
Abstract
Cryptococcus neoformans is a pathogenic fungus that causes meningitis worldwide, particularly in human immunodeficiency virus (HIV)–infected individuals. Although amphotericin B is the “gold standard” treatment for cryptococcal meningitis, the toxicity and inconvenience of intravenous injection emphasize a need for development of new anticryptocccal drugs. Recent data from humans and animal studies suggested that a nutrient-deprived host environment may exist in cryptococcal meningitis. Thus, a screening assay for identifying fungicidal compounds under nutrient-deprived conditions may provide an alternative strategy to develop new anticryptococcal drugs for this disease. A high-throughput fungicidal assay was developed using a profluorescent dye, alamarBlue, to detect residual metabolic activity of C. neoformans under nutrient-limiting conditions. Screening the Library of Pharmacologically Active Compounds (LOPAC) with this assay identified a potential chemical scaffold, 10058-F4, that exhibited fungicidal activity in the low micromolar range. These results thus demonstrate the feasibility of this alamarBlue-based assay for high-throughput screening of fungicidal compounds under nutrient-limiting conditions for new anticryptococcal drug development.
Introduction
Cryptococcus is an opportunistic pathogen that mainly affects immunocompetent and immunocompromised individuals worldwide, killing approximately 600,000 people per year. 1 In the United States, cryptococcal infection continues to be a problem, with an estimated $54 million in direct hospitalization costs per year. 2 Standard treatment of cryptococcosis includes treatment with amphotericin B and fluconazole, but the disease continues to have up to a 60% mortality rate in developing countries where the disease is most common. 3 In addition, fluconazole-resistant strains have become widespread in these regions since the onset of the AIDS epidemic, 4 although this rate has decreased in developed regions where antiretroviral therapy is more widely available. 5 Such common resistance patterns and poor outcomes to fluconazole underscore the need to find novel and cost-effective drug candidates for individuals, especially in resource-poor settings.
A recent review highlighted the possible superiority of fungicidal versus fungistatic regimens in some infections. 6 For example, therapy of cryptococcal meningitis has traditionally been more effective in the early treatment period with amphotericin B–based fungicidal regimens, with improved initial outcomes and more rapid cerebrospinal fluid (CSF) sterilization. 7 More recent data by Bicanic et al. 8 have provided further microbiological support of this concept, showing that, while both regimens arrested fungal growth, measurable clearance from CSF was achieved only with fungicidal amphotericin-based regimens versus poor rates of clearance using standard doses of the fungistatic fluconazole. Further studies have shown that rates of clearance from CSF show a correlation with survival. 9 Such a requirement for fungicidal activity could be due to the unique nature of the fungal-host interaction in cryptococcal meningitis. CSF composition in fungal meningitis typically includes elevated protein and lactate levels 10 as well as low glucose11,12 that can result in considerable fungal stress. In addition, recent data implicating roles of fungal autophagy, 13 as well as the importance of high-affinity copper,14,15 glucose, 12 and nitrogen uptake, 16 suggest that effective survival within a nutrient-deficient environment may be key to successful virulence of the pathogen. In addition, virulence factors such as fungal laccase are most inducible in media that are nutrient deficient,17,18 and the presence of melanin has been found to increase resistance to antifungals, 19 leading to further challenges to treatment under nutrient-deprived conditions. Furthermore, poor clinical outcomes based on antifungal agents such as fluconazole that effectively inhibit growth under nutrient-rich conditions containing glucose, 5 but lose activity under nutrient-deprivation conditions, may suggest that effectiveness against a fungus in a reduced nutrient environment may better predict therapeutic success. In summary, these data suggest that screening of compounds for fungicidal activity under nutrient deprivation conditions may provide a new approach in drug discovery against this organism.
Thus, in the present study, asparagine minimal media, a nutrient-deficient media used previously to successfully classify high-virulence from low-virulence isolates from a cohort of organ transplant patients, 14 were used to induce nutrient stress response conditions for Cryptococcus neoformans. A fluorescent redox indicator, alamarBlue dye, was used to detect metabolic activity of C. neoformans as a cell viability assay for C. neoformans. The assay was found to successfully differentiate between the fungicidal response of amphotericin and the static response of fluconazole. A test screening of the Library of Pharmacologically Active Compounds (LOPAC) identified a fungicidal compound, 10058-F4, with potency in the low micromolar range. Therefore, this fungicidal assay under nutrient starvation conditions is useful for high-throughput screening (HTS) of large compound collections for the identification of effective new anticryptococcal lead compounds.
Materials and Methods
Optimization of Fungal Cell Concentration and Nutrients Using an alamarBlue-Based Fungicidal Assay in a 96-Well Format
Wild-type Cn strain H99 (ATCC 208821; American Type Culture Collection [ATCC], Manassas, VA) was grown in a 30 °C incubator overnight (24 h) to mid-log phase in YPD (2% glucose, 2% bactopeptone, 1% yeast extract), then washed twice in sterile ASN media (1 g/L asparagine, 10 mM sodium phosphate, pH 6.5). Then, 100 µL of 2.76 × 106 cells/mL of ASN was added to 96-well plates and serially diluted by one-half (total well volume 100 µL). Next, 0.5% glucose, 0.1% acetate, 0.1% lactate, or no substrate was added as indicated. Cells were then incubated at 37 °C overnight (24 h) with shaking. Then, 100 µL alamarBlue solution (Invitrogen, Carlsbad, CA) was added, and cells were again incubated overnight (24 h) at 37 °C with shaking. Fluorescence was read with λ of 530- to 560-nm excitation and 590-nm emission (TECAN Genios Fluorescence and Luminescence Plate Reader; TECAN, Männedorf, Switzerland).
alamarBlue Assay in a 1536-Well Format
The alamarBlue assay was miniaturized to a 1536-well plate format. Briefly, the C. neoformans suspension, prepared in ASN starvation medium, was plated at a seeding density of 2200, 4500, and 9100 cells/well with a final volume of 5 µL/well using the Multidrop-Combi dispenser (Thermo Scientific, Logan, CT) in black 1536-well plates. Cells were incubated for 24, 48, and 72 h at 37 °C supplied with 5% CO2. The alamarBlue dye (supplied by Invitrogen as 10× stock) was prepared as a 2× working concentration using ASN media and added at 5 µL/well. The assay plates were incubated at 37 °C supplied with 5% CO2 for 2 h. The assay plates were then read using fluorescence intensity mode (excitation = 525 nm, emission = 598 nm) on the ViewLux plate reader (PerkinElmer, Waltham, MA). Etest agar (bioMérieux Clinical Diagnostics, Marcy l’Etoile, France) minimum inhibitory concentration (MIC) was conducted on strain H99, reading inhibition of the growth at 24 h according to the manufacturer’s directions.
Validation and Screening of Pharmaceutically Active Compounds Using a Cryptococcal Fungicidal Assay
Wild-type Cn strain H99 (ATCC 208821; ATCC) was grown in a 30 °C incubator overnight to mid-log phase in YPD, then washed twice in sterile ASN media. Wells of a 96-well plate contained 1 × 106 cells/mL of mid-log or heat-killed (70 °C for 30 min) C. neoformans in ASN, as indicated, with no drug, 1% pure water, amphotericin B (0.5 µg/mL; Sigma Aldrich, St. Louis, MO) diluted in pure water, or fluconazole (8.0 µg/mL; Sigma Aldrich) diluted in DMSO, such that no more than 1% was DMSO solvent. Alternatively, 5 µM of compounds diluted in DMSO from a Library of Pharmaceutically Active Compounds (LOPAC; lot 031M4707/110M4713; Sigma) was added such that no more than 1% volume was DMSO. Cells were then incubated at 37 °C for indicated times (24, 48, or 72 h) with shaking. Then, 100 µL alamarBlue solution was added (Invitrogen), and cells were again incubated for 24 h at 37 °C with shaking, and fluorescence was measured (excitation = 530-560 nm, emission = 590 nm) using a TECAN Genios Fluorescence and Luminescence Plate Reader.
Effect of 10058-F4 on C. neoformans Survival by a Colony-Forming Unit Viability Assay
Wild-type Cn strain H99 (ATCC 208821; ATCC) was grown to mid-log phase at 30 °C in YPD, then washed twice in sterile ASN media. Then, 100 µL of a suspension of 7.87 × 103 cells/mL of ASN was added to two 96-well plates. No drug, 1% DMSO, indicated amounts of amphotericin B, or 10058-F4 diluted in DMSO was added such that no more than 1% volume was DMSO. Cells were then incubated at 37 °C with shaking for 24 h. Samples from each well were plated on YPD media, incubated in a 30 °C incubator for 48 h, and colonies counted.
Statistical Methods
Nonparametric statistical analyses (medians and interquartile ranges, IQRs) were performed using Minitab Student v.14 (Minitab, Inc., State College, PA). Excel (Microsoft, Redmond, WA) was used to graph medians and IQRs using bar graphs. Mann-Whitney was used for one-variable comparisons (effects of substrates on fluconazole activity, effect of amphotericin B and fluconazole, inherent fluorescence of amphotericin B and fluconazole). Medians were also calculated using the Friedman test to evaluate efficacy of fluconazole with increasing dosages and incubation times. The signal-to-basal (S/B) ratio was calculated by dividing the signal in C. neoformans–only wells by the basal signal from wells containing 50 µM amphotericin B that completely killed C. neoformans. All values were expressed as the mean ± SD (n = 3).
Results
Development of a Whole-Cell Fungicidal Assay for C. neoformans under Nutrient-Deprived Conditions
Cell density optimization
We used an alamarBlue dye that becomes fluorescent with metabolic activity of C. neoformans, conducted under nutrient-deplete conditions. In the presence of fungicidal compounds such as amphotericin B, the fluorescence signal yields resulting from an incubation of alamarBlue dye with C. neoformans were significantly reduced. Despite reduced metabolism due to the suboptimal nutrient conditions, the total fluorescent signal was correlated with cell number ( Fig. 1A ), demonstrating that this alamarBlue assay is a useful cell-viability assay for the fungus. The cell density of 1 × 106 cells/mL of the fungus yielded an optimal fluorescent signal and was used in subsequent 96-well plate experiments.
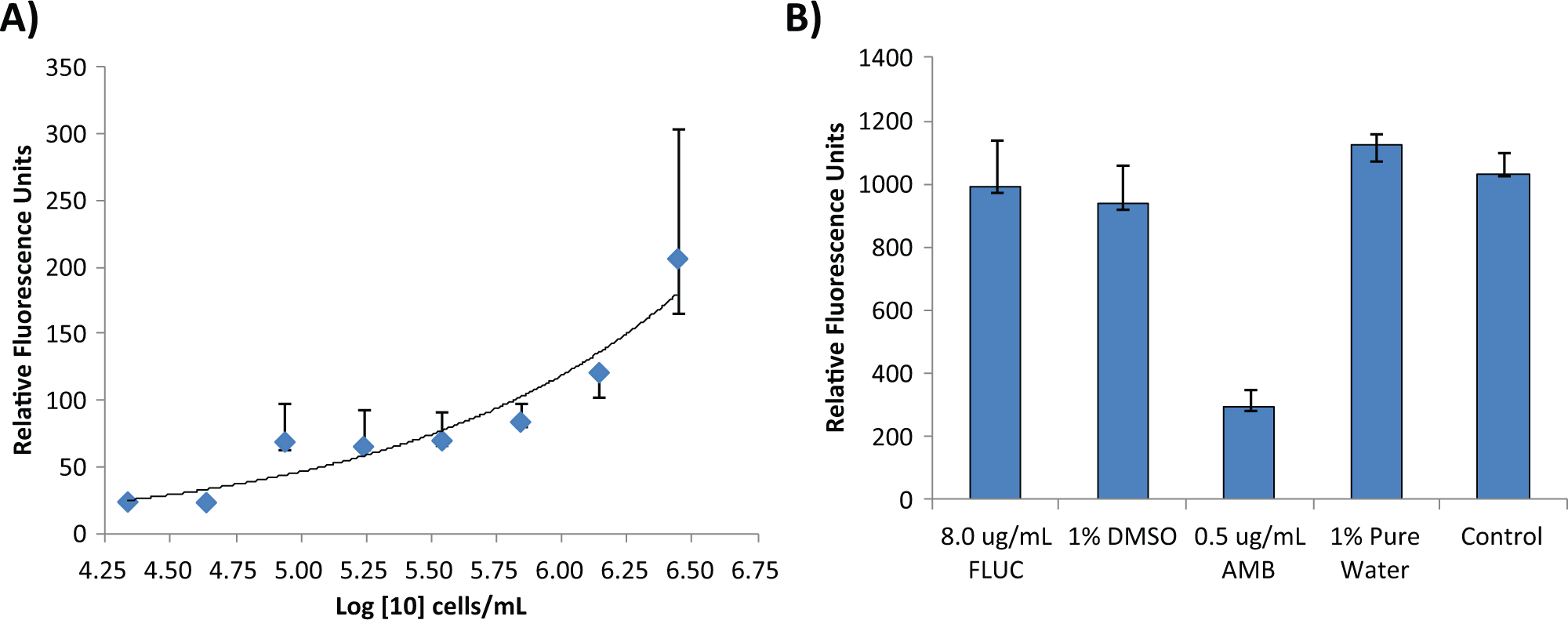
Cell density titrations and compound incubation times. Median relative fluorescence units for (
Fungicidal activity
We then examined the fungicidal and static effects of the antifungal drugs amphotericin B and fluconazole in this assay. As shown in Figure 1B , the fluorescent signal from C. neoformans treated with 0.54 µM (0.5 µg/mL) amphotericin B was significantly reduced, while the signal from 26.1 µM (9 µg/mL) fluconazole-treated cells showed no difference compared with the solvent (1% DMSO) control. To further confirm the fungicidal activity of amphotericin B, C. neoformans was killed by heating and then measured in the alamarBlue assay. We found that the fluorescent signal determined from heat-killed cells was similar to that after treatment with amphotericin B (data not shown). On the other hand, increasing fluconazole to over 20× the MIC (200 µg/mL) or addition of small amounts of nutrients such as 20 mg/dL of glucose, 50 mg/dL of acetate or lactate, or 0.5× yeast nitrogen base (data not shown) did not result in significant reductions in fluorescent signal, consistent with a lack of fungicidal activity under the conditions tested. In addition, since differences in MICs have been reported under nutrient-replete conditions after differing incubation times, 20 we tested the fungicidal activity of fluconazole under nutrient-depleted conditions at 24, 48, and 72 h. The total signals decreased slightly over the incubation times mainly due to reduced metabolic capacity of C. neoformans, but the overall types of inhibitions did not change (data not shown). Specifically, amphotericin B at 0.5 µg/mL exhibited potent fungicidal activity at each of the time points, and fluconazole at levels of 80 to 200 µg/mL again failed to show significant fungicidal activity compared with the control groups. The results indicated that the alamarBlue assay under nutrient deprivation conditions can identify potential fungicidal compounds such as amphotericin B against C. neoformans and successfully excludes static inhibitory compounds such as fluconazole.
Identification of 10058-F4 as a Putative Fungicidal Molecule against C. neoformans in a Screen of 1280 Compounds
We then used this nutrient-depleted fungicidal assay to screen the LOPAC library of 1280 compounds in a 96-well plate format to identify fungicidal compounds against C. neoformans. The screen was carried out using a compound concentration of 5 µM with a final concentration of 1% DMSO. The well-to-well variation was small for this assay as the S/B ratio was 11.5-fold and the Z′ factor was 0.894 based on eight positive and negative controls ( Fig. 2A ). A total of two primary hits were found from the screen, although one (S-(p-azidophenacyl)-glutathione, APA-SG) showed only minor inhibition. Because of the more potent activity and data on mammalian toxicity, 21 we then used a conventional C. neoformans assay 14 to confirm the fungicidal activity for the inhibitor, 10058-F4 (203 vs. 650 relative fluorescence units [RFU] for DMSO alone). Incubation of C. neoformans in ASN media in the presence of 40 µM 10058-F4 resulted in 70% killing of C. neoformans over 24 h, confirming the fungicidal activity of the compound ( Fig. 2B ). Furthermore, growth inhibitory effects of 10058-F4 were potentiated by the presence of the detergent sodium dodecyl sulfate (SDS), and the fungus showed slightly larger colony sizes in the presence of the osmotic stabilizer, mannitol, suggesting that surface-active agents may act together with 10058-F4 ( Fig. 2C ). However, we did not find potentiation of 10058-F4 with the membrane-active agent amphotericin B (data not shown), suggesting that this may be a detergent-specific effect. This demonstrated that this alamarBlue assay for C. neoformans under nutrient starvation conditions is useful to identify fungicidal compounds by screens of compound collections.
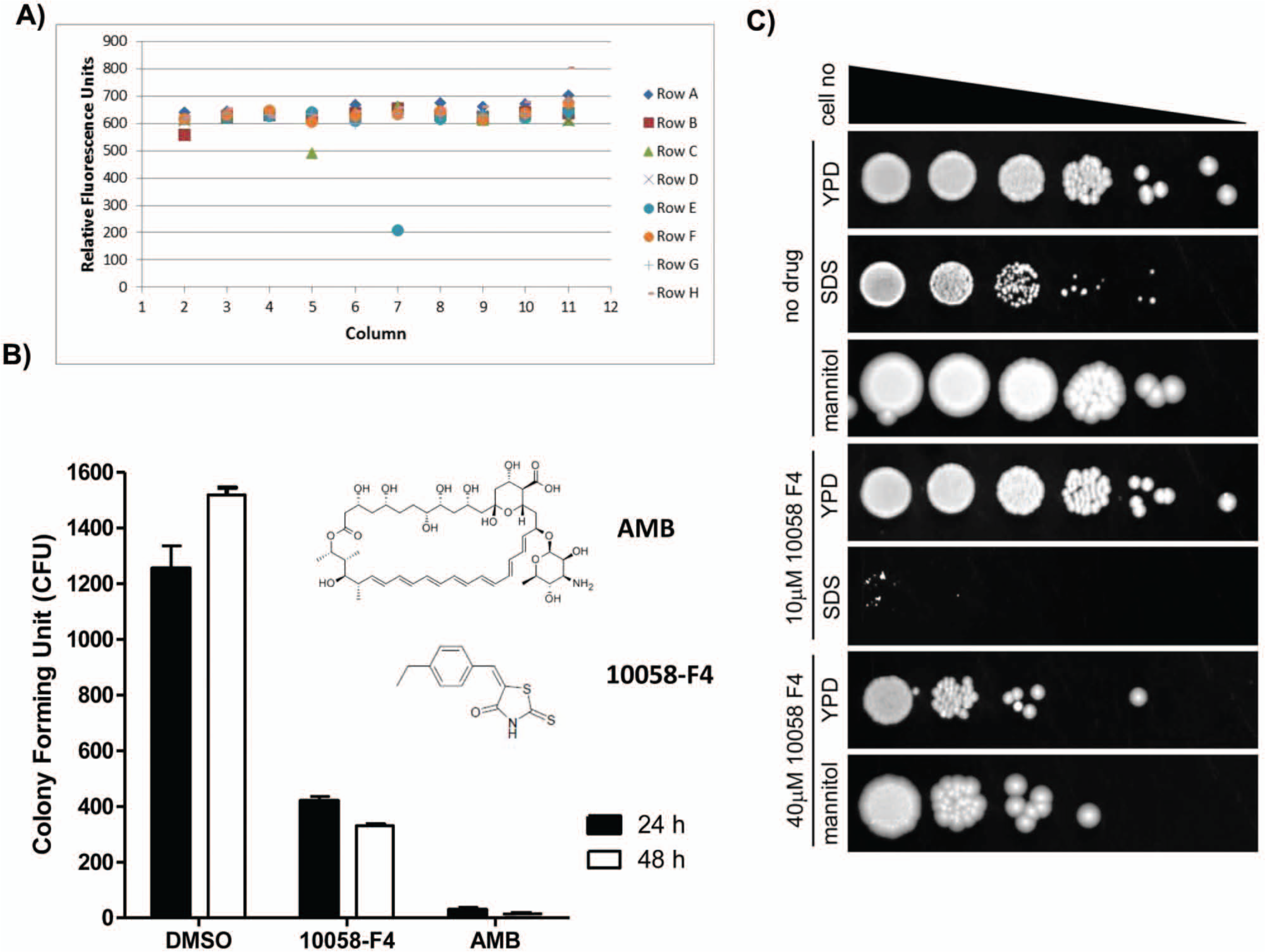
alamarBlue assay screening of the Library of Pharmacologically Active Compounds (LOPAC) in a 96-well format identifies 10058-F4 as a potential fungicidal lead compound. (
Assay Miniaturization to a 1536-Well Plate Format
To allow ultra-HTS of large compound collections using this assay to identify lead compounds, we miniaturized it to a 1536-well plate format. Briefly, the assay started with addition of 2.5 µL/well of culture medium with alamarBlue dye to 1536-well plates followed by addition of 23 nL compound in DMSO using a pintool workstation. The C. neoformans suspension was added at 2.5 µL/well to the plates that were incubated for 24, 48, or 72 h. The plates were then measured for fluorescent intensity in a plate reader. The results showed that the fluorescent signal increased with increases in cell densities from 2200, 4500, and 9100 cells/well, while the basal fluorescent signals did not change ( Fig. 3A ). The S/B ratios at 2200, 4500 and 9100 cells/well were 2.3-, 3.1-, and 3.5-fold, respectively ( Fig. 3B ). In addition, the concentration responses of amphotericin B were determined at different cell densities in the 1536-well plate assay. The IC50 values of amphotericin B at 2200, 4500, and 9100 cells/well were 0.35, 0.31, and 0.45 µM, respectively ( Fig. 3C ), but fluconazole again showed no significant inhibition under these conditions (data not shown). Thus, 4500 cells/well was selected as an optimal cell density for the experiment in a 1536-well plate format. Notably, the amphotericin B IC50 values were similar to the traditional MIC of 0.21 µM determined by a clinically validated Etest. 22 We also tested the effects of compound incubation time on the S/B ratio of the assay and potency of amphotericin B. The results showed that 24-h incubation had a better S/B ratio, and the potencies of amphotericin B were similar in the different compound incubation times. Using a cell seeding density of 4500 cells/well in the 1536-well format gave an S/B ratio of 4-fold and a Z′ factor of 0.79, demonstrating a robust assay suitable for HTS. To further compare the ability of this assay for assaying compound activities in 1536-well plates with that in 96-well plates, we selected an additional 13 compounds that are active against other fungi reported in the literature. We found that three of these compounds—zinc pyrithione, captan, and 10058-F4—exhibited activities against C. neoformans with IC50 values of 3 to 10 µM ( Table 1 ). Pyrrolnitrin showed a partial activity in both 96-well and 1536-well plate assays, while another nine compounds were not active ( Table 1 ). Taken together, the results demonstrate that this assay in the 1536-well plate format is useful for primary screens of large compound collections to identify lead fungicidal compounds.
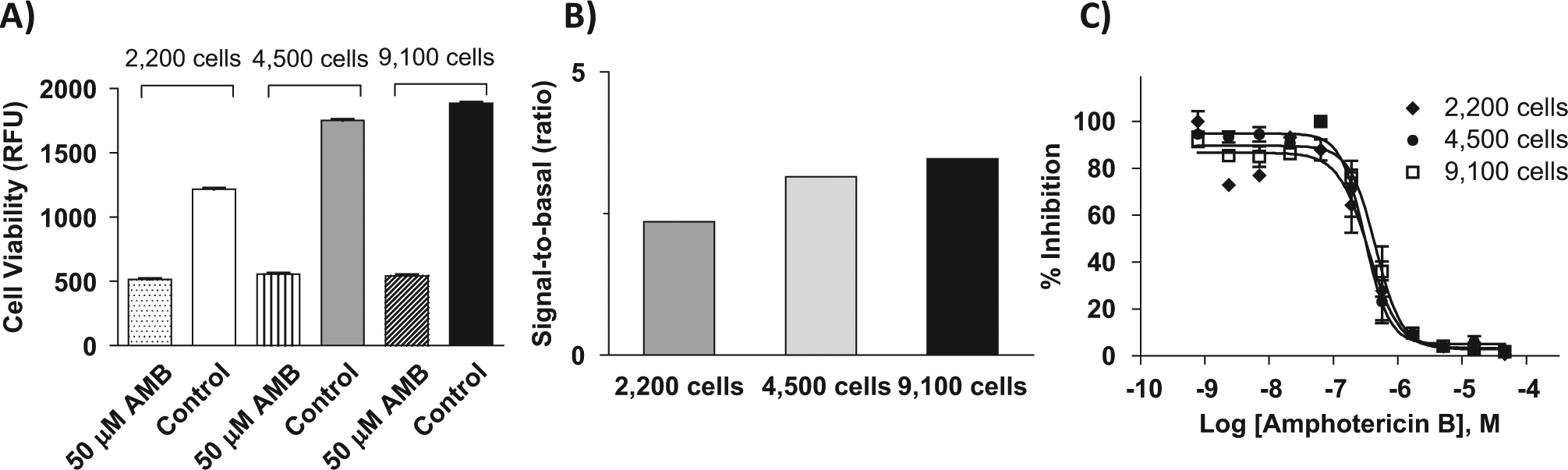
Cell density titration using a miniaturized 1536-well format. (
Comparison of Fungicidal Activities of 14 Compounds in 96- and 1536-well Formats.
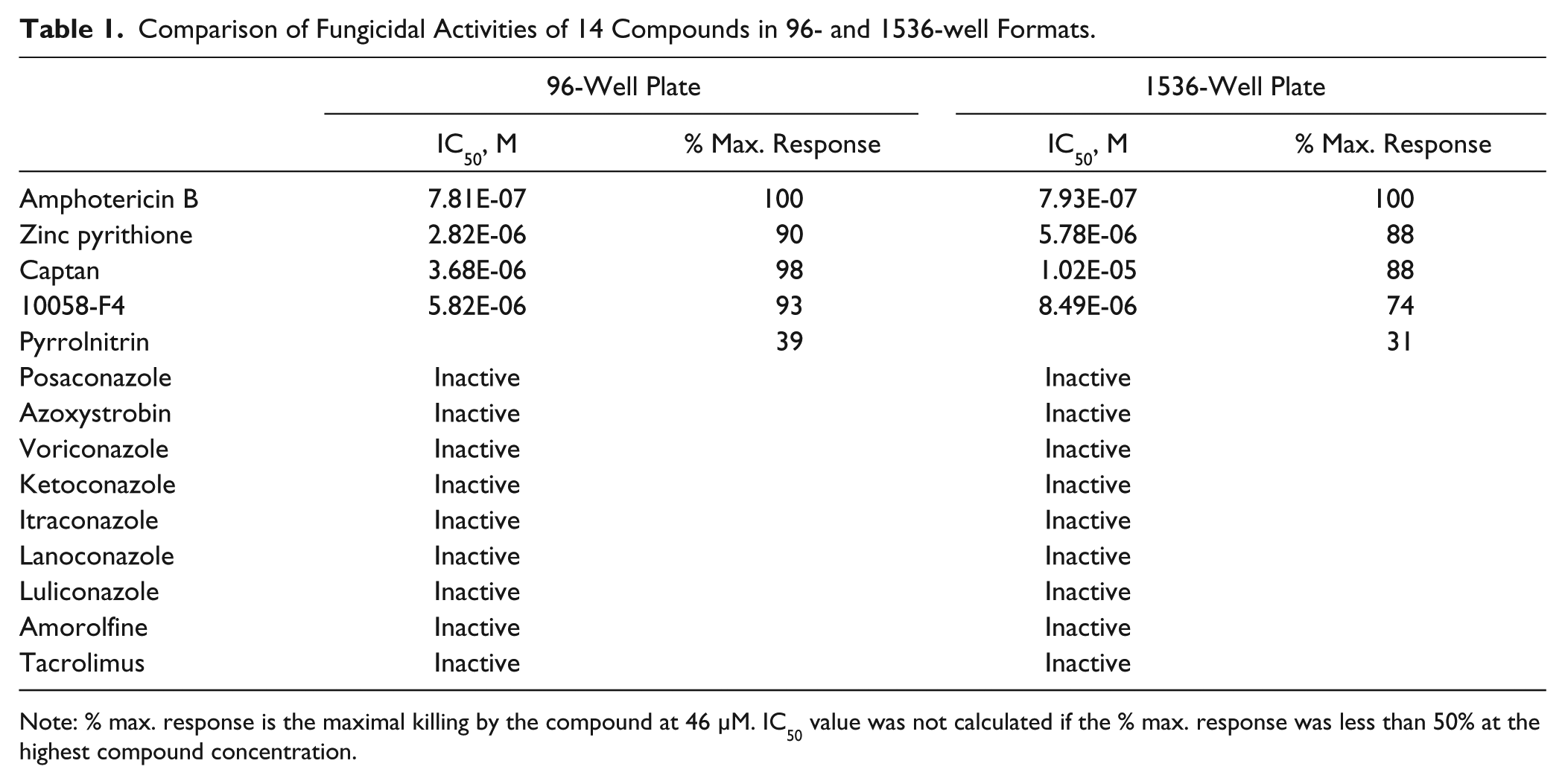
Note: % max. response is the maximal killing by the compound at 46 µM. IC50 value was not calculated if the % max. response was less than 50% at the highest compound concentration.
Discussion
To identify potential new anticryptococcal fungicidal compounds, we developed a fungicidal assay using the metabolic fluorescent sensor, alamarBlue, that measures residual metabolic activity after starvation to determine the viability of C. neoformans. It has been reported that the sensitivity of the cell viability assay using alamarBlue is much higher than other dyes such as 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT). 23 Our results showed that the alamarBlue assay is sensitive and reproducible, and it can be used to measure residual fungal metabolic activity up to 72 h in starvation media. Since C. neoformans does not grow under these starvation conditions, 13 reductions in metabolic activity in the presence of small molecules would be expected to be due to reductions in cell viability. Indeed, the fungistatic compound fluconazole failed to show reductions in the signal of this alamarBlue assay even at the compound concentration of 20× the MIC determined under nutrient-replete conditions for this strain. 5 However, the fungicidal compound amphotericin B displayed potent activity in this assay, confirming its ability to identify fungicidal compounds. Low levels of substrates such as glucose present at 20 mg/mL, 11 acetate 24 or lactate, 10 or nitrogenous sources that may be present at low concentrations in brain tissue and CSF from infected patients did not affect the respective properties of fungicidal versus fungistatic effects from either drug, suggesting similar inhibitory results under less severe nutrient-deprived conditions that could be present under varying severities of clinical infection.
Recent data suggest that the survival of C. neoformans under nutrient-deprived conditions may be a key aspect of cryptococcal virulence. For example, uptake of fungal cells by host macrophages results in induction of cellular programs suggestive of a starvation environment such as autophagy. 25 In addition, starvation-protective cellular programs have been implicated in virulence such as autophagy, 13 high-affinity glucose, 12 and iron 26 and copper transport. 15 However, such nutrient-deprived conditions may increase the difficulty of antifungal treatment of infections caused by C. neoformans. For example, antifungal agents such as fluconazole are highly active as growth inhibitors under nutrient-replete conditions, 5 but the agent results in little decrease in fungal organisms in glucose-deprived conditions, including the CSF of patients over time. In contrast, the fungicidal drug amphotericin B that is active under all nutrient conditions results in a time-dependent killing of the organism in patients. 8 Such data have prompted some investigators to reflect on the possible superiority of fungicidal over fungistatic antifungal agents in certain fungal infections such as cryptococcosis. 6 Thus, a fungicidal assay under nutrient-deprived conditions as described in the present study may provide a unique and alternative approach to identify lead compounds for drug discovery of C. neoformans that may not be identified through conventional compound screening methods. In addition, this assay has been miniaturized into a 1536-well plate format for ultra-HTS of large compound collections. Again, while fluconazole did not demonstrate fungicidal activity in the miniaturized assay, amphotericin had a similar IC50 value to that evaluated by a traditional MIC assay.
The present assay was used to screen a library of 1280 pharmacologically active compounds (LOPAC), which identified a potential chemical scaffold, 10058-F4. The fungicidal activity of 100058-F4 against C. neoformans was confirmed by conventional viability assays. While originally described as a c-Myc inhibitor, 27 recent evidence suggested its role in bromodomain-containing protein binding of acetylated lysine residues on histones 28 that may be responsible for its off-target effects. Indeed, the classical c-Myc regulatory pathway does not exist in yeast, and no homologs are present in C. neoformans. Thus, it may act at different protein or proteins in C. neoformans that remain to be elucidated. Further studies showed potentiation by the surface-active detergent SDS but little potentiation with the membrane-active agent amphotericin and little protection by the osmotic stabilizing agent, mannitol. This suggests that potentiation may have occurred due to the detergent’s effect as a permeation enhancer. 29 Interestingly, while 10058-F4 was found to be fungicidal in the low micromolar range, the compound induces cell cycle arrest of mammalian cells such as HL-60 cells at 60 to 100 µM, 30 suggesting a favorable fungicidal activity against C. neoformans. Furthermore, mouse studies have shown tolerable adverse effect profiles of the compound with peak levels in the 400-µM range. However, its half-life (t½) in these studies was previously reported to be 10 min in mice, 21 which indicates this compound is not suitable for testing in a mouse cryptococcal meningitis model. Nevertheless, the fungicidal activity against C. neoformans under nutrient limitation by 10058-F4 is of interest for further study as well as for further development for chemical optimization.
In conclusion, we have applied the use of alamarBlue dye to develop a sensitive fungicidal assay under nutrient starvation conditions for identifying fungicidal compounds against C. neoformans. The assay successfully distinguished the fungicidal activity of amphotericin B from the fungistatic effect of fluconazole and was successful in a test screen of the LOPAC library of 1280 compounds. We have also miniaturized this assay into a 1536-well plate format for ultra-HTS.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Intramural Research Program of the Therapeutics for Rare and Neglected Diseases, National Center for Advancing Translational Sciences, National Institutes of Health and the intramural program of the National Institute of Allergy and Infectious Diseases.