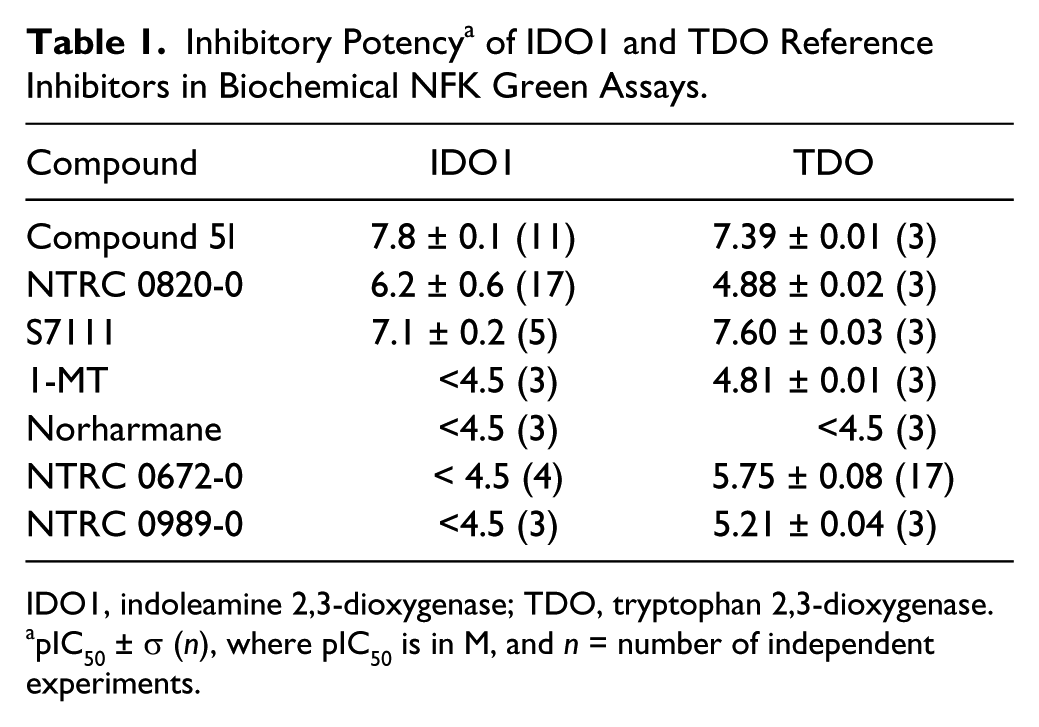
IDO1, indoleamine 2,3-dioxygenase; TDO, tryptophan 2,3-dioxygenase.
pIC50 ± σ (n), where pIC50 is in M, and n = number of independent experiments.
TDO Assay
The biochemical assay to determine NFK production by TDO was as described for IDO1, except that 100 mM NaH2PO4, pH 7.0, supplemented with 0.01% Tween-20 was used, and catalase and methylene blue were omitted from the reaction. The final concentration of ascorbic acid was 100 µM, and 0.2 mM L-tryptophan was used. Concentration of TDO was 50 nM. TDO was more stable than IDO1, and the enzyme solution could be used up to 6 h, when kept on ice, before the start of an assay. Standard reaction time was 15 min. Assay performance was monitored as described above for IDO. Reference TDO inhibitor used in assay optimization was NTRC 0672-0.
NFK Deselection Assay
To deselect screening hits that interfere with NFK Green, assays were carried out in which compounds were mixed with purified NFK in the absence of enzyme. Then, 10 µL of compound solution was combined with 20 µL purified NFK in the well of a black 384-well plate and incubated for 30 min at room temperature. After addition of 10 µL 0.4 mM L-tryptophan, the incubation was continued for 1 h. Next, 10 µL of 5× NFK Green solution was added, the plate was sealed, and the reaction was developed for 4 h at 37 °C. After removal of the seal, fluorescence was read on an EnVision multimode reader.
Cell-Based Assay for TDO
Cell lines were purchased from LGC Standards GmbH (Wesel, Germany). A-172 glioblastoma cells were cultured in RPMI 1640 tissue culture medium (Life Technologies, Bleiswijk, The Netherlands), supplemented with 10 % (v/v) bovine calf serum (BCS). Compounds were dissolved in DMSO and diluted in RPMI 1640. Final DMSO concentration in the assay was 0.4 % (v/v). Eight thousand cells per well in 40 µL were seeded in a black 384-well tissue culture plate (cat. no. 781086) from Greiner Bio-One GmbH (Frickenhausen, Germany) and allowed to adhere by incubation at 37 °C, 95% humidity, and 5% CO2 for 3 h. After addition of 5 µL compound solution and 1 h of incubation, 5 µL of 10 mM L-tryptophan in RPMI 1640 (sterilized by filtration) was added and incubation was continued for 42 h. To determine NFK levels, 10 µL NFK Green was added to each well, and the plate was sealed and incubated for 4 h at 37 °C. Fluorescence was measured on an EnVision multimode reader as described above for the biochemical assays. Possible cellular toxicity of compounds was determined with ATPlite 1Step assay (Perkin Elmer, Groningen, The Netherlands), performed in the same well.
Cell-Based Assay for IDO1
MDA-MB-231 breast cancer cells were cultured in RPMI 1640, supplemented with 10% BCS. Compounds were dissolved in DMSO and diluted in RPMI 1640. Final DMSO concentration in the assay was 0.4% (v/v). Eight thousand cells per well in 35 µL were seeded in a black 384-well tissue plate (cat. no. 781086; Greiner) in RPMI 1640 plus 10% BCS and allowed to adhere by incubation at 37 °C, 95% humidity, and 5% CO2 overnight. Then, 5 µL of compound solution was added to the cells 1 h prior to stimulation with 5 µL of 500 ng/mL IFN-γ (R&D Systems, Minneapolis, MN) in RPMI 1640 medium. At the same time, 5 µL of sterile 0.5 mM L-tryptophan in 20 mM Hepes (pH 7.4) was added, making the final L-tryptophan concentration in the medium 75 µM. Incubation was continued for 48 h. To determine NFK levels, 10 µL NFK Green was added to each well, and the plate was sealed and incubated for 4 h at 37 °C. Fluorescence was measured on an EnVision multimode reader as described above for the biochemical assays.
Results
NFK Green
A NFK-specific chemical probe was identified by screening a proprietary library of chemicals for compounds that specifically react with NFK. To limit the potential problem of autofluorescence of library compounds and also to limit the problem of compound interference, a probe was selected that gave a fluorescent product with relatively long absorption and emission wavelengths, to reduce interference with autofluorescent compounds in HTS libraries. The NFK-specific chemical probe, NFK Green, reacts specifically with NFK and not with L-tryptophan or kynurenine ( Fig. 1 ). The reaction product has an emission optimum wavelength at 510 nm and an excitation optimal wavelength of 400 nm, which can be detected on most multimode readers. The NFK Green signal is linear proportional to the NFK concentration up to at least 200 µM NFK, and the minimal detectable NFK concentration is 1 µM ( Fig. 1 ). To investigate the potential use of the probe in cellular assays, calibration curves were prepared in a variety of cell culture media. In all cases, fluorescence signal was linear to NFK concentration and was not affected by the presence of bovine calf serum (examined up to a concentration of 10 % (v/v)) in the medium.
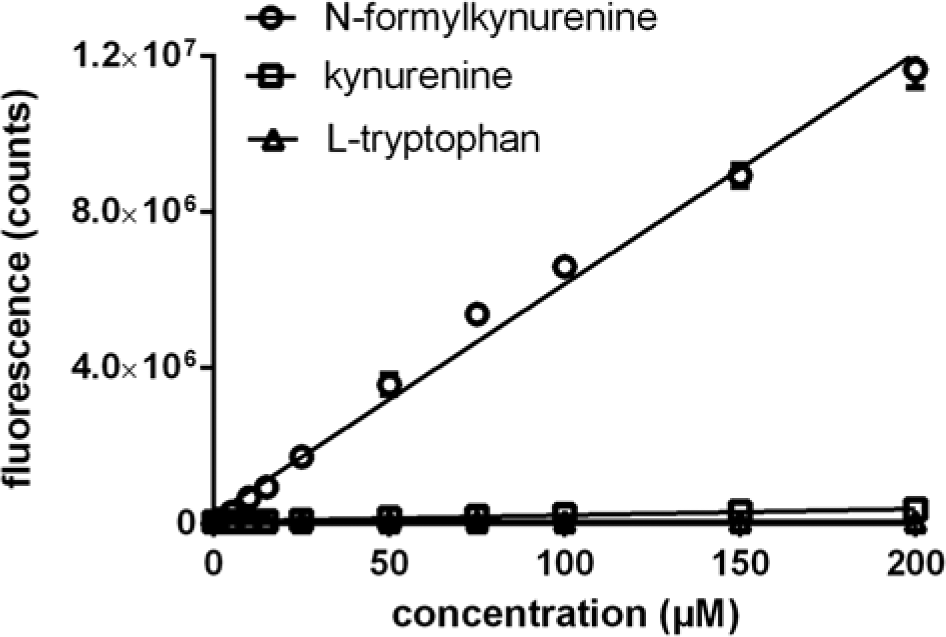
Selectivity of N-formylkynurenine (NFK) Green for NFK and dynamic range. Reaction of NFK Green with NFK, by measurement of fluorescence at 510 nm, and lack of reaction with L-tryptophan and kynurenine.
Biochemical Assay Development
Human IDO1 and TDO were expressed in E. coli as hexahistidine-tagged proteins and purified by affinity chromatography to >95% purity. Enzymatic activity was characterized in spectrophotometric assays measuring the conversion of L-tryptophan to NFK by detecting the UV absorption of NFK. To facilitate high-throughput and automated testing, biochemical assays for IDO1 and TDO were developed to be “addition only.” First, compound and enzyme were combined. The enzyme concentrations in both assays were minimized to enable detection of potent IC50s without ligand depletion effects. A preincubation step was introduced to enable equilibrium binding of slow-binding compounds. Next, the reaction was started by addition of L-tryptophan. For the high-throughput assay, L-tryptophan concentrations were chosen at the KM,Trp for TDO to ensure a sufficient assay window but also sufficient sensitivity to detect L-tryptophan competitive inhibitors. For IDO1, the L-tryptophan concentrations were chosen at 16 times the K to ensure a sufficient assay window. After 15 to 60 minutes, when the enzymatic conversion was still in the linear range, the reaction was stopped by the addition of NFK Green and developed by incubation at 37 °C for 4 h. Typically, at the end of the reaction, 11.7% of the substrate was converted to NFK for IDO1 and 3.1% for TDO. Average Z′ factor was 0.7 ± 0.1 for IDO and 0.8 ± 0.05 for TDO. Extending the NFK Green reaction overnight increased the assay window but had no effect on Z′ factor. Incubation times shorter than 3 h resulted in a lower S/B but did not affect the Z′ factor.
Since NFK Green is sensitive to pH, it is important that the reaction is developed at atmospheric CO2 (0.03%) rather than at the 5% CO2 usually maintained in incubators for cell culture. However, cell culture incubators can be used if plates are sealed with plastic seal after the addition of NFK Green. Because both IDO1 and TDO use oxygen as a substrate, 28 we took care to never cover plates during the enzyme reaction or to place them in an anoxic environment. Flushing the assay reagents in a cuvette with nitrogen was sufficient to stop NFK formation completely in the case of TDO and, counterintuitively, not for IDO. Furthermore, the addition of strong reducing agents, such as dithiothreitol (DTT), decreased enzyme activity, probably because of a reaction with oxygen. Finally, because both enzymes contain heme, which is active in the ferrous (Fe2+) state and inactive in the ferric (Fe3+) state, it is important to maintain a reductive environment that does not scavenge oxygen and keeps the heme group in the reduced state. Ascorbate, which is present in the reaction buffer of both enzymes, serves this role. 29 The reaction buffer for the IDO1 assay also contains methylene blue and catalase. Catalase acts as an H2O2 scavenger in case of auto-oxidation of ascorbic acid. 20 Methylene blue is considered an electron carrier from ascorbic acid to the ferric enzyme, 30 which could explain why the IDO1 assay is not completely inhibited in the absence of oxygen.
The pharmacology of the assay was verified with reference inhibitors ( Fig. 2 ), including, for IDO1, the preclinical compound Compound 5l9; S7111, an inhibitor published by NewLink Genetics, Inc. (Ames, IA) 27 ; and two historically frequently used inhibitors, 1-MT 5 and norharmane 26 ( Fig. 2 , Table 1 ). For TDO, the assay was validated using a compound described by Wellcome Research Laboratories in 1996, 18 which was synthesized in house and is referred to as NTRC 0672-0 ( Fig. 2 , Table 1 ). An analogue, described by Dolušić et al., 25 was also synthesized and is referred to as NTRC 0989-0 ( Fig. 2 , Table 1 ). Compound 5l and S7111 potently inhibited IDO1, with IC50 values around 15 nM and 100 nM, respectively ( Fig. 3A ), which are close to literature values—that is, 67 nM for Compound 5l9 and 75 nM for S7111 (Selleck/Absource GmbH). Notably, both compounds also potently inhibited TDO ( Fig. 3B ). Compound 5l was one-half log unit more active on IDO1, whereas S7111 was one-half log unit more active on TDO ( Table 1 ). NTRC 0820-0, a compound described by Röhrig et al., 19 was more than 20 times selective for IDO1 (i.e., 1½ log unit difference in IC50) ( Table 1 ). NTRC 0672-0 and NTRC 0989-0 are low micromolar potent inhibitors of TDO and do not inhibit IDO1 ( Fig. 3B , Table 1 ).
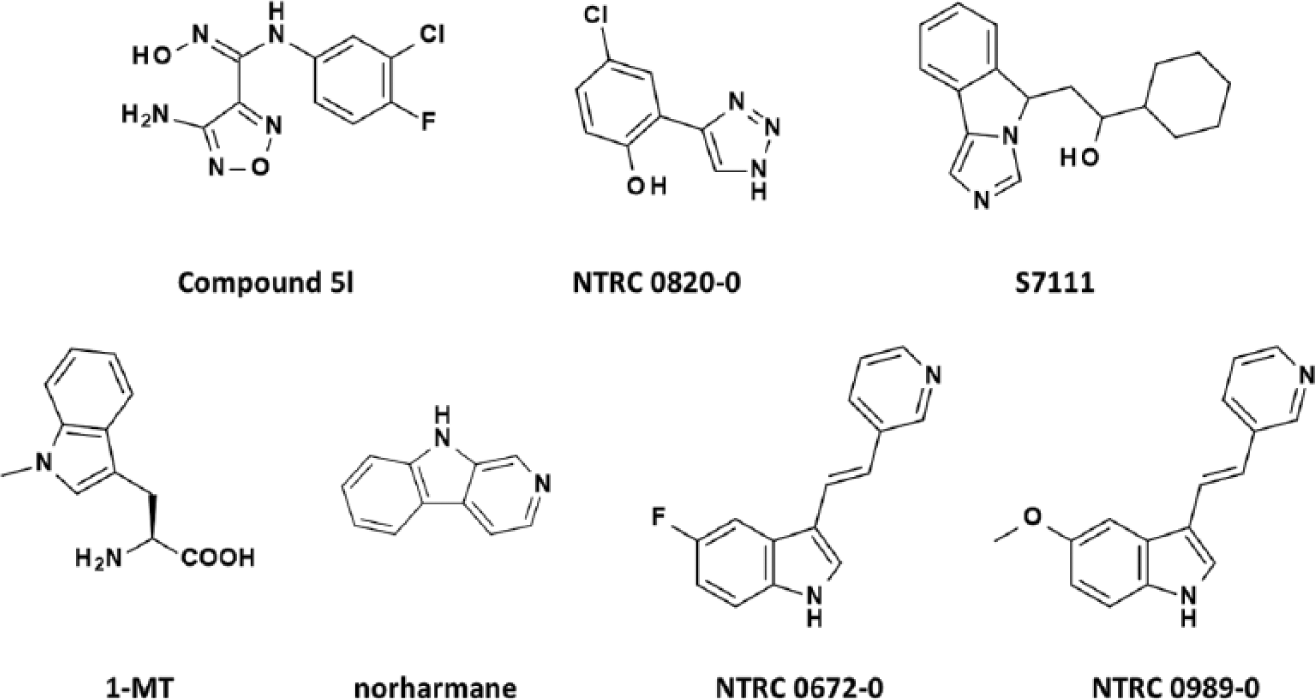
Chemical structures of indoleamine 2,3-dioxygenase (IDO1) and tryptophan 2,3-dioxygenase (TDO) inhibitors used in this study. S7111 is an inhibitor from NewLink Genetics, Inc. (Ames, IA) but referred to here under a Selleck/Absource GmbH (Munich, Germany) catalog code to ensure a unique identifier.
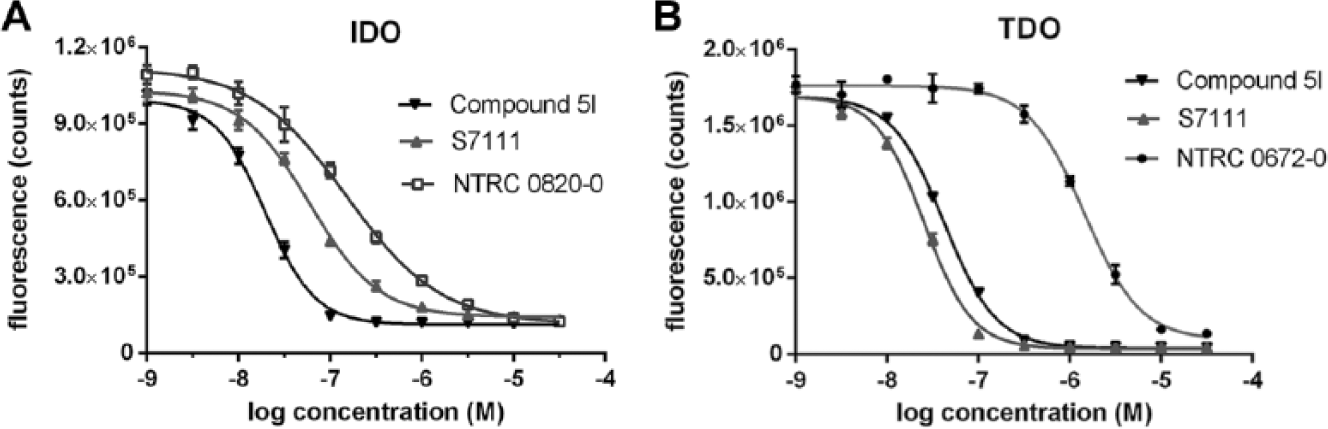
Inhibitor sensitivity of indoleamine 2,3-dioxygenase (IDO1) and tryptophan 2,3-dioxygenase (TDO) in N-formylkynurenine (NFK) Green assay. (
Cell-Based Assays
Several tumor cell lines have been reported to express IDO1, TDO, or both. 11 By measuring NFK production with NFK Green in the supernatant of cell lines cultured overnight in the presence or absence of L-tryptophan, we confirmed the relatively high tryptophan-catabolizing activity of the A-172 glioblastoma cell line. 11 Fluorescence could be inhibited in a dose-dependent manner by addition of a TDO inhibitor ( Fig. 4A ) and not by an IDO1 inhibitor (not shown), indicating that the production of NFK was due to overexpression of TDO. In total, all 44 cancer cell lines from NTRC’s Oncolines cancer cell line panel, 31 which have been derived from diverse tumor tissues, were screened for tryptophan-catabolizing enzyme activity. Other than A-172, TDO activity was detected only in colorectal carcinoma SW48 cells and only after a longer incubation time (i.e., after 72 h) (data not shown).
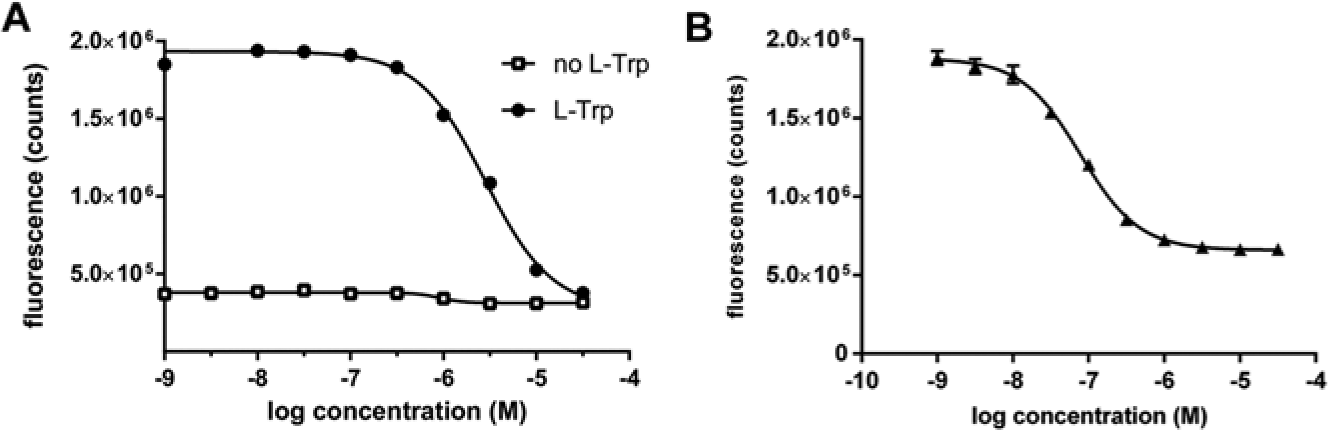
Cell-based assays for tryptophan metabolizing enzymes. (
The breast cancer cell line MDA-MB-231 has been shown to express IDO1 upon stimulation with IFN-γ 32 and was used to set up a cellular assay for the evaluation of IDO1 inhibitors. Three cell lines were identified in the Oncolines panel that showed increased tryptophan conversion after stimulation with IFN-γ. All three were colon cancer cell lines (i.e., LS 174T, SW480, and HCT-116). IFN-γ–stimulated tryptophan-catabolizing activity could be inhibited with the IDO1 inhibitor NTRC-0820 ( Fig. 4B ) or with Compound 5l, whereas TDO-specific inhibitors were not active (not shown). The data show that NFK Green is an efficient chemosensor to detect NFK production in cellular assays for IDO1 and TDO.
HTS
To further validate NFK Green as an assay readout for IDO1 and TDO and to demonstrate its application in HTS, a library of 87,000 compounds was screened for inhibitors of IDO1 or TDO. For TDO, the assay was further miniaturized to 1536-well format and a final assay volume of 5 µL. All compounds were screened at a concentration of 10 µM. Average Z′ factor of the IDO1 screen was 0.62 with an S/B of 5.6 ( Fig. 5A ). Average Z′ factor of the 1536-well TDO screen was 0.76 with an S/B of 24.5 ( Fig. 5B ). With a threshold set at 50% inhibition, the hit rate was 0.19% for IDO1 and 1.3% for TDO. Compounds with greater than 50% inhibition were retested at a concentration of 10 µM. Of the hits, 53% were confirmed for IDO1 and 70% for TDO. All compounds were subsequently examined for possible interference with complex formation of pure NFK and NFK Green, in an assay where compounds dissolved in reaction buffer were mixed with purified NFK in the absence of enzyme. With this NFK deselection assay, none of the hits on IDO1 and 10% of the hits on TDO were deselected, demonstrating the use of this assay to improve hit quality. Dose-response testing of the hits was performed, revealing 36 hits with IC50 <1 µM for IDO1 and 331 hits with IC50 <1 µM for TDO.
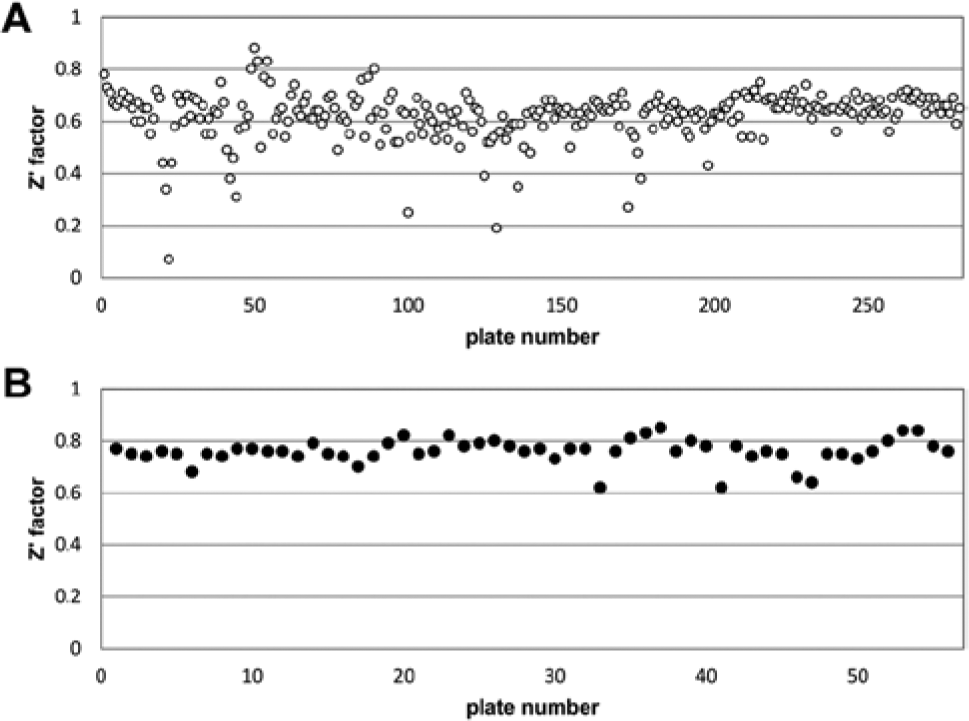
Performance of indoleamine 2,3-dioxygenase (IDO1) and tryptophan 2,3-dioxygenase (TDO) N-formylkynurenine (NFK) Green assays in high-throughput screening. (
Discussion
Whereas IDO1 and TDO are increasingly recognized as important drug targets, there are few tools available to screen compound collections for small-molecule inhibitors using enzymatic or cellular assays. Efforts in medicinal chemistry have been primarily focused to mimic the indole moiety of tryptophan, resulting in a limited chemical diversity of synthetic IDO1 and TDO inhibitors.10,13,25,26 However, when random screening has been attempted, nonindole scaffolds amenable for medicinal chemistry have been identified, as illustrated by INCB024360, the most advanced IDO1 inhibitor, which originates from a screening hit. 9
The critical step in any assay for IDO1 or TDO is the detection of NFK, the product of the enzyme reaction. Here, we present a new fluorescent assay for tryptophan metabolizing enzymes, based on a specific chemical probe that reacts with NFK to form a green fluorescent product. The NFK Green assay is a homogeneous mix-and-measure assay that avoids heating and addition of very acidic or toxic reagents, common for other IDO/TDO assays.19–23 We have demonstrated that the reagent can be used to detect NFK in biochemical assays of IDO1 and TDO, as well as in cellular assays of tryptophan consumption. Potentially, it can also be used in other assays, such as detection of arylformidase activity, which depletes NFK, and IDO2, a recently identified homologue of IDO1 with low tryptophan metabolizing activity. 33
The activity of a number of published IDO1 and TDO inhibitors was compared in the assays. The preclinical compound Compound 5l was by far the most potent IDO1 inhibitor, with an IC50 of 15 nM. Two other rationally designed compounds, NTRC 0820-0 and S7111, were also more potent than the natural products 1-MT and norharmane. Actually, we found both natural products to be very weak inhibitors of IDO1, with an IC50 >33 µM (pIC50 >4.5), which is in agreement with literature, reporting Ki values of 10 µM for 1-MT 6 and 120 µM for norharmane. 26 The most selective IDO inhibitor tested is NTRC 0820-0, which is more than 20 times selective for IDO1 over TDO.
As for the inhibition of TDO, NTRC 0672-0 and NTRC 0989-0 are the most selective inhibitors, with an IC50 in the low micromolar range in biochemical assays and no activity on IDO1. NTRC 0672-0 is somewhat more potent compared with NTRC 0989-0 and is therefore the best TDO tool inhibitor. Our side-by-side comparison of reference inhibitors reveals that Compound 5l and S7111 cross-react potently with TDO. Interestingly, other laboratories found that Compound 5l is a selective IDO1 inhibitor without any activity on TDO.9,34 The difference might come from these groups using transfected cell lines rather than biochemical assays with purified enzymes. Based on our result, the tumor-suppressive effects of Compound 5l in vivo could be related to IDO1 inhibition but also to TDO or dual TDO/IDO1 inhibition. More selective inhibitors are required to make a pharmacologically more rigid distinction between both targets. On the basis of our assay results, we propose applying the tool compounds NTRC 0672-0 for TDO and NTRC 0820-0 for IDO1.
The IDO1 and TDO biochemical assays were applied to screen a library of 87,000 compounds in 384-well or 1536-well format, respectively. The assays proved very robust with Z′ factors of 0.6 and 0.8 and a hit rate of 0.2% and 1.3%, respectively. This shows that the screening collection did not harbor many IDO or TDO inhibitors, but they could be efficiently identified with the NFK Green assay. A significant number of confirmed positive hits contained nonindole scaffolds, with several considered by a medicinal chemistry program. As such, the NFK Green assay has opened up new areas of chemical space for discovery of inhibitors of two important drug targets.
Footnotes
Acknowledgements
We thank Linda Brouwers for protein expression and enzyme assays and Hans Jansen, Gerben Rovers, Helma Rutjes, and Jan van Groningen from the Pivot Park Screening Centre for automation and screening support.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.