
Abstract
Vascular endothelial growth factor (VEGF) is a potential target for cancer treatment because of its role in angiogenesis and its overexpression in most human cancers. Currently, anti-VEGF antibodies have been shown to be promising tools for therapeutic applications. However, large size, poor tumor penetration, immunogenicity, and production in cost- and labor-intensive conditions are major drawbacks of such agents. The antigen-binding regions of camelid single-chain antibodies (VHHs), due to their unique biophysical characteristics, offer an alternative to conventional antibodies for tumor-targeting purposes. The present study was undertaken to generate and characterize anti-VEGF VHHs from an immune VHH library using phage display. Four rounds of panning were performed, and selected VHHs were characterized using various immunological techniques. Assessment of the antigenic profile of VHHs was done using competition enzyme-linked immunosorbent assay (ELISA). Selected VHHs reacted strongly to VEGF in indirect ELISA and cross-reactivity ELISA tests. The binding affinity of three VHHs, ZFR-1, ZFR-2, and ZFR-5, ranged from 2.5 to 80 nM, and among them, ZFR-5, which was selected for proliferation assay, significantly inhibited the endothelial cell growth in a dose-dependent manner. Taken together, our results indicate that ZFR-5 and other VHHs may be promising tools in cancer research and treatment.
Keywords
Introduction
Angiogenesis is a highly regulated and complex event required for a variety of normal proliferative processes. 1 Angiogenesis has also been shown to be essential for tumor growth and metastasis. 2 This process is the result of a dynamic balance between proangiogenic and the antiangiogenic factors. 3 So far, several angiogenic factors have been identified, including vascular endothelial growth factor-A (VEGF-A; generally referred to as VEGF), basic fibroblast growth factor, transforming growth factor–α and –β, tumor necrosis factor, platelet-derived endothelial growth factor, hepatocyte growth factor, angiogenin, interleukin-8, and placenta growth factor. VEGF is distinct among these factors in that it acts exclusively on endothelial cells during angiogenesis. 4 The angiogenesis effects of this molecule are mediated by its high-affinity receptors, human kinase domain receptor (KDR), and Fms-like tyrosine kinase receptor (Flt-1), located on vascular endothelial cells. 1 VEGF165, the mature and active form of VEGF, is a strong inducer of vascular permeability that promotes the migration and differentiation of endothelial cells, remodeling of the extracellular matrix, formation of capillary tubules, and vascular leakage. 1 Increased expression of VEGF has been reported in several types of human tumors and has been shown to correlate with tumor-associated angiogenesis, tumor growth, and a poor prognosis. 5 Experiments with neutralizing antibodies and other inhibitors demonstrated that blocking the VEGF pathway is sufficient to significantly suppress angiogenesis associated with solid tumors. 1 Thus, VEGF-targeted therapy may provide a promising approach for cancer treatment. Until now, several strategies targeting the VEGF axis have been developed, including anti-VEGF antibodies, soluble receptors binding directly to the VEGF ligand, anti-VEGF receptor (VEGFR) antibodies, and VEGFR tyrosine kinase inhibitors.6,7 Anti-VEGF antibodies have been shown to be effective in reducing tumor size, metastasis, and blood vessel formation in vivo. 8
Critical factors in the development of effective monoclonal antibodies for cancer therapy are specificity, high solubility, stability, and a small size. 9 Thus, many attempts to reduce the size of the conventional monoclonal antibodies (~160 KD), while retaining their antigen-binding properties, have been reported. 10 Various formats of antigen-binding fragments, including Fab and scFv, have been generated by phage display technology. In this technique, variable domains of the heavy and light chains are expressed on the surface of filamentous phages by fusion to an endogenous phage coat protein, and antigen-binding fragments are isolated by affinity selection. 11 These fragments have improved tumor penetration and reduced immunogenicity compared with the whole antibody. 9 They are also easier and faster to produce in recombinant form. Despite their beneficial properties, these fragments are still needed to be improved in terms of stability, expression yield, protease resistance, and aggregation caused by synthetic linkers. 9
Members of the Camelidae family generate a unique class of immunoglobulins that consist of two identical heavy chain molecules, devoid of light chains. 12 The antigen-binding region of these antibodies, known as VHH or nanobody, is the smallest functional antigen-binding fragment (~15 KDa) with a diameter of 2.8 nm and height of 4.4 nm. 9 VHHs possess important technological and biophysical advantages. Their small size and effective tissue penetration make them ideal molecules for targeting antigens in hindered locations and tumors. 9
Four hydrophobic amino acids in the framework 2 region (positions 37, 44, 45, and 47; Kabat numbering) that are highly conserved in conventional VHs are substituted in VHHs, with more hydrophilic residues. 13 These substitutions along with the extended length of the CDRH3 loop (17 residues in comparison with 9–12 residues in classical CDRH3 loops) and diversity enhance the solubility and stability at high concentration in aqueous solutions. 14 It has also been described that VHHs naturally show a high sequence homology with the human VH of family III. 14 Moreover, VHH coding DNAs can readily be cloned and expressed in bacteria and yeasts to yield high levels of soluble protein. 15 Contrary to conventional antibodies, VHHs have been shown to remain functional at 90 °C. 16
The main purpose of the present study was to generate and characterize VHHs against VEGF. We used phage display method to generate specific VHHs. Selected VHHs were characterized in terms of binding specificity, recognition of native VEGF, and affinity. Furthermore, we assessed an antigenic profile of selected VHHs and their binding relation to bevacizumab. Proliferation inhibition assay was also performed to test VHH potential in the neutralization of biological activity of VEGF.
Materials and Methods
Materials
Recombinant human VEGF165 (rhVEGF165) was obtained from BioVision (Milpitas, CA). Anti-M13 monoclonal antibody conjugated to horseradish peroxidase (anti-M13-HRP) and HRP conjugated mouse anti-hemagglutinin (anti-HA) were supplied by Roche (Mannheim, Germany). Isopropyl-β-D-thiogalactopyranoside (IPTG) was purchased from Fermentas (Germany). DAB (3, 3′-diaminobenzidine) and goat anti-human Fc-HRP antibody were obtained from Sigma (St. Louis, MO). The nickel nitrilotriacetic acid (Ni+-NTA) resin was supplied by Qiagen (Hilden, Germany). Bevacizumab was purchased by Roche (Basel, Switzerland). The 6-well plates and MaxiSorp 96-well plates were from Nunc (Roskilde, Denmark). The MTT Cell Viability Assay Kit was supplied by Biotium (Hayward, CA).
Bacterial Strains and Plasmids
Escherichia coli TG1 (Pharmacia, Uppsala, Sweden) and E. coli Rosetta-gami 2 (Novagen, Madison, WI) were used as hosts for phagemid manipulation and expression of soluble VHHs, respectively. The pComb3X and M13KO7 were used as a phagemid vector and a helper phage, respectively, for biopanning.
2XYT medium, supplemented with ampicillin, and Luria-Bertani medium, supplemented with MgCl2 and glucose, were used for the selection of clones and electroporation, respectively.
Cell Culture
Primary-cultured human umbilical vein endothelial cells (HUVECs) were obtained from Pasture cell bank (Iran Pasture Institute, Tehran, Iran) and cultured in DMEM (Gibco/Invitrogen, Carlsbad, CA). The medium was supplemented with 20% fetal bovine serum (FBS; Gibco/Invitrogen), 2 mM L-glutamine, penicillin (100 µg/mL), and streptomycin (100 µg/mL). Cells were grown at 37 °C in a 5% CO2 humidified incubator. HUVECs between passage 2 and 3 were used in this research.
VHH Library Biopanning
A previously prepared, an immune VHH library derived from Camelus dromedaries immunized with homogenized human cancerous tissues 17 was used for VHH selection. This VHH library harbored in phagemid vector pComb3X (complexity of ~109) was subjected to four rounds of panning in 6-well plates. In this study, a subtractive biopanning in which one well was coated with bovine serum albumin (BSA) as an irrelevant antigen and another with the rhVEGF165 (VEGF hereafter) as a target antigen was performed. One well of a 6-well plate was coated with 2 mL of 0.5 µg/mL VEGF in coating buffer (0.1 M NaHCO3, pH 8.6) and stored at 4 °C for 16 h. The subtraction well was coated with 2 mL of 0.5 µg/mL BSA. Target and subtraction wells for all panning rounds were coated with the same amount of solutions and incubated at 4 °C for 16 h. The wells were washed three times with phosphate-buffered saline (PBS) and blocked for 90 min at room temperature with 5% skimmed milk in PBS (5% MPBS). Phages from each round of the panning were also preincubated with the blocking solution for 1 h at 37 °C before phages were added to coated wells. Three milliliters of the phage library mix, consisting of approximately 1012 phages in 5% MPBS, was added to the subtraction well, incubated for 1 h, and the supernatant was transferred to the target well coated with VEGF and incubated for 1 h rocking at 37 °C. The supernatant was discarded, and nonspecific phages were removed by washing two times with PBS containing 0.5% (v/v) Tween 20 (0.5% PBST) followed by two times with PBS. Stringency of washing was increased using 0.5%, 1%, 1.5%, and 2% Tween 20 for rounds 1, 2, 3, and 4 of panning, respectively. Bound phages were eluted with 1 mL of 100 mM freshly prepared solution of triethylamine (pH 10). After incubation for 10 min, the eluted particles were neutralized with 1 mL of 1M Tris-HCl buffer (pH 7.4). After each round of selection, 100 µL of eluted phages were used to infect mid-log phase E. coli TG1 (OD600 = 0.6) grown at 37 °C for phage titration. Enrichment value of VEGF-specific VHHs was also assessed to monitor the progress of the selection process. Remaining eluted phages were used to infect E. coli TG1 (OD600 = 0.6) for subsequent amplification. The bacteria were subsequently superinfected with M13KO7 helper phage at a ratio of 20:1 (phage: bacteria) to rescue phage particles. A mixture of kanamycin (50 µg/mL) and ampicillin (100 µg/mL) was added to the culture, and bacteria were further grown overnight with shaking at 220 rpm at 37 °C. The cultures were centrifuged at 3700 × g for 10 min, and the supernatants were added to 20% (w/v) polyethylene glycol 6000/2.5M NaCl (PEG/NaCl) to precipitate the phages. The samples were incubated on ice for 1 h and then centrifuged for 20 min at 19,000 × g at 4 °C. The pellets were resuspended in 4% MPBS, and 1012 transducing units were used in subsequent rounds of panning. The panning procedure was repeated for another three rounds.
Polyclonal Phage Enzyme-Linked Immunosorbent Assay
Following four rounds of panning, output from each round was screened by polyclonal phage enzyme-linked immunosorbent assay (ELISA). Wells of MaxiSorp 96-well plates were coated with 0.5 µg/mL VEGF in coating buffer overnight at 4 °C. An equivalent concentration of BSA was used as a control for nonspecific binding. After washing with PBS, the remaining protein-binding sites in the wells were blocked for 90 min at 37 °C with 5% MPBS. The blocking reagent was discarded, and approximately 1 × 1012 plaque-forming units (pfu) of amplified outputed phages (5 × 109 phage particles per well) were added to appropriate wells and incubated while shaking for 1 h at 37 °C. The supernatant was discarded, and nonspecific phages were eliminated by washing four times with 0.05% PBST. Detection of the interaction between antigen and the phage-VHH was performed using a 2500-fold diluted solution of anti-M13-HRP. After incubation for 1 h at 37 °C, the wells were rinsed with PBST and PBS, respectively. The color was developed using 100 µL/well of tetramethylbenzidine (TMB) substrate, and the enzymatic reaction was stopped by addition of 2M HCl. The absorbance was read at 450 nm using an ELISA plate reader (STAT FAX 2100 Awareness Technology Inc, Ramsey, MN).
Screening of Monoclonal Phage-VHH
A marked increase in binding affinity occurred after the third round of selection. Therefore, phage outputs from the third round were used to infect exponentially growing E. coli TG1 cells by incubating for 1 h at 37 °C. Infected cells were spread on a 2XYT plate containing ampicillin (100 µg/mL), subsequently, the plate was incubated overnight at 32 °C. Sixty randomly selected clones were picked and tested for binding to VEGF by phage ELISA as described above. ELISA-positive clones were defined as those that exhibited at least two times stronger ELISA signals on antigen-coated plates in comparison to signals on BSA-coated plates. In parallel, the genetic diversity of the ELISA-positive clones was determined using DNA sequencing.
Expression of Soluble VHHs
To produce soluble antibodies, E. coli Rosetta-gami 2 was infected with selected phages. Bacteria were grown at 37 °C in 2XYT medium supplemented with ampicillin until an OD600 of 0.9 was achieved. VHH expression was induced with 1 mM IPTG for 16 h at 30 °C. The cells were subsequently harvested using centrifugation at 4000 × g for 15 min at 4 °C and dissolved in PBS buffer. The suspension was sonicated for 10 min at 50% pulse (Dr. Hielscher, GmbH). After centrifugation for 20 min at 16,000 × g, VHH proteins were purified by immobilized metal affinity chromatography with 5 mL Ni+-NTA resin. For selection of functional soluble VHHs, plates were coated and blocked as described previously. One hundred microliters of cytoplasmic extracts obtained from induction experiments was added to each well and incubated for 1 h. The wells were washed three times with 0.05% PBST and incubated with a 1:10,000 dilution of HRP conjugated anti-HA for 1 h. The ELISA was developed using TMB substrate, and the A450 nm was measured.
Western Blotting
Protein extracts were initially separated on a 12% discontinuous sodium dodecyl sulfate–polyacrylamide gel (SDS-PAGE). Following SDS-PAGE, the separated proteins were transferred onto a nitrocellulose membrane and blocked overnight in 5% MPBS at 4 °C. The membrane was then incubated in a 5000-fold dilution of an HRP conjugated anti-HA for 1 h at 37 °C. After washing with 0.05% PBST and PBS, the membrane was developed with DAB.
Specificity
Cross-reactivity of selected VHHs with various antigens was examined by ELISA. VEGF, human epidermal growth factor receptor 2 (HER2/neu), tumor-associated glycoprotein 72 (TAG-72), endoglin, cancer-associated mucin (MUC1), porcine submaxillary mucin (PSM), casein, skimmed milk, and BSA were coated onto 96-well ELISA plates at 0.5 µg/mL. Wells were blocked and then incubated with selected VHHs. VHHs bound to the wells were detected by incubation with HRP conjugated anti-HA followed by development with HRP substrate. All experiments were carried out in triplicate.
ELISA Inhibition Assay
VEGF-specific VHHs were serially diluted in PBS buffer, and titration ELISA was performed. VHH concentrations gave 50% to 70% saturating signal in titration experiments, were incubated with increasing concentration of VEGF (0–0.5 µg/mL) or two irrelevant antigens, BSA and skimmed milk (50 μg/ml), for 1 h at 37 °C and then transferred to plates coated with VEGF. After 1 h incubation, the plates were washed and VHH bound to immobilized VEGF was detected by HRP conjugated anti-HA followed by development with HRP substrate. The percentage of binding was determined as follows: (OD in the presence of competitor/OD in the absence of competitor) × 100. Half-maximal inhibition (IC50) values were then calculated as the concentration of soluble VEGF or irrelevant antigens that blocked 50% of the VHH binding to the immobilized antigens.
Affinity Measurement
Affinity of selected VHHs toward the antigen were determined as described by Beatty et al. 18 Wells of 96-well microtiter plates were coated with 100 µL of two different concentrations of VEGF (0.5 and 0.25 µg/mL). Each antigen was coated in triplicate wells with triplicate background wells per clone. After blocking, wells were incubated for 1 h with serial dilutions of VHHs. The plates were washed and developed with TMB substrate as described before. The affinity constant (Kaff) of each VHH was calculated using the equation 1.
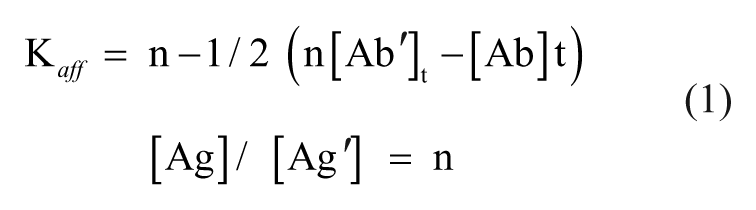
Competition ELISA
Wells of 96-well plate were coated with VEGF (0.5 µg/mL) overnight at 4 °C. Plates were washed with 0.05% PBST and blocked for 90 min at 37 °C with 5% MPBS. Bevacizumab, at a predetermined subsaturating dilution (0.5 µg/mL), was mixed with increasing concentrations of each of the selected VHHs (0-10 µg/mL) or anti-MUC1 VHH (negative control), and the mixtures were transferred in triplicate to VEGF-coated microtiter plates. After 1 h incubation, plates were washed and incubated with the goat anti-human Fc-HRP antibody. Plates were developed with TMB, and the absorbance was measured at 450 nm. Reductions in OD of less than 20%, 20% to 60%, and more than 60% were taken as an indication of noninterfering, moderately interfering, and strongly interfering binding, respectively, of the VHH to VEGF.
To further assess epitope binding of the VHH to VEGF, serial dilutions of bevacizumab (starting from 2 µg/mL) were examined in an ELISA with and without the constant concentration of ZFR-2 (5 µg/mL), and the mixtures were added to the VEGF-coated microtiter plate (0.5 µg/mL). After 1 h incubation and washing, wells were incubated with the goat anti-human Fc-HRP antibody, and microtiter plates were developed with TMB substrate.
Endothelial Cell Proliferation Inhibition Assay
HUVECs were plated in 96-well plates (5000 cells/well) in DMEM supplemented with 20% FBS and allowed to attach overnight. Medium was replaced with the medium containing 2% FBS. After 24 h, various amounts (0.5, 2.5, 5, and 10 µg/mL) of bevacizumab (positive control) or anti-MUC1VHH (negative control) or ZFR-5 VHH preincubated with a constant amount of VEGF (5 ng/mL final concentration) were added to the cells. Cells were then incubated for 72 h. During the incubation time, the medium was replaced with the fresh medium containing VEGF supplemented with bevacizumab or anti-MUC1VHH or ZFR-5, every 24 h. At the end of the incubation, MTT reagent (20 µL) was added to the wells and incubated for an additional 4 h at 37 °C. After incubation, the medium was removed, and the dye was solubilized by adding 100 µL dimethyl sulfoxide into each well. Absorbance was measured at a wavelength of 540 nm.
Statistical Analysis
Results are expressed as mean ± standard deviation (SD). Statistical analyses of the data were performed using one-way analysis of variance and t tests by SPSS. A p value of <0.05 was considered statistically significant.
Results
Monitoring of Panning and Enrichment of VEGF-Specific Binders
To find specific VHHs against VEGF, a VHH library containing ~109 independent clones was screened by panning. Four rounds of panning were carried out on VEGF coated on the microtiter plates. To evaluate the enrichment of VEGF-specific phages, total phage particles obtained by amplification of eluted phages after each round of panning were subjected to polyclonal phage ELISA. A sharp increase of signal intensity was observed after the third round of panning (data not shown).
Screening of VEGF-Specific Monoclonal Phages
The ELISA signal intensity and enrichment factor were markedly increased after the third round of selection. Therefore, outputed phages from the third round were used to assess VEGF binding of individual clones. Sixty individual clones obtained by infection of E. coli TG1 with the eluted phages were picked and tested for binding to VEGF using monoclonal phage ELISA. Out of 60 clones, 15 clones produced phage particles that bound to VEGF (data not shown). PCR analyses of these 15 clones showed that 6 clones, ZFR-1, ZFR-2, ZFR-3, ZFR-4, ZFR-5, and ZFR-6, contained an insert corresponding to the size of a VHH. Selected clones were chosen for additional characterization following their soluble expression.
Production of Soluble VHHs
Unlike TG1, Rosetta-gami 2 is not able to suppress the amber stop codon upstream of the gene III sequence in pComb3X vector, thus resulting in expression of soluble VHH. Therefore, we used E. coli Rosetta-gami 2 to produce soluble antibodies. After IPTG induction and purification, each VHH clone was analyzed by ELISA ( Fig. 1A ) and immunoblotting ( Fig. 1B ). Protein concentrations were determined using Bradford assay.
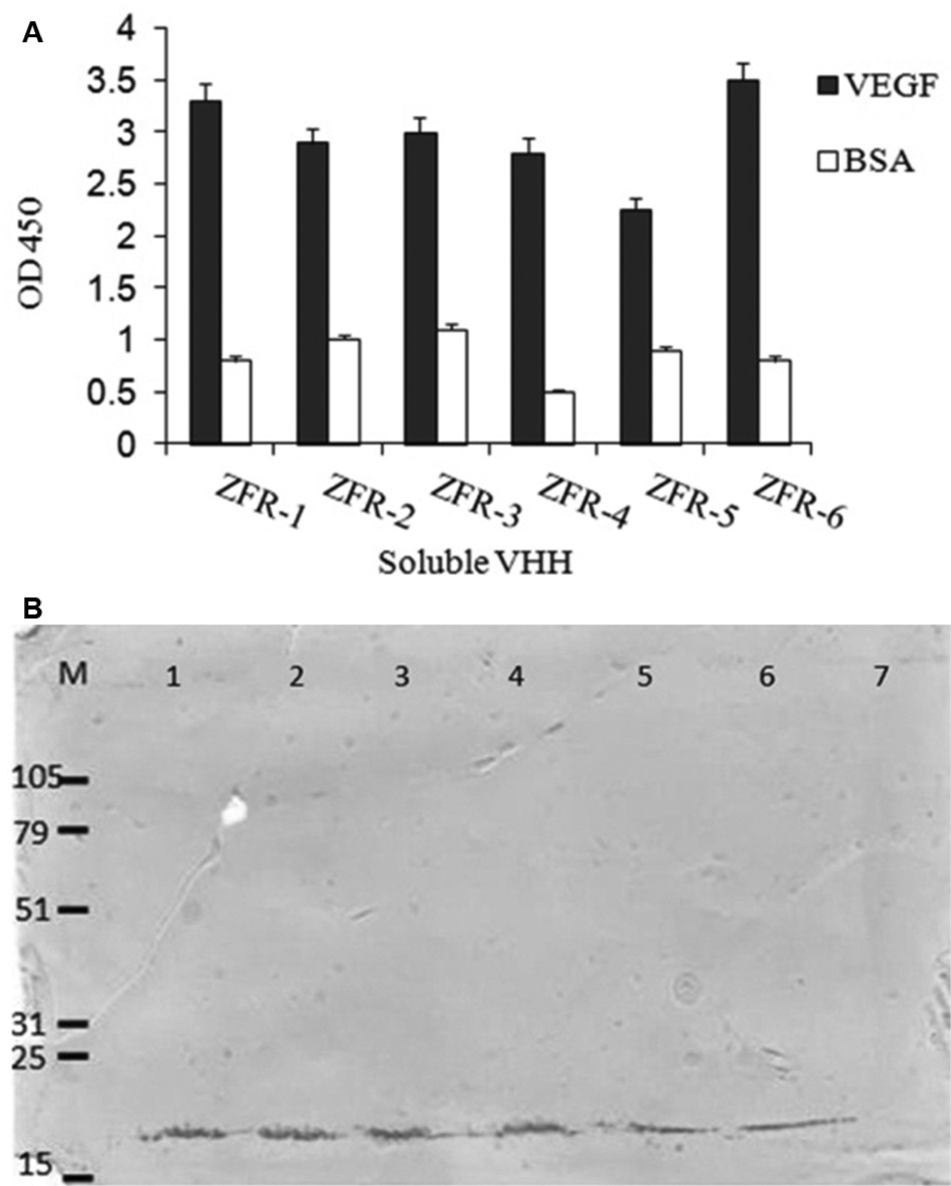
Results of production of soluble VHHs. (
Specificity
To assess the binding specificity of soluble VHHs, they were examined in a cross-reactivity ELISA using VEGF and different proteins including a growth factor (HER2/neu), glycoproteins (TAG-72, endoglin, MUC1, PSM
Specificity of soluble VHHs. a
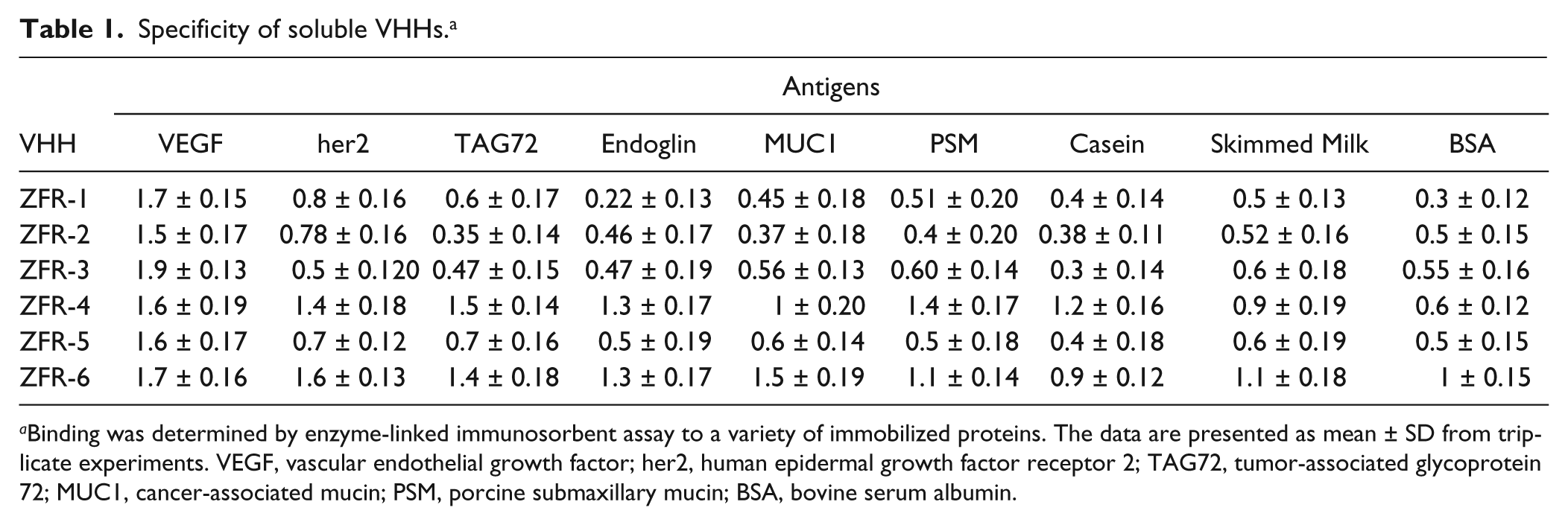
Binding was determined by enzyme-linked immunosorbent assay to a variety of immobilized proteins. The data are presented as mean ± SD from triplicate experiments. VEGF, vascular endothelial growth factor; her2, human epidermal growth factor receptor 2; TAG72, tumor-associated glycoprotein 72; MUC1, cancer-associated mucin; PSM, porcine submaxillary mucin; BSA, bovine serum albumin.
ELISA Inhibition Assay
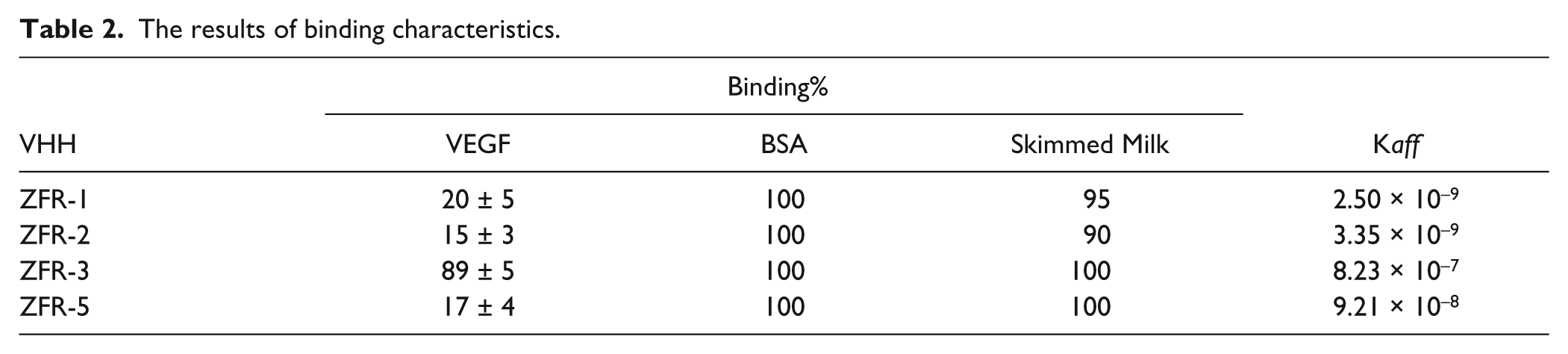
To investigate whether identified VHHs can recognize native VEGF, inhibition ELISA was performed. Table 2 shows the results of ELISA inhibition experiments. Except for ZFR-3, binding of selected anti-VEGF VHHs to immobilized VEGF was inhibited by preincubating the VHHs with 0.5 µg/mL solution of native VEGF. ELISA inhibition results indicate that three VHHs can recognize native VEGF, suggesting that these VHHs recognize conformational epitopes. As expected, irrelevant antigens such as BSA and skimmed milk, which were used at much higher concentrations than VEGF, could not compete with VEGF for binding to the VHHs, thus confirming that these VHHs are highly specific for VEGF ( Table 2 ).
The results of binding characteristics.
The binding curves of VHHs to immobilized VEGF are shown in Figure 2 . Based on the analysis of binding curves of the VHHs, the IC50 value (the concentration of soluble VEGF that blocked 50% of the VHH binding to the immobilized VEGF) for individual VHH was calculated. IC50 results of four anti-VEGF VHHs are shown in Figure 2 . These results suggest that IC50 for ZFR-3 is higher than 0.5 µg/mL.
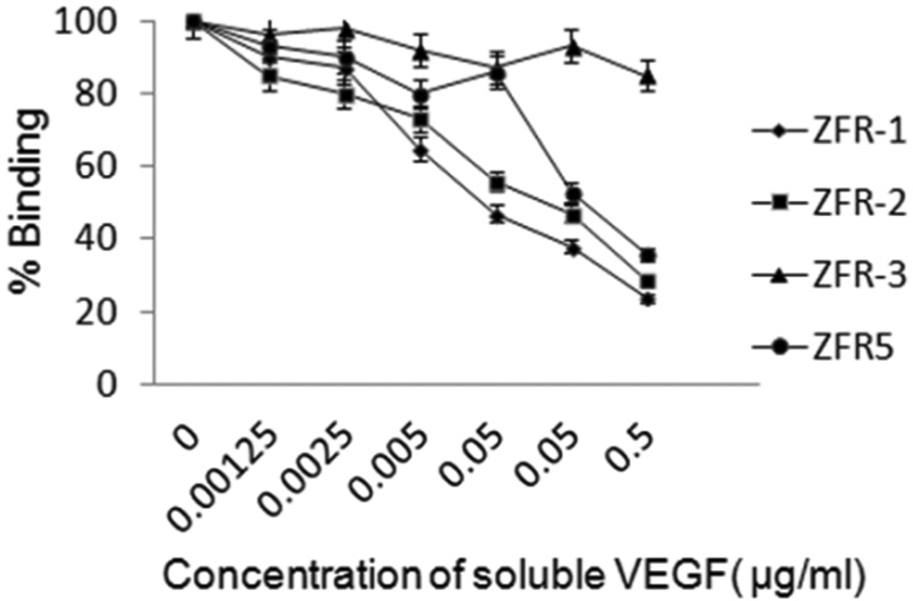
Enzyme-linked immunosorbent assay (ELISA) inhibition test. Wells were coated with 0.5 µg/mL vascular endothelial growth factor (VEGF), and the binding of soluble VHHs to immobilized VEGF was competed with different amounts of soluble VEGF. The percentage of binding is expressed as follows: [absorbance (A450) in the presence of soluble VEGF divided by absorbance (A450) in the absence of soluble VEGF] × 100. The data are presented as mean ± SD from triplicate experiments.
Affinity
Table 2 shows the results of affinity determination, according to the method of Beatty et al. 18
Epitope Mapping
Assessment of the antigenic profile of anti-VEGF VHHs was performed in two steps. First, competition ELISA was done to examine whether selected VHHs share any epitope with bevacizumab. 19 To this end, plates were coated with VEGF and incubated with bevacizumab (as a reference antibody) in the presence of increasing amounts of VHHs or anti-MUC1 VHH (as a negative control). Whenever the VHH recognized the same epitope of bevacizumab, binding to VEGF was reduced or abolished. Furthermore, when the VHH recognized the distinct epitope on VEGF, binding of bevacizumab to VEGF was not altered by VHH. As shown in Figure 3A , binding of bevacizumab to VEGF was not affected by anti-VEGF VHHs or by the negative control VHH at low concentrations. The VHH concentrations were further increased, and at intermediate concentration range, ZFR-1 showed very little competition. However, binding to VEGF was abolished at the maximum concentration of ZFR-1, indicating that its epitope positions near the binding site of bevacizumab. ZFR-5 showed no competition even at the highest concentration, suggesting that the epitope recognized by bevacizumab was separate enough to permit independent binding of ZFR-5 to VEGF. The negative control, anti-MUC1, also showed no competition even at high concentrations. The binding of bevacizumab to VEGF was increased when excess amounts of ZFR-2 were added. Subsequently, to confirm this experience, bevacizumab was tested in ELISA in serial dilution, in the presence or absence of a constant concentration of ZFR-2. In the presence of ZFR-2, the binding activity of bevacizumab was enhanced ( Fig. 3B ).
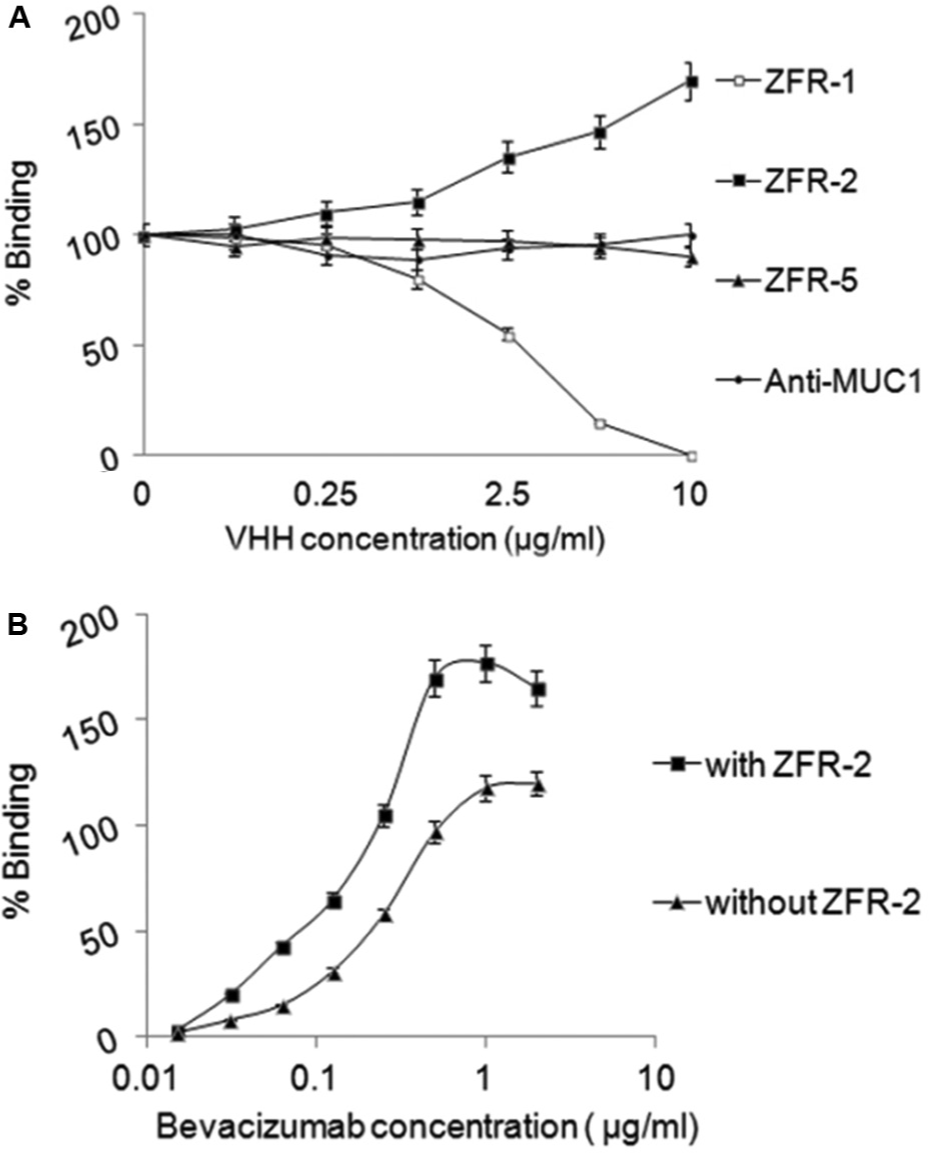
Assessment of antigenic profiles of selected VHHs. (
Inhibition of VEGF-Stimulated HUVEC Proliferation
Because the ZFR-5 VHH recognized a distinct epitope on the VEGF in comparison with bevacizumab, we decided to investigate whether this VHH might inhibit HUVEC proliferation in response to VEGF. To assess the effect of ZFR-5 on the proliferation of human endothelial cells, an MTT assay was performed. As shown in Figure 4 , ZFR-5 VHH significantly suppressed the proliferation response of HUVECs to VEGF in a dose-dependent manner. By contrast, equivalent concentrations of anti-MUC1 VHH (negative control) had no significant effect on the proliferation of endothelial cells.
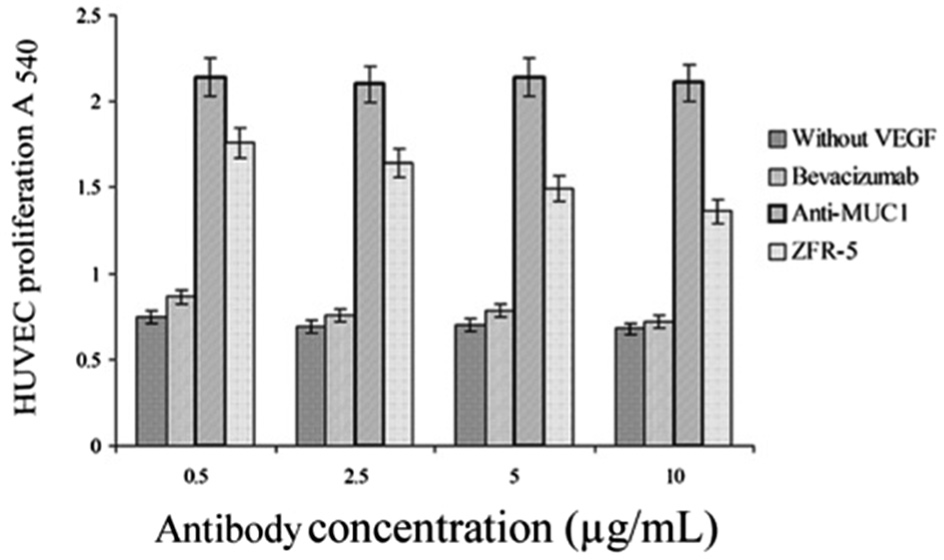
Inhibition of vascular endothelial growth factor (VEGF)–induced endothelial cell proliferation. Human umbilical vein endothelial cells (HUVECs) were cultured in the presence of VEGF preincubated with various amounts of bevacizumab (positive control) or anti-MUC1 VHH (negative control) or ZFR-5 at 37 °C. After 72 h, cell proliferation was assessed by MTT assay, and the absorbance was read at 540 nm. The data are presented as mean ± SD from triplicates experiments.
Discussion
The purpose of the present study was to generate and characterize specific VHHs against VEGF. Previous studies have demonstrated that VEGF plays an essential role in developmental angiogenesis and is important also in pathological angiogenesis. 1 VEGF-targeted therapies have been shown not only to be beneficial for tumor growth inhibition and metastasis prevention, but also might improve immunotherapy and the general condition of cancer patients. 20 Several VEGF pathway inhibitors have been approved for the treatment of various malignancies. 21 Bevacizumab, a humanized anti-VEGF monoclonal antibody, has been approved by the Food and Drug Administration (FDA) as a first-line treatment for metastatic colorectal cancer in combination with chemotherapy. 22 However, to date, bevacizumab therapy alone has demonstrated little antitumor activity or survival benefit in humans. 23 Moreover, cost intensiveness and labor intensiveness of the production of conventional monoclonal antibodies by the hybridoma technique and their purification, presence of the Fc that might impair a good tumor targeting, their large size, and immunogenicity are other hindrances to treatment of cancer by conventional monoclonal antibodies. 24
To overcome some of these shortcomings, varieties of smaller constructs, such as Fabs and scFvs, have been generated by phage display technology as alternatives to conventional antibodies for diagnostic and therapeutic applications. 25 Despite this, the use of antibody fragment libraries suffer from several limitations including the unsatisfactory yield of functional products in heterologous expression systems and aggregation caused by synthetic linkers. 26
Alternatively, VHHs provide desirable characteristics for tumor targeting including rapid tumor uptake and plasma clearance, reduced immunogenicity, resistance to serum proteases, as well as more homogeneous penetration of the tumor compared with larger antibody formats. 10 Other advantages of their small size are their intrinsic stability, easy cloning, solubility, binding to cavities and hidden antigens, and easy production. 16
In this study, we report the successful isolation and characterization of a panel of anti-VEGF VHHs. Several reports have already described the isolation of recombinant antibody and antibody fragments specific for the VEGF.27,28 However, to our knowledge, this is the first report describing the identification and in vitro characterization of specific VHHs for VEGF.
In this study, we used the phage display library derived from Camelus dromedaries 17 to isolate binders of VEGF. During the panning against VEGF, an increase in the number of binders was observed from round 2 to round 3 (data not shown). This observation indicates that clones from round 3 might have strong specificity and affinity for VEGF. To obtain monoclonal sources of antibody fragments, 60 randomly selected clones from round 3 were chosen for binding to VEGF. Out of these 60 clones, 15 clones giving strong ELISA signals on VEGF were isolated. PCR analyses of these 15 clones showed that 6 clones, clones ZFR-1, ZFR-2, ZFR-3, ZFR-4, ZFR-5, and ZFR-6, contained an insert corresponding to the size of a VHH. These clones were successfully produced in E. coli Rosetta-Gami 2. Then the soluble VHHs were analyzed by ELISA to investigate production of functional VHHs. Our results show that the binding intensity of soluble VHHs is significantly higher than that of the phage VHHs. This increase may be due to better interaction of soluble VHHs to coated VEGF in the absence of phage proteins contained in phage VHHs. To investigate binding specificity of selected VHHs, various proteins were examined in cross-reactivity assays. Two of these clones, clones ZFR-4 and ZFR-6, showed strong signals with irrelevant antigens. In addition, four VHHs showed a high specificity for binding to VEGF. The results of Western blotting against VEGF show that these VHHs recognized denatured VEGF. Moreover, three of these VHHs, ZFR-1, ZFR-2, and ZFR-5, react with native VEGF in ELISA inhibition. Therefore, ZFR-1, ZFR-2, and ZFR-5 VHHs most likely recognize linear epitopes that are also exposed in native VEGF. Interestingly, binding of ZFR-3 VHH to coated VEGF was not inhibited by incubation of VHH with 0.5 µg/mL of soluble VEGF. The most probable explanation for this phenomenon is that this VHH might recognize a buried epitope in native VEGF and becomes exposed following coating the VEGF on a solid phase.
To further characterize VHHs, binding affinity was determined using the Beatty method. 18 Binding affinity of these clones toward VEGF ranged from 2.5 to 80 nM, which is within the affinity range of other single-domain antibodies. 10 To examine whether the antigenic profile of generated VHHs is similar to that of bevacizumab, competition ELISA was performed. Our results showed that ZFR-1 showed partial competition with bevacizumab, indicating that its epitope positions near the binding site of bevacizumab. Moreover, according to the results of the competition experiment, ZFR-5 showed no competition even at the highest concentration, demonstrating that ZFR-5 recognizes a distinct epitope on the VEGF in comparison with bevacizumab. Interestingly, the binding of bevacizumab to VEGF was increased in the presence of excess amounts of ZFR-2. This increase may be due to the synergistic or additive effects of two antibodies. Based on the results of the specificity, ELISA competition test, and affinity measurement, three VHHs (ZFR-1, ZFR-2, and ZFR-5) were chosen as potential VEGF-targeted VHHs for further analyses. Because ZFR-5 recognized a different epitope on the VEGF other than bevacizumab, we became interested in investigating the ZFR-5 potential in neutralization of biological activity of VEGF. According to the results of the proliferation assay, ZFR-5 effectively inhibited HUVEC proliferation in a dose-dependent manner. However, equivalent concentrations of the control VHH had no significant effect on the cell proliferation. Although ZFR-5 was not as potent as bevacizumab in the HUVEC proliferation inhibition test, the lower potency of ZFR-5 is likely due in part to its monovalent format. Taken together, these results indicate that ZFR-5, due to the unique characteristics of a VHH, can lead to a novel antiangiogenic agent.
Currently, two VEGF pathway inhibitors, an anti-VEGF antibody (bevacizumab) 22 and two tyrosine kinase inhibitors (sorafenib and sunitinib), 29 have been approved by the FDA for cancer therapy. Although bevacizumab is indicated as a first- or second-line treatment in five different tumor types, several adverse effects are associated with bevacizumab therapy. 30 In addition, most small kinase inhibitors usually hit multiple targets. Moreover, for therapeutic use, small molecules are perfect candidates for targeting antigens in hindered locations.
This article has demonstrated the generation and characterization of high-affinity anti-VEGF VHHs. The main conclusion to be drawn from this study was that ZFR-5 or other VHHs have in vitro properties that are well suited for VEGF targeting. Further work needs to be done to establish whether selected VHHs have in vivo efficacy to act as antiangiogenic and antitumor agents. Finally, the favorable biophysical and pharmacological characteristics of VHHs and the ease with which they can be produced make them promising agents for diagnostic and therapeutic applications.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by Shahid Beheshti University of Medical Sciences, Tehran, Iran and Iran National Science Foundation (INSF) reference 90007324.