
Abstract
The three members of the organic cation transporter novel subfamily are known to be involved in interactions with xenobiotic compounds. These proteins are characterized by 12 transmembrane segments connected by nine short loops and two large hydrophilic loops. It has been recently pointed out that acetylcholine is a physiological substrate of OCTN1. Its transport could be involved in nonneuronal cholinergic functions. OCTN2 maintains the carnitine homeostasis, resulting from intestinal absorption, distribution to tissues, and renal excretion/reabsorption. OCTN3, identified only in mouse, mediates also carnitine transport. OCTN1 and OCTN2 are associated with several pathologies, such as inflammatory bowel disease, primary carnitine deficiency, diabetes, neurological disorders, and cancer, thus representing useful pharmacological targets. The function and interaction with drugs of OCTNs have been studied in intact cell systems and in proteoliposomes. The latter experimental model enables reduced interference from other transporters or enzyme pathways. Using proteoliposomes, the molecular bases of toxicity of some drugs have recently been revealed. Therefore, proteoliposomes represent a promising experimental tool suitable for large-scale molecular screening of interactions of OCTNs with chemicals regarding human health.
Introduction
Membrane transporters can be considered among the most important targets of administered drugs.1–5 Indeed, these proteins play roles both in drug delivery and interaction and thus are major determinants of the pharmacokinetic, safety, and efficacy profiles of drugs.2,6,7 On one hand, drug delivery in target tissues can be improved by the presence of suitable transport systems that increase bioavailability. On the other hand, drugs or toxic compounds can simply bind to transporters, leading to off-target interactions. If the binding constants are below their concentration in extracellular fluids, the drugs will interfere with the transport of physiological substrates, causing more or less severe side effects. In other cases, chemical compounds may exert beneficial effects inhibiting or stimulating transport systems, which are overexpressed or down-regulated, respectively, in pathological conditions such as cancer. Thus, understanding transport mechanisms and drug-transporter interactions is a very important task, contributing to predict bioavailability, therapeutic benefits, or side effects. 8 Some questions concerning drug-transporter interactions are fundamental for drug development, for example, which transporters are important in drug absorption, disposition, and interaction. Package inserts (PIs) of recently approved molecules have been surveyed to gain information on relationships of these molecules with transporters. 6 ABC efflux transporters are mentioned in 86% of the PIs citing transport systems. Organic anion transporting polypeptides are mentioned in 18%, breast cancer resistance protein in 15%, and organic anion transporters (OATs) in 8% of the PIs. Organic cation transporters (OCTs) have been also widely recognized to be involved in the interaction with cationic drugs, which represent about half of the molecules used in human therapy.9–11 Transporters of this subfamily are mentioned in 15% of the PIs. In addition to the OCTs, another subfamily of organic cation transporters has been more recently identified in higher organisms, the organic cation transporters novel (OCTN), including three members that are known to interact with several drugs. 12 Even though this novel family is not yet listed in any PIs, they are certainly involved in interactions and constitute potential pharmacologic targets because of their wide tissue distribution and well-defined association with several diffused pathologies.
In the following sections, an update will be given on (1) the little-known information on transporters’ structure and the physiological roles of each OCTN member, (2) the direct and indirect relationships of the transporters with human pathologies, (3) the perspective of assay developments to predict primary and/or off-target interactions with drugs and toxic compounds, and (4) the known mechanisms of interactions with drugs.
Structure Information and Physiological Roles of Each OCTN Member
The OCTNs constitute a subfamily of the SLC22 family, which, at higher classification, belongs to the major facilitator superfamily including more than 1000 identified members, which accounts for about 25% of all transporter families (http://www.tcdb.org/). 13
The OCTN subfamily members share several structural properties. The amino acid sequences of the three members in mice show 66% identity. 14 Hydropathy profiles of the three proteins show similarity in the 12 transmembrane segments, whereas the C-terminus and intracellular hydrophilic loop moieties are very different ( Fig. 1 ). In particular, the transmembrane segments show an average degree of identity of 68.9%, with the lowest value of 52% in the third transmembrane segment and the highest, 85.7%, in the first transmembrane segment. The large extracellular loop shows higher identity degree (71.2%) than the large intracellular loop (50.8%). The C-terminus shows the lowest identity degree, 16.3%. These data allow us to hypothesize that the specificity of the transporters is related mainly to the intracellular and extracellular loops, the C-terminus moiety, and the transmembrane segments 3, 7, 8, and 9 ( Fig. 1 ). The hydropathy profile of the transporters infers a structural asymmetry. A nucleotide binding motif located between transmembrane 4 and 5 on the intracellular side has been predicted.14–17 No data on tertiary structure are available so far, with the exception of an homology structural model of OCTN1. 18 However, this model lacks 142 N-terminal amino acids, including the large extracellular hydrophilic loop. Few structures of cation transporters of bacteria have been resolved, which do not contain large hydrophilic loops and therefore are not suitable templates for homology modeling of OCTNs.19–21 The large loop of OCTNs contains four Cys residues. At least one of these residues can be targeted by externally added hydrophilic SH reagents, in agreement with the predicted topology.15–17,22 Cys residues in the hydrophilic loop and/or in the transmembrane moiety could come in close proximity and form disulphide bonds. 14 Functional data combined with inhibition studies gave information on molecular interactions occurring among the OCTN1 or OCTN2 proteins and the substrates ( Fig. 2 ). In both cases, the substrate-binding sites can be represented by models in which three subsites interact with the main functional groups of the substrates (i.e., the ammonium group, the carboxyl/hydroxyl group, and, in the case of OCTN2, a sodium site). This reflects the different ability of the transporters in interacting with the substrates/inhibitors. However, the resolution of at least one of the OCTN subfamily member’s structure by x-ray crystallography is mandatory to obtain a detailed view of the structure-function relationships.
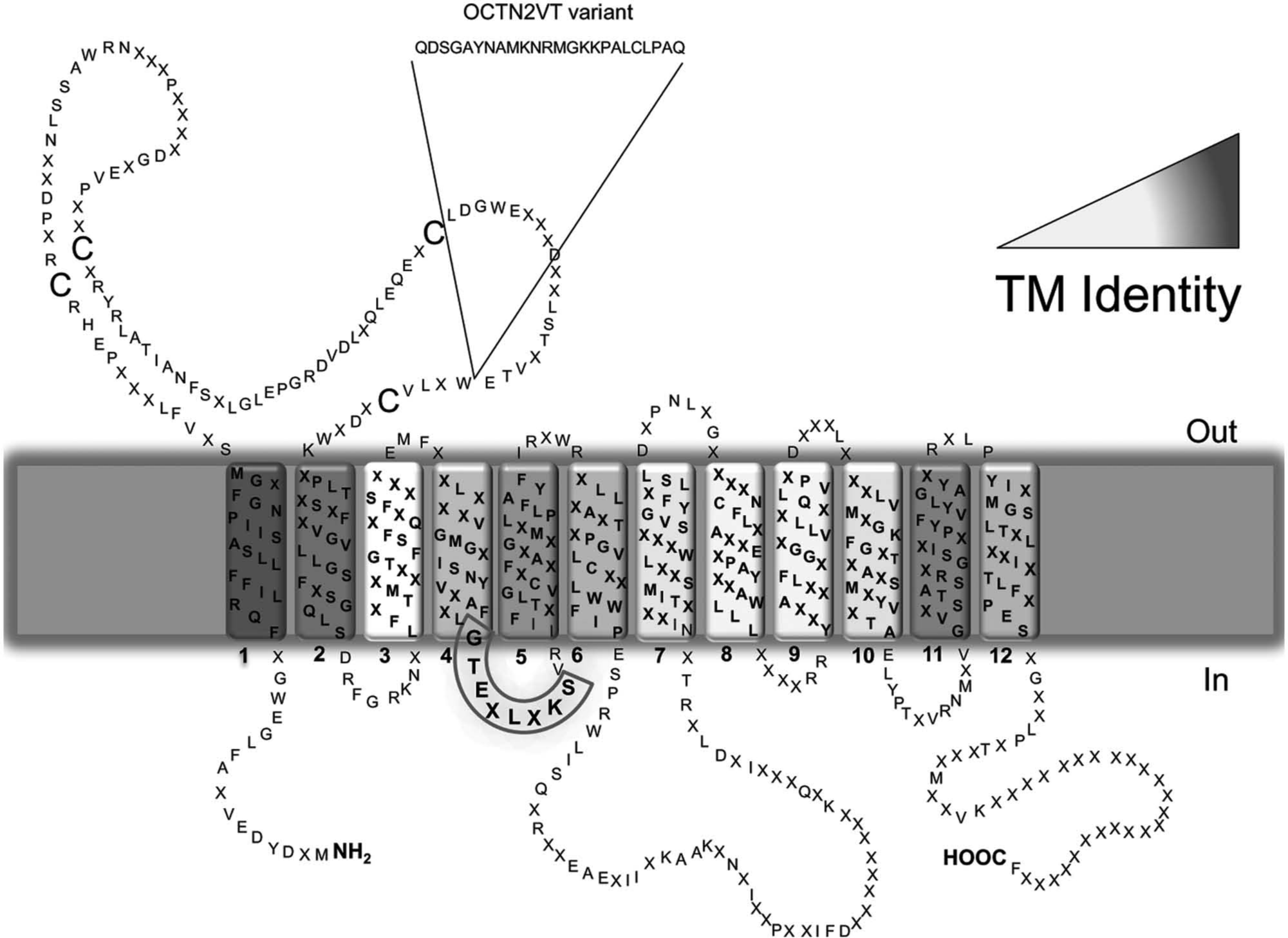
Topology model of the organic cation transporters novel (OCTN) subfamily. The model was created on the basis of the sequences of hOCTN1, hOCTN2, and mOCTN3 using the TM-PRED prediction software (output format: min 17, max 29) and modified considering the established criteria for transmembrane proteins. The cell membrane is represented in gray with the indication of the extracellular (out) and intracellular (in) sides. Transmembrane (TM) segments of the transporters are numbered (1–12), and the extracellular loop together with intracellular C-ter and N-ter are indicated. Residues that are conserved in the three members are indicated by the respective one letter amino acid code; residues that are not conserved are indicated by X. The identity among the TM segments is highlighted by the grayscale color reported. The insert of 24 amino acids of the hOCTN2VT variant is indicated. The nucleotide binding motif is highlighted by a gray box between TM4 and TM5.
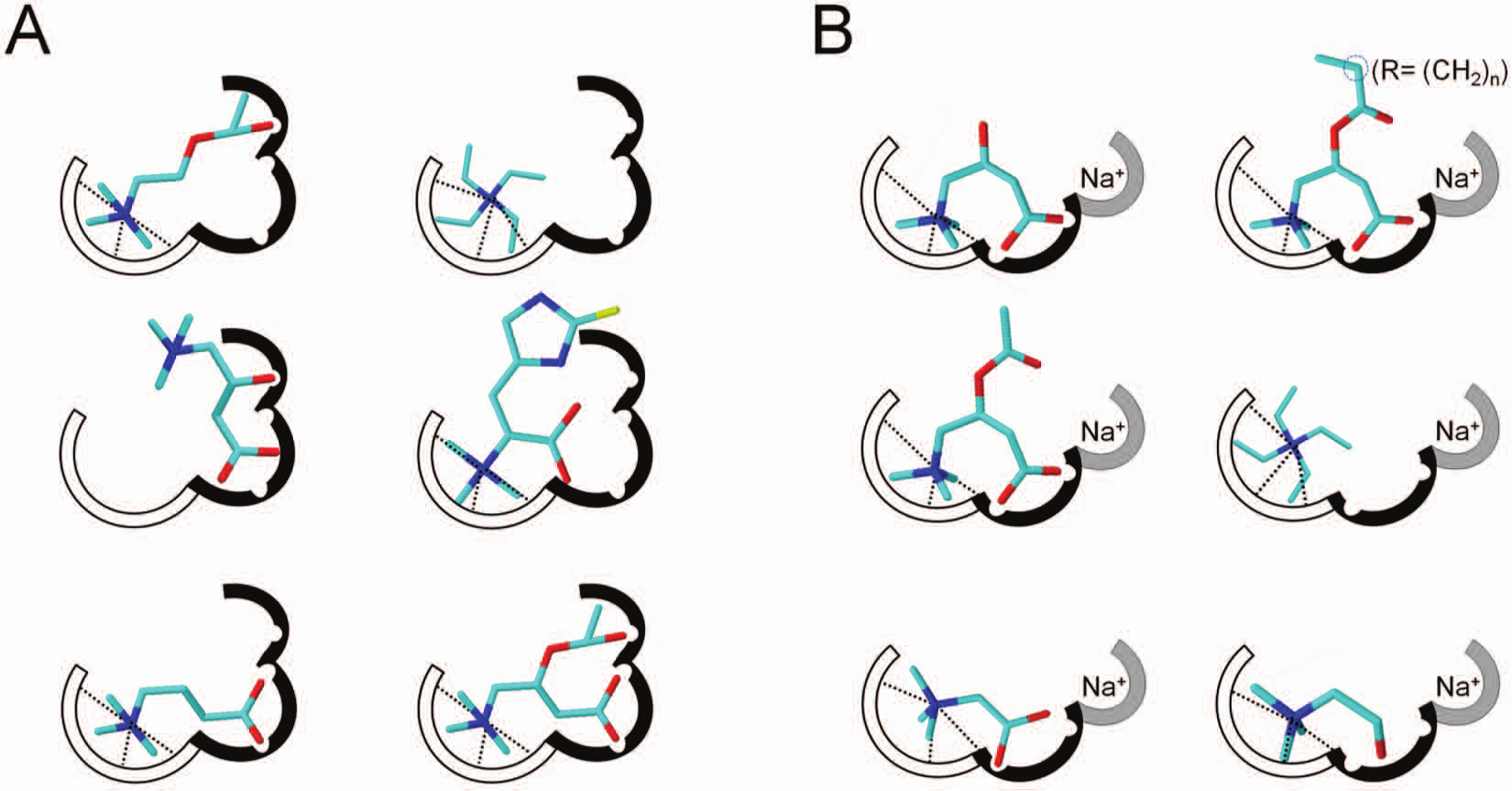
Binding sites of OCTN1 and OCTN2. The binding sites are composed of three sub-sites. (
OCTN1
Despite the numerous studies performed so far, the physiological role of OCTN1 is still not clear, even though acetylcholine was recently found as a physiological substrate.24–29 The expression profile of the transporter has been extensively investigated: OCTN1 is strongly expressed in renal epithelium and, at a lower level, in trachea, bone marrow, liver, skeletal muscle, prostate, lung, pancreas, intestine, placenta, heart, uterus, spleen, spinal cord, and neurons.24,26 -28 OCTN1 expression has also been described in different cancer cell lines.24,30 Only indirect evidence suggests an apical sublocalization of OCTN1 in kidney, airway, placenta, intestine, corneal, and conjunctival epithelial cells.28,31 -34 Although most of the experimental evidence shows OCTN1 in the plasma membrane, its localization in mitochondria has also been suggested.35,36 In intact cell systems, it was found that the transporter mainly catalyzes the transport of organic cations, whose prototype is tetraethylamonium (TEA).24,28 Moreover, studies of inhibition by pharmacologic compounds (see the subsection “Interactions of OCTNs with Drugs and Xenobiotics Studied in Cell Systems”) suggested that this transporter is involved in the excretion of cationic xenobiotics from renal epithelium.24,25 It was also found that OCTN1 could catalyze, at a very low rate, carnitine transport.25 -27 More recently, it has been reported that OCTN1 transports a mushroom metabolite, the antioxidant ergothioneine (ET), whose precise physiological role in human physiology and the consequences of its deficiency are still unclear.37 -39 Very recently, hOCTN1 has been overexpressed in Escherichia coli and reconstituted in liposomes ( Fig. 3 ).14,16 Some data were in agreement with cell system studies, and novel properties emerged from proteoliposomes.16,26,29,40-42 It was clarified that carnitine and ET are transported with much lower efficiency with respect to TEA ( Table 1 ). Moreover, transport and/or interaction of acetylcholine, acetylcarnitine, and γ-butyrobetaine have been revealed. Competitive interactions were found among TEA and acetylcholine, acetylcarnitine, and γ-butyrobetaine, whereas noncompetitive interaction was exhibited by carnitine on TEA transport. 16 These data fit with the substrate-binding site depicted in Figure 2A . The oxygen binding pockets may be constituted by different amino acid residues able to form hydrogen bonds, and the ammonium binding site may be constituted by tryptophan residues able to form cation-π interactions. This correlates well with features of bacterial carnitine transporter structures.20,21 Acetylcholine, acetylcarnitine, and γ-butyrobetaine fit with at least two subsites. In the case of carnitine and ET, a different orientation of the molecules prevents the overlap with the cation site in agreement with the noncompetitive interaction with TEA. 16
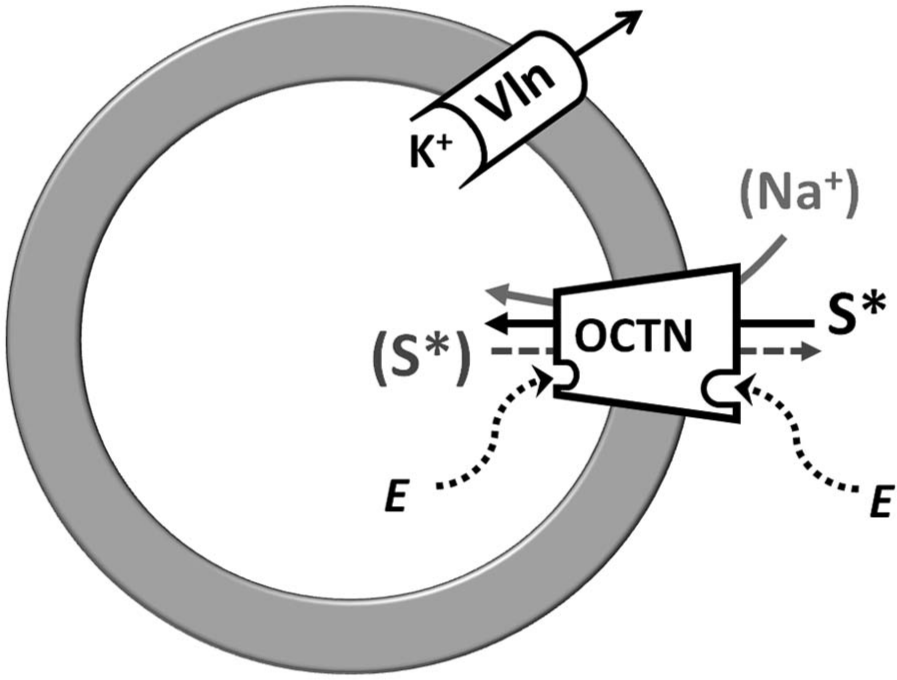
Description of the experimental system of proteoliposome and screening example. Proteoliposome membrane is represented by a ring. A generic organic cation transporters novel (OCTN) transporter is asymmetrically inserted in the membrane vesicles as in cell membrane. The radioactive labeled substrate, S*, is transported into the vesicles. The dot arrow indicates the possibility of a labeled or unlabeled countersubstrate S (in gray); the light gray arrow shows the case of uptake Na+ dependent. “E” inside and outside of the proteoliposome compartments represents a generic effector (endogenous molecules, drugs, or xenobiotics) stimulating or inhibiting OCTN transporter activity. To generate membrane potential, the ionophore valinomycin (Vln) is inserted in membrane mediating efflux of intraliposomal K+. The proteoliposome system allows preloading of vesicles with labeled substrate, by transporter-mediated equilibration, following its efflux from proteoliposome (S*).
OCTN transporters: substrate specificity, roles in cell physiology, association of altered function with pathologies.
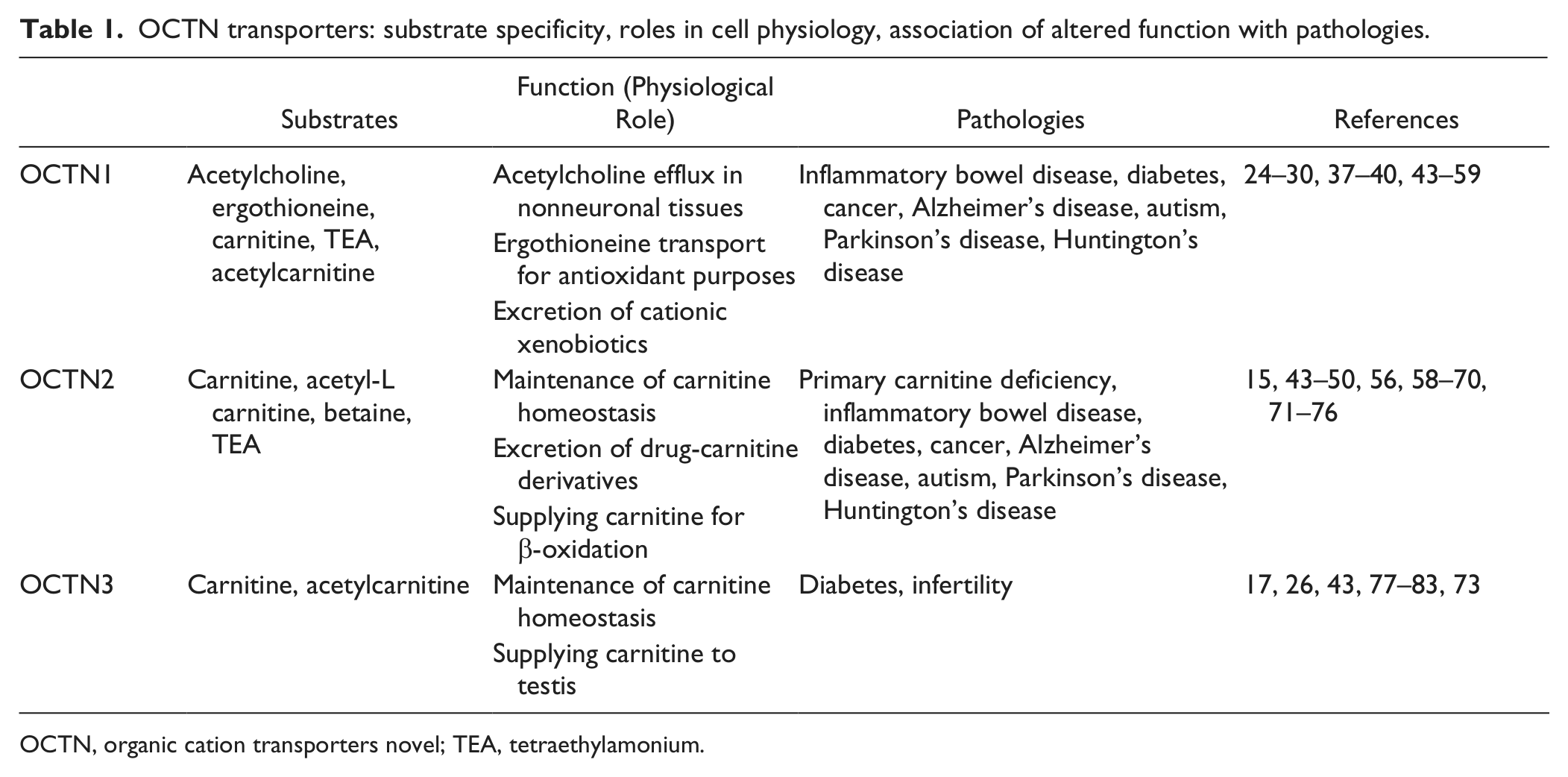
OCTN, organic cation transporters novel; TEA, tetraethylamonium.
hOCTN1 catalyzes a bidirectional transport of acetylcholine.29,40 This emerged function is consistent with the nonneuronal cholinergic system, a novel metabolic pathway recently described in many tissues.84,85 Indeed, it was shown that both epithelial (skin, airways, intestine) and immune cells (lymphocytes) express all components of the cholinergic system independent of neuronal innervations. In these tissues, acetylcholine does not behave as a neurotransmitter but is involved in the regulation of basic cell functions such as proliferation, differentiation, organization of the cytoskeleton, local release of mediators (for example, NO, proinflammatory cytokines), locomotion, secretion, and ciliary activity.84,85 hOCTN1 should catalyze the release of the neosynthesized acetylcholine from cells to activate the receptor cascade in autocrine and paracrine mode ( Fig. 4 ). Very interestingly, acetylcholine controls inflammatory processes in epithelia via the nonneuronal cholinergic system. This anti-inflammatory role was described in 2003 for innervated tissues and hypothesized for nonnervous cells. 86 The transporter is more efficient in catalyzing efflux toward the extracellular environment than uptake of acetylcholine, which is depressed by extracellular Na+. 40 Transport is stimulated by intraliposomal (intracellular) adenosine triphosphate (ATP), according to the presence of a nucleotide binding domain in the structure of the transporter and in line with the structural asymmetry ( Fig. 1 ). The role of hOCTN1 in the nonneuronal cholinergic system opens new perspectives in designing specific therapies for human pathologies caused by alterations of acetylcholine metabolism. In particular, in case of a lack/decrease of acetylcholine synthesis, a supply of acetylcarnitine might help to restore this alteration according to the cell metabolic network, which involves the enzyme carnitine acetyltransferase ( Fig. 4 ; and see the section “Involvement of OCTNs in Human Pathology”). In the heart, OCTN1 may affect the absorption/elimination of organic cationic drugs playing potential roles in the modulation of myocardial bioenergetics, valvular function, and acetylcholine generation for parasympathetic vagal innervations of the cardiac conduction system. 87 An evolutionary conserved feature of the SLC22A family is the scarce specificity toward several exogenous compounds. 12 Cationic drugs should interact with the cationic site of OCTN1 ( Fig. 2A ) owing to the possibility that this subsite can host molecules with different sizes. 30
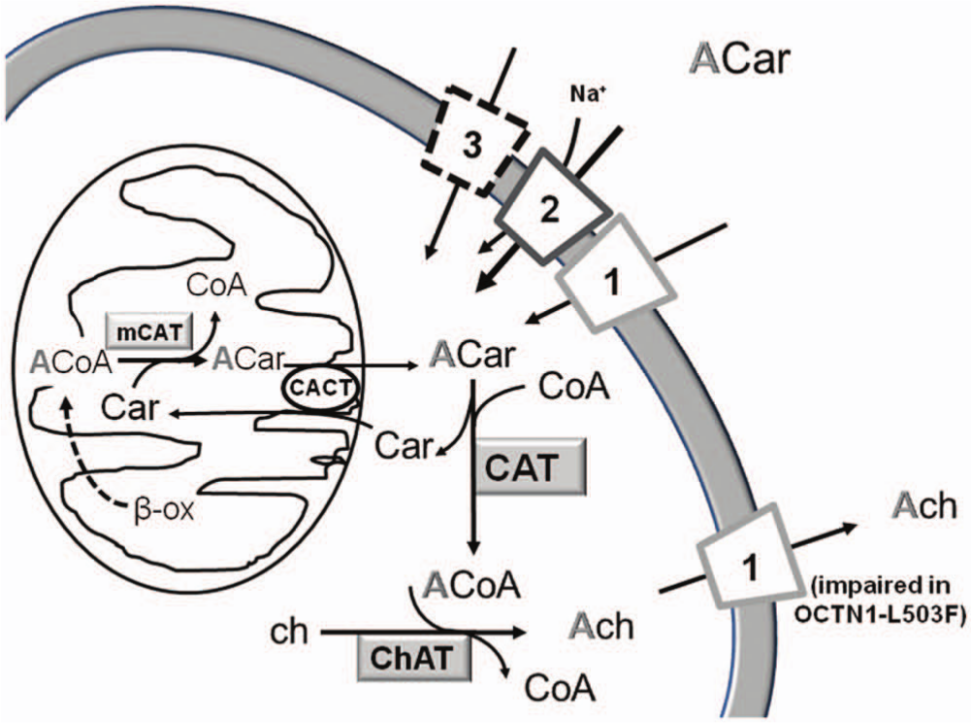
Metabolism of acetylcarnitine and acetylcholine in a representative human nonneuronal cell. The figure depicts the OCTN1, OCTN2, and OCTN3 proteins indicated by the respective numbers. Dotted lines represent still undefined transporter function. Cytosolic acetyl-carnitine (ACar) is provided either by extracellular pool via OCTN1, 2, and possibly 3 or by endogenous mitochondrial production. Cytosolic and mitochondrial pathways are depicted, and enzymes with metabolites are indicated: cytosolic carnitine acetyl transferase (CAT), mitochondrial carnitine acetyl transferase (mCAT), choline acetyl transferase (ChAT), acetylCoA (ACoA), acetylcholine (Ach), and choline (ch). Synthesized Ach is released by OCTN1. The pathway of Ach synthesis from Acar is also active in neural cells (see the text for details).
Long-term hormonal regulation of OCTN1 has been described: testosterone, differently by estrogen, may regulate apical OCTN1 expression.88,89
OCTN2
OCTN2 is a plasma membrane carnitine transporter playing the function of providing cells with carnitine, an essential cofactor for the mitochondrial β-oxidation of fatty acids ( Fig. 5 ). Carnitine must be supplied by diet, because the endogenous synthesis is insufficient for the body requirement. 43 OCTN2 expression is not limited to polarized cells of intestine, kidney, placenta, and mammary gland but has been found in many other tissues such as liver, heart, testis, skeletal muscle, and brain, guaranteeing carnitine absorption and distribution within the entire organism.26,60,61 Thus, the physiological and the pathological relevance of OCTN2 is in its tissue-specific functions ( Fig. 5 ; Table 1 ). For example, although in intestine or kidney, OCTN2 mediates the absorption or reabsorption, respectively, of carnitine for maintaining a constant plasma carnitine level (50 µM), in the distal epididymis, OCTN2 is important for accumulating carnitine inside the cells ( Fig. 5 ). In this district, carnitine reaches the highest level of the body.43,52,53 Functional data obtained in cells, in membrane vesicles, or in proteoliposomes demonstrated that this transport system has a well-defined substrate specificity for carnitine and its derivatives, whose transport is coupled to sodium ( Table 1 ).15,26,43,60,61,64–67,77,78,90 The structural requirements for interaction with the transporter are a trisubstituted charged amino group (ammonium) and a carboxylic group ( Fig. 2B ); the distance among these groups is not important. 15 Na+ interacts very specifically with its subsite ( Fig. 2B ); Li+ or K+ is not tolerated. 15 Different transport modes have been described for OCTN2. OCTN2 catalyzes Na+-dependent symport or antiport.43,65,91 The Na+-dependent antiport of carnitine (with a 1:1 stoichiometry of transport) with carnitine analogues in kidney allows excretion of acyl groups produced either physiologically or as drug catabolites, such as pivaloylcarnitine and valproylcarnitine with simultaneous carnitine reabsorption ( Fig. 5 ; Table 1 ).15,22,43,92–95 The Km for carnitine shows differences depending on the tissue. The value ranging from 8 to 80 µM indicates a high substrate affinity. 43 Using proteoliposomes, the Km for the internal site has been measured (1.2 mM); it is much higher than that of the external one, indicating a functional asymmetry that correlates with the hydropathy profile ( Fig. 1 ). 15 A tissue-specific function of the transporter could be corroborated by the existence of physiological variants of OCTN2 or by different kinds of regulation of the transporter. Besides pathological variants (see the section “Involvement of OCTNs in Human Pathology”), only one splice variant, OCTN2VT, has been described, so far; it is characterized by the insertion of 24 amino acids in the first extracellular loop ( Fig. 1 ). This variant has been localized in the endoplasmic reticulum, where it can play a role in acylcarnitine trafficking ( Fig. 5 ).43,96
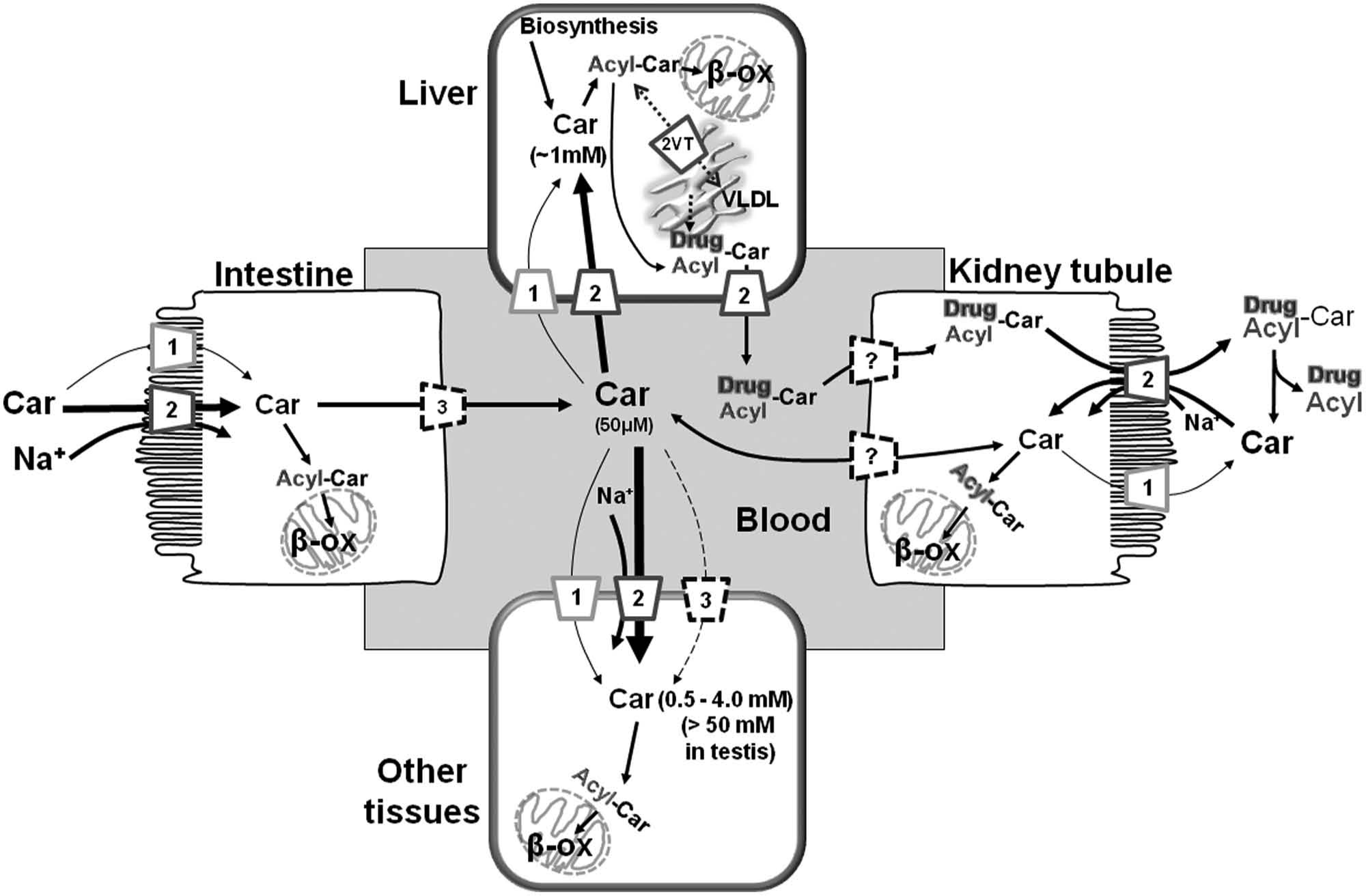
The carnitine homeostasis and transporter network. Interplay among epithelial polarized and other cells. Intestine on the left, kidney on the right side; liver on the top, and other tissues on the bottom. OCTN1, OCTN2, OCTN3, and the splice variant OCTN2VT are indicated by the respective numbers. Dotted lines represent still undefined transporter function. The question mark represents undefined transporter potentially involved in transport (OCTN3 or CT2/Flipt). Intestine lumen, blood, and tubular lumen are indicated. Mitochondria and endoplasmic reticulum are depicted by sketches. Simplified cytosolic and mitochondrial pathways are depicted: synthesis of acylcarnitines (Acyl-Car) from carnitine (Car) and transport in mitochondria mediated by enzymes and transporter of the carnitine cycle, AcylCoA synthetase, carnitine palmitoyl transferases, and carnitine/acylcarnitine transporter; mitochondrial oxidation (β-ox); very-low-density lipoprotein synthesis (VLDL). Formation of carnitine derivatives with drug metabolites are indicated (Drug-Car).
Regulation of OCTN2 by protein-protein interaction or transcription/translation modulation has been reported.97,98 The activation of peroxisome proliferator activated receptor α (PPARα) was also described to up-regulate OCTN2 expression; this effect can be mediated by endogenous ligands and also by drugs such as fibrates. 99 Hormonal regulation exerted by insulin has been described. 100 Intestinal epithelial OCTN2 expression is increased by inflammation. These findings suggest that OCTN2 may participate in restoration of intestinal homeostasis under conditions of inflammation-associated stress. Interferon-γ and tumor necrosis factor–α increased intestinal OCTN2 expression. 101 OCTN2 is an estrogen-dependent gene regulated via a newly identified intronic estrogenic responsive element. 102
OCTN3
OCTN3 was first characterized in HEK293 cells. 26 High-level expression of OCTN3 was found in mice testis, whereas OCTN3 was weakly expressed in liver peroxisomes and brain.77,79,103,104 Recently, OCTN3 has been localized in myoepithelial at all stages of breast development. 80 Finally, OCTN3 is also expressed in the apical membrane of kidney cells and in the basolateral membrane of intestine cells.78,81 This arrangement can mediate the intestinal absorption of carnitine from the diet: once carnitine is concentrated into the enterocytes by the apical OCTN2, it reaches the blood via the basolateral OCTN3 ( Fig. 5 ). 43
OCTN3 catalyzes sodium-independent carnitine transport, whereas it is not able to transport cations such as TEA ( Table 1 ).26,81 Studies performed preloading brush-border membrane vesicles with L-carnitine, betaine, γ-butyrobetaine, or TEA showed that both L-carnitine and betaine trans-stimulated carnitine uptake.77,78,81 mOCTN3 was recently overexpressed in E. coli and reconstituted in liposomes. 17 mOCTN3 is stimulated by intraliposomal (intracellular) ATP according to the presence of a nucleotide binding motif in the intracellular side ( Fig. 1 ). 14 The uptake of carnitine was inhibited by substrate analogues and by the neurotransmitter γ-aminobutyric acid. This molecule has been shown specifically to inhibit carnitine uptake in the blood-brain barrier, shedding new light on the possibility that OCTN3 could be one of the players in the carnitine network in the brain ( Figs. 4 and 5 ).17,79 In rat astrocytes, a PPARα activator has been found to stimulate carnitine uptake mediated by OCTN3. 82 Moreover, an increased peroxisomal localization of OCTN3 was found upon PPARα stimulation, indicating a role as peroxisomal transporter involved in fatty acid metabolism in astrocytes.82,105 Even though the OCTN3 protein has been identified in human skin fibroblasts using OCTN3-specific antibody, no human homolog of the murine OCTN3 sequences could be identified by bioinformatics (not shown). 105 This may indicate that either OCTN3 is not present in humans or it corresponds to a splice variant of other genes.
Involvement of OCTNs in Human Pathology
OCTN family members play pivotal roles in maintaining cell homeostasis. Thus, OCTNs’ defects are associated with different pathologies, exhibiting a wide range of severity ( Table 1 ). In particular, some of these pathologies are caused by derangements of carnitine homeostasis ( Fig. 5 ) and are known as primary carnitine deficiency (PCD). Inflammatory bowel diseases (IBDs) have been quite clearly associated, at different levels, to genetic mutations of OCTNs.41,106 It is noteworthy that diabetes, Alzheimer’s disease, and male infertility have been also associated with carnitine homeostasis impairments.62,107,108 Novel findings link cancer development/progression to fatty acids and carnitine metabolism alterations.68,109
Primary Carnitine Deficiency Is Caused by Defects of OCTN2
PCD (OMIM No. 212140) is a metabolic autosomal recessive disorder caused by defects of OCTN2 with well-characterized symptomatology.69,70,110 However, PCD should be differentiated from other causes of carnitine deficiency.64,111,112 Carnitine palmitoyl transferase 2 and carnitine/acylcarnitine transporter (CACT) deficiencies cause secondary carnitine deficiency.113–115 More than 100 different mutations in the human gene have been associated with pathology onset (http://www.hgmd.cf.ac.uk/ac/index.php; http://www.arup.utah.edu/database/OCTN2/OCTN2_display.php).43,69,70 In 2004, a mutation in the OCTN2 gene (R282D) was associated with sudden infant death syndrome. 116 It emerged that even though the link to genetic and phenotypic mutations of OCTN2 is quite clear, there is wide variability in the severity of manifestation which can be related to epigenetic or exogenous factors. The PCD patients respond to carnitine supplementation (50–400 mg/kg/d) if started before irreversible organ damages occur. 69 The therapy efficacy can be explained by the capacity of other transporters such as OCTN1 to transport carnitine, even though with lower efficiency with respect to OCTN2 ( Fig. 5 ; Table 1 ).26,29 Because PPARα positively regulates OCTN2 expression, its agonists might be potential candidates for treatment of those forms of PCD characterized by decreased expression of OCTN2. 110 Another transporter able to catalyze carnitine membrane translocation, CT2/Flipt2, might also substitute and/or synergize with OCTN2 in mediating carnitine uptake.83,117
IBD: Association with OCTN1 and OCTN2 Defects
IBDs are both environmental and genetic illnesses. More than 100 genes or loci involved in the regulation of innate or acquired immune response have been identified and linked to IBDs, which include Crohn’s disease and ulcerative colitis.118,119 Chronic inflammatory diseases also include celiac disease. 119 Increasing evidence strongly suggests that IBDs result from an inappropriate inflammatory response to intestinal microbes in a genetically susceptible host.118,9119 Indeed, the gut host microbiota has the potential to exert pro- and anti-inflammatory responses and, hence, to maintain the correct functioning of the host immune system.120–122 Dietary changes, drug treatments, and infections can alter gut microbiota composition, referred to as dysbiosis, which is a hallmark of IBD patients. 122 Very recently, the involvement of specific vitamin-based bacterial metabolites in host immune response activation has been proposed. 123 These findings suggest the existence of metabolic signatures of microbial infections stimulating immunosurveillance.44,121,122 The inflammatory diseases are comparable to autoimmune disorders.121,122 In line with this, many IBD patients show rheumatoid arthritis. 124 Some transporters involved in control of water and ion fluxes have been associated with IBDs. 44 IBDs and Crohn’s disease, in particular, are associated with mutations of genes encoding SLC22A4 and SLC22A5 (i.e., OCTN1 and OCTN2, respectively; Table 1 ). The mutation in the SLA22A5 gene is located in the promoter region; it is a transversion 270G→C, which disrupts a heat shock binding element. This mutation causes carnitine transport impairment. Recently, a link between carnitine uptake derangement and clinic phenotype of Crohn’s disease has been proposed. 45 Carnitine, in fact, has been successfully used to treat gut inflammation due to Crohn’s disease.45,46 In the case of the SLC22A4 gene, a missense mutation causes the amino acid substitution L503F. 41 So far, the mechanism underlying this phenotype is not clear; indeed, defects of ET, proposed as OCTN1 substrate, are not in line with Crohn’s phenotype. 39 Defects in acetylcholine transport will have more strict correlation with the disease (see the subsection “OCTN1” and Fig. 4 ). Indeed, acetylcholine has an anti-inflammatory action in neuronal and nonneuronal cells as well. Very interestingly, the L503F substitution in hOCTN1 causes severe impairment of acetylcholine release. 29 An important role in IBD treatment can be played by acetylcarnitine, which could be responsible for boosting acetylcholine production. Acetylcarnitine, physiologically provided by mitochondria, might also be exogenously supplied by means of the transport function of OCTNs ( Fig. 4 ).16,29,43,47,125,126
OCTNs and Diabetes
Besides the well-documented role in fatty acid metabolism, carnitine is essential for carbohydrate metabolism regulation. In humans, less than 25% of carnitine is synthesized endogenously in kidney, liver, and brain; the remaining fraction derives from diet.127,128 Besides the already described carnitine deficiencies, alterations in carnitine metabolism have been recently linked also to diabetes mellitus and obesity. 129 Carnitine, in fact, plays the role of shuttling acyl and especially acetyl groups from the mitochondrial matrix to liver cytosol and other tissues ( Figs. 4 and 5 ). This causes an increase of glucose metabolism according to the Randle hypothesis on the glucose–fatty acid cycle.48–50,129 Moreover, it has been reported for several years that lipid-induced insulin resistance is a hallmark for diabetes mellitus patients.49,50,108 Carnitine, in fact, would facilitate the export of fatty acids from tissues as acylcarnitines, alleviating lipid-induced insulin resistance and thus stimulating glucose utilization ( Fig. 4 ). 129
OCTNs and Cancer
Cancer cells shift from aerobic to anaerobic ATP production, bypassing the mitochondrial function; this phenomenon is known as the Warburg effect. 130 In other words, the energy in cancer cells comes mainly from anaerobic glycolysis and amino acid oxidation rather than fatty acid β-oxidation in mitochondria. The fatty acid oxidation is regulated in cells, among other factors, by carnitine availability; in this scenario, carnitine homeostasis has been suggested to play a regulatory function in the metabolic switch observed in cancer.68,109 Recently, down-regulation of OCTN2 expression in several epithelial cancer cell lines has been reported. 68
A link between OCTN1 and cancer has been also proposed. 30 The transporter seems to be associated with sporadic colorectal cancer in early age, and its polymorphisms, such as L503F, may help to predict malignant progression of the disease in IBD patients. 51 OCTN1 has been found to be overexpressed in tumors, being involved in the uptake of anticancer drugs. 30 The most effective drugs are positively charged, thus being potential substrates of OCTN1 ( Fig. 2 ). 52
OCTN’s Relationship with Alzheimer’s Disease and Neurological Disorders
The brain is the most metabolically active organ in the body. 53,107 One of the hallmarks of a normal aging process is the decay of mitochondrial function with consequent impairment of energy production, cell injury, and death.53,54 The decline in brain function is typical of neurological diseases, such as Alzheimer’s and Parkinson’s diseases. 54 In this scenario, any metabolic intervention that could improve mitochondrial function and ATP production may have great importance. Carnitine and acetylcarnitine may play these roles.53,55 Acetylcarnitine, entering into cells via OCTN2 or OCTN1, may have the beneficial effects of stimulating the Krebs cycle for ATP production.79,55,56 Most research, however, has focused on the effects of carnitine and acetylcarnitine on dementia.53,113,131 In humans, prolonged treatment of Alzheimer’s disease patients with 1.5 to 3.0 g/d of acetylcarnitine improved both clinical scales and psychometric tests. 131 Acetylcarnitine would reduce reactive oxygen species–induced neurotoxicity. 132 Notwithstanding, Alzheimer’s disease is a multifactorial pathology, and the real effectiveness of acetylcarnitine supplementations is still controversial. 132 In line with the role of neurological disorders, acetylcarnitine exerts beneficial effects in neurologic postischemia due to boosting of aerobic energy metabolism together with inhibition of drug-induced lactic acidosis.55,133 It is noteworthy that autism spectrum disorders (ASDs) have been very recently associated with mitochondrial dysfunction. In this context, carnitine supplementation seems to be helpful for children with ASD. 57 Recently, an improvement of cognitive function has been observed upon carnitine supplementation in Huntington’s disease patients. 58
Furthermore, regarding neurotransmission modulation, acetylcarnitine has been shown to be involved in neurotransmitter synthesis and function ( Fig. 4 ). 55 Ultimately, it has to be highlighted that in the brain, acetylcarnitine may act as a transcription factor of signal transduction and inflammatory proteins, whereas outside of the brain, acetylcarnitine, via histone acetylation/deacetylation mechanism, is also involved in regulation of genes linked to energy metabolism.71,72
Other Potential Involvement of OCTNs in Human Pathology
The OCTN subfamily members are involved also in other cellular functions linked to carnitine metabolism, such as energy production in the fetus and maturation of spermatozoa. Carnitine, in fact, is an essential nutrient for newborns.73,80,125 L-carnitine is avidly absorbed via OCTN2 for the rapid postnatal up-regulation of fatty acid oxidation and ketogenesis critical for cerebral energy metabolism of suckling infants.74,80
Acquisition of spermatozoa motility and maturation needs carnitine and acetylcarnitine. These processes occur in the epididymal tract, where the concentration of carnitine rises up to 63 mM, which is more than thousand-folds the average plasma value of 50 µM ( Fig. 5 ).43,125 To sustain this high-concentration gradient, plasma carnitine transporters are necessary. Indeed, a link between spermatozoa motility and acetylcarnitine uptake mediated by OCTN2 has been reported.62,134
Development of Assay Approaches on OCTNs: Proteoliposome as a Screening Tool
Evaluation of toxic and/or beneficial effects of xenobiotics and drugs using animal models implies very high costs and ethical problems. Thus, in vitro and in silico models for screening adverse effects and/or potential benefits of drugs are very welcome if these strategies could give suitable information on toxicity avoiding, at least partially, animal experimentation. This might be helpful both in reducing costs and in accelerating the process of drug development from in vitro studies to application of human therapy.
Functional studies on OCTN proteins have been performed since their identification, using cell systems. These consist of two principal types of experimental models: (1) intact cells extracted from animal tissues and (2) modified cells, such as Xenopus laevis oocytes or mammalian cell lines in which the homologous or heterologous transporters can be expressed by injecting cRNAs or by transient or stable transfection with recombinant plasmids. In the first model, the specific OCTN transporters have been functionally identified using specific substrates and inhibitors that allow discrimination of their activities from those of other transporters.32,33,78,81,91 In the second model, the protein of interest is overexpressed, improving the discrimination with respect to other similar endogenous transporters.24 -28,33,37,41,60 –62,64 –67,97,98 This procedure, in fact, allows an increase in the transport signal specifically due to the expressed OCTN transporter with respect to the background measured in control cells (i.e., cells not injected or transfected with empty vectors). The transport activity is monitored, in both systems, using radioactive labeled substrates or drugs added to cells at time zero. At defined times of transport, the external substrates that have not entered the cells are removed using filtration procedures, and the internalized radioactivity is counted. These systems allow measurement of the rate of uptake and, hence, the derivation of transport kinetics related to the extracellular side of the transporter. Effects of interacting molecules such as drugs can also be studied. Parameters such as the IC50 and the Ki can be calculated by inhibition kinetics, whereas it is difficult to perform measurements of efflux rates for defining properties of the intracellular side of the transporters. Additional experimental problems arise using the cell models because of the complexity of the systems. Interferences may affect the transport measurements, caused by enzyme pathways that could chemically modify the taken-up substrates or by the presence of similar transporters in the same cells, which can contribute to substrate uptake, leading to overestimation of the transport signal. Proteoliposomes constitute a good alternative to intact cells.15–17,22,29,40,41,135–137 Proteoliposomes are obtained by freeze-thaw sonication procedures, which have been successfully used for reconstituting the recombinant human glucose-Na+ symporter hSLGT1, rat organic cation and anion transporters rOCTs and rOATs, and human organic cation transporter hMATE2.138 -140 In this procedure, transport proteins are inserted in the membrane bilayer during fast freezing at a very low temperature in liquid nitrogen and with slow thawing. A different procedure of proteoliposome formation was pointed out some decades ago and used for studying mitochondrial transporters. 141 This method has since been modified and optimized for studying the OCTN plasma membrane transporters.15–17,22,29,40,41,137 It is based on the slow removal of detergent from micelles of phospholipids, protein, and detergent and leads to formation of unilamellar phospholipid vesicles of about 100 nm average diameter. This procedure allows the transporter insertion in the artificial membrane with the same orientation as in the cell membrane. This is caused by the asymmetrical structure of the proteins (see Fig. 1 ) and by the small radius of the starting micelles. The transport activity is then assayed by measuring the uptake (or efflux) of radioactive substrates such as 3H-carnitine, 3H-TEA, 3H-acetylcholine, or other compounds, externally (or internally) added to proteoliposomes. Using this protocol, several types of experiments can be performed, by changing or adding different components or modifying some steps of the procedure (see the sketch in Fig. 3 ). The efflux can be detected as well upon including the radioactive substrate inside the vesicle, by transporter-mediated equilibration.29,40 The main advantages of this model with respect to the cell systems are the reliable control of the experimental conditions in the internal compartment and, hence, the possibility of determining the internal Km for different substrates or the action of internal effectors on the transport ( Fig. 3 ); the elimination of interferences caused by enzyme pathways, which could modify the taken-up substrates; the absence of other transporters in the same vesicles guaranteed by the higher phospholipids/protein ratio (one transport protein molecule per liposome) with respect to the cell membrane (several transport protein molecules per cell); and the presence of a single protein species in the case of purified protein preparation.15–17,29,40,43,135–137 Moreover, the lipid composition of the proteoliposomal membrane can be varied to obtain information on the influence of specific lipids on the activity of a transport system. This is an important feature because, after lipidomic development, the role of lipids in regulating membrane protein function is becoming emergent. 142 As examples, using proteoliposomes, CACT and ornitine/citrulline mitochondrial transporters were found to be regulated by specific phospholipids. 143 Some disadvantages of the proteoliposome system compared with entire cells may be the lack of possible physiological protein-protein interactions or altered insertion in the membrane phospholipids that in cells is assisted during the trafficking process. These discrepancies can be excluded by comparing some basic functional properties analyzed both in cells and in proteoliposomes; their coincidence is a useful indication of correct assembly of the transporters in proteoliposomes.15–17,135
Interestingly, and in line with the new needs of drug experimentation, the proteoliposome model allows screening of interactions of xenobiotics and, in general, chemicals with any specific reconstituted transporter. Indeed, this methodology is highly reproducible, much faster than the use of cell systems, and generally less expensive. Furthermore, because in the proteoliposome experimental system it is possible to reconstitute precise amounts of transporters, standardized specific activities can be assayed. Thus, each interesting molecule can be studied not only in terms of affinity but also of mechanism of interaction. In proteoliposomes, but also in X. laevis, site-directed mutants of recombinant proteins can be studied to identify amino acid residues responsible for the interaction/transport.43,115 Using the described experimental models, functional properties of many rat and human transporters have been revealed. In particular, hOCTN1 and mOCTN3 overexpressed in E.coli and rOCTN2 extracted from rat kidney have been extensively studied in proteoliposomes, revealing structure/function relationships and mechanism of interaction with drugs and toxic compounds.14–17,22,137 Experiments using the proteoliposome model ( Fig. 3 ) led to description of novel properties of the OCTN1 transporter, such as the ability to transport acetylcholine, the dependence of this transport function by internal ATP, and the inhibition by acetylcarnitine and choline, which are potential substrates. A screen of some amino acid–specific chemical reagents has been performed. The data demonstrate interaction of most of these compounds with thiol or amino groups of the transport protein.16,29 The proteoliposome model will contribute to solving the existing discrepancies on the OCTN1 function, such as the electrical nature of transport. With regard to this aspect, it was formerly found that potassium diffusion potential, generated in proteoliposomes by the ionophore valinomycin (see the sketch in Fig. 3 ), did not influence the transport of both TEA and acetylcholine, indicating an electroneutral mechanism (Pochini et al., unpublished results).
Interactions of OCTNs with Drugs and Xenobiotics Studied in Cell Systems
Studies conducted for many years in cell systems highlighted that several types of pharmacologic compounds interact with OCTN subfamily members, validating the definition of OCTNs as polyspecific transporters. 4 In particular, hOCTN2 was described to mediate transport of the antipsychotic sulpiride, pirylamine, quinidine, verapamil, and valproate.64,111,144 These drugs are also reported to induce carnitine deficiency; as an example, a study on patients with Huntington’s disease showing hypocarnitemia and treated with valproate has been conducted. 58 Several β-lactams were found to inhibit and compete with the transport of carnitine catalyzed by OCTN2; among them, cephaloridine and cefepime were the most effective (IC50 0.23 mM and 1.7 mM, respectively). 75 More recently, it was described that OCTN2, from rat, human, and chicken, is involved mainly in the transport of mildronate. 94 Drugs used in treatment of neurologic disorders, such as serotonin, dopamine, norepinephrine, thiamine, diphenhydramine, procainamide, and lidocaine, inhibit carnitine transport mediated by mOCTN2. 65 Mouse and human OCTN2 isoforms were also described as cephaloridine and imatinib transporters, respectively.95,145 Zidovudine, levofloxacin, and grepafloxacin were found to inhibit hOCTN2 with a noncompetitive mechanism.146,147 In some cases, the type of inhibition was defined, but little or no information is available concerning the molecular mechanisms of interaction. The IC50 values for some drugs were in the micromolar range, thus being compatible with an in vivo effect, whereas the IC50s found for antibiotics were higher than the concentrations of these compounds in the blood, after administration. 75 Quantitative structure/activity relationships for the interaction of OCTN2 with drugs have been documented in cells. 76 Twenty chemical structure fragments were identified as possible determinants of OCTN2 inhibition. For some structures containing such fragments, the inhibition mechanism was also revealed in proteoliposomes. 22
OCTN2, expressed also in the heart (see the subsection “OCTN2”), is inhibited by many cardiovascular drugs commonly used in human therapy, such as verapamil, spironolactone, and mildronate, for which a competitive mode of inhibition was observed. 92 OCTN2 and OCTN1 have been described to be responsible for oxaliplatin transport in embryonic kidney 293 cells and rat dorsal root ganglion neurons.11,59 hOCTN2 is inhibited by other antitumor agents such as etoposide, vincristine, and vinorelbine. 148
Interestingly, it should be highlighted that although OCTN2 seems to be quite specific for physiological substrates (see the subsection “OCTN2” and Fig. 2 ), this is not the case regarding the interaction with drugs.22,149 This feature makes it very interesting for biomolecular screening and for delivery of acetylcarnitine that can be used as a precursor of acetylcholine biosynthesis ( Fig. 4 ), which is connected with glucose homeostasis and IBDs (see the section “Involvement of OCTNs in Human Pathology”).
Less information concerning the interaction of OCTN1 with drugs is available. So far, interactions with gabapentin, ipatropium, pyrilamine, quinidine, and verapamil have been described, even though no information on the mechanism of inhibition has been obtained.149,150 On the basis of studies in cell systems, it was proposed that OCTN1 mediates the cellular uptake of mitoxantrone and doxorubicin, thereby conferring cellular sensitivity to anticancer agents. 30
Nonetheless, the studies described give very little information on the mechanism of interaction of drugs with OCTNs. This is probably due to previously cited limits of the cell experimental model, such as the interferences affecting transport measurements. This gap may be filled up by using experimental systems that allow fine control of experimental conditions.
Interactions of OCTNs with Drugs and Xenobiotics Studied in Proteoliposomes
The proteoliposome experimental model overcomes some of the limits of the cell systems, thus allowing the study of mechanisms of interaction.
Investigations of the effect of xenobiotics on the OCTN2 extracted from rat kidney brush borders have already been approached in proteoliposomes. The interactions of carnitine nitro-derivatives, omeprazole, and mercury toxic compounds have been recently investigated.22,137,151
Because carnitine is a physiological compound that is taken up by cells from diet, this molecule can be used as a vehicle for specific small molecules provided that a linkage could be achieved between carnitine and each of these molecules. This strategy has been used to prepare nitro-derivatives of carnitine, which could be used in ischemia. The synthesized molecules have been assayed for transport by OCTN2 with positive results. This is an example of proteoliposome use for prediction of increased bioavailability of newly synthesized molecules. 151
Moreover, the proteoliposome tool has been used for a very accurate analysis of the interaction of the widely used proton-pump inhibitor omeprazole with rOCTN2. This has highlighted, for the first time, the existence of two different mechanisms of interaction with the transporter. One mechanism consists of the formation of a mixed disulphide between the drug and the protein, according to the structural properties of omeprazole. This interaction occurs between an active group of the drug and a Cys residue of the protein. A second mechanism involves noncovalent binding of the drug to a protein site that is close to the substrate binding site, leading to competitive inhibition. Both types of interaction of omeprazole with rOCTN2 lead to complete inactivation of the transporter with different IC50. In the case of covalent interaction, the IC50 is 5.7 µM, lower than that measured for the noncovalent interaction, which is 20.4 µM. The lower value is in the range of the omeprazole concentration in blood after oral administration; it is thus plausible that in vivo omeprazole will interact with OCTN2, causing impairment of carnitine transport. Interestingly, the side effects of omeprazole overlap the symptomatology of the PCD, although in a milder form. 22 This represents strong evidence that in the proteoliposome system, it would be possible to predict transporter-drug interactions and, hence, to predict possible off-targets and potential side effects.
The same tool has been used to gain further insight in the mechanism of toxicity exerted by a group of environmental pollutants, that is, mercuric compounds (CH3Hg and HgCl2), on rOCTN2. One of the most common mechanisms of interaction of mercurials with proteins is the reaction with thiol groups of Cys residues leading to a formation of an Hg-S covalent bond. As in the case of omeprazole, OCTN2 can interact with HgCl2 and CH3Hg via Cys thiol residues exposed in the active site of the protein, leading to a strong inhibition of carnitine uptake mediated by the reconstituted transporter. The calculated IC50 values were 2.5 µM for HgCl2 and 7.4 µM for CH3Hg, both in the range of mercury concentration in the tissue upon exposure to pollutants. In particular, kinetic analysis showed a noncompetitive inhibition mechanism for HgCl2, with a Ki of 4.2 µM, and a mixed inhibition mechanism for CH3Hg, with a Ki of 13 µM. This led to the conclusion that the active and the inhibitory sites overlap; in fact, the addition of the substrate carnitine before the inhibitor molecules exerted complete protection of the inhibition. This would suggest the use of carnitine to partially prevent the toxicity of mercury compounds. 137 Even though the experiments have been performed using rOCTN2, it should be highlighted that hOCTN2 and rOCTN2 share 85% identity; thus, the results obtained can be extended to humans. Moreover, recombinant human transporters could be used for studies in proteoliposomes. On the basis of the description above and other studies conducted on amino acid transporters, the proteoliposome system can be validated as a suitable model for large-scale screening of xenobiotics and drug effects on membrane transport proteins. These procedures will provide new efforts in predicting potential bioavailability and side effects, thus providing important information on toxicity and allowing the bypass of some steps of the animal experimentation for developing new drugs.137,151
Perspectives
The interest in the study of transport across the cell membrane increased exponentially in the past decade. This was essentially due to the relevance of this research field, not only in human physiology but also in human pathology and drug interactions. In mammalian cells, the transport of nutrients and metabolites is carried out by a huge number of proteins, which differ in structure and function. Many transporters have been functionally characterized using eukaryotic cell systems or special cell lines such as HEK293 or X. laevis oocytes in which specific transporters were expressed.24–27,60,103,153 Other transporters that have been predicted on the basis of gene sequences have been studied only to some extent, and their function and specificity for substrates are still lacking or controversial. The state of knowledge on transport systems is clearly and systematically described in a number of reviews.154–161 In the present review, the development of the proteoliposome assay as a tool for studying the function of and for large-scale screening of xenobiotics and drug interactions with membrane transporters such as OCTNs is highlighted, in line with a recent study conducted on the amino acid transporter ASCT2 for screening specifically designed antitumor molecules. 152 The broad specificity of OCTNs and their link with many human pathologies make them very interesting targets of cationic and zwitterionic compounds with potential effects on cell functions and, hence, application to therapy.
Abbreviations
ASD, autism spectrum disorder; CACT, carnitine/acyl-carnitine transporter; CAT, acetylcarnitine transferase; ET, ergothioneine; IBD, inflammatory bowel disease; OAT, organic anion transporter; OCT, organic cation transporter; OCTN, organic cation transporter novel; PCD, primary carnitine deficiency; PI, package insert; PPARα, peroxisome proliferator activated receptor α; TEA, tetraethylamonium.
Footnotes
Acknowledgements
We thank Dr. Barbara Caponsacco for grammar and style reviewing.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Ministry of Instruction University and Research (MIUR)–Italy, by a grant from PON-ricerca e competitività 2007-2013 (PON project 01_00937: “Modelli sperimentali biotecnologici integrati per la produzione ed il monitoraggio di biomolecole di interesse per la salute dell’uomo”).