
Abstract
The relaxin hormone is involved in a variety of biological functions, including female reproduction and parturition, as well as regulation of cardiovascular, renal, pulmonary, and hepatic functions. It regulates extracellular matrix remodeling, cell invasiveness, proliferation, differentiation, and overall tissue homeostasis. The G protein–coupled receptor (GPCR) relaxin family receptor 1 (RXFP1) is a cognate relaxin receptor that mainly signals through cyclic AMP second messenger. Although agonists of the receptor could have a wide range of pharmacologic utility, until now there have been no reported small-molecule agonists for relaxin receptors. Here, we report the development of a quantitative high-throughput platform for an RXFP1 agonist screen based on homogenous cell-based HTRF cyclic AMP (cAMP) assay technology. Two small molecules of similar structure were independently identified from a screen of more than 365 677 compounds. Neither compound showed activity in a counterscreen with HEK293T cells transfected with an unrelated GPCR vasopressin 1b receptor. These small-molecule agonists also demonstrated selectivity against the RXFP2 receptor, providing a basis for future medicinal chemistry optimization of selective relaxin receptor agonists.
The relaxin hormone is involved in a variety of biological functions in normal tissues and diseases. The role of relaxin is well established in female reproduction and parturition, mammary gland and endometrial development, and maintenance of myometrial quiescence during pregnancy. Relaxin signaling through its cognate G protein–coupled receptor (GPCR), the relaxin/insulin-like family peptide receptor 1 (RXFP1), results in extracellular matrix remodeling through regulation of collagen deposition, cell invasiveness, proliferation, and overall tissue homeostasis. Significantly, the therapeutic effects of relaxin in the treatment of renal, cardiac, skin, lung fibrosis, inflammation, and wound healing in animal models are well established. 1 Other data strongly indicate the significance of relaxin in prostate, breast, thyroid, and other tumorigenesis. 2 Several clinical trials have been conducted with recombinant human relaxin hormone as a treatment for scleroderma, cervical ripening, fibromyalgia, preeclampsia, and congestive heart failure. The wide range of effects and applications of relaxin underscores the importance of this hormone in human physiology and diseases. Small-molecule agonists would be extremely useful in furthering the understanding of relaxin hormone biology and have the potential for therapeutic development.
The cognate relaxin receptor, RXFP1, is a member of the relaxin/insulin-like family of GPCR peptide receptors.3,4 RXFP2 is another member of this family (previously called LGR7 and LGR8, respectively), which shares 60% sequence identity and similar structure. RXFP2 is a cognate receptor for insulin-like 3 peptide, belonging to the same group of peptide hormones as relaxin. 5 Relaxin can activate cyclic AMP (cAMP) production in cells transfected with RXFP2 in vitro, however with lower potency. RXFP1 was deorphanized in 2002 and was found to couple to Gαs proteins, which in turn stimulate adenylate cyclase (AC) to generate cAMP. 3 In addition, RXFP1 also couples to Gi/Go proteins to further modulate the cAMP production. The activated G-α derived from Gi/Go inhibits the cAMP production by AC, whereas the G-βγ subunits activate the delayed cAMP response via PI3K and PKCζ. 4 The RXFP1 and RXFP2 receptors contain a unique low-density lipoprotein class A (LDLa) module at the amino terminus, followed by an extracellular leucine-rich repeats (LRRs) and seven transmembrane helices of the transmembrane domain. 1 The LRRs and the exoloops within the transmembrane domain have been shown to participate in ligand binding,6,7 whereas the LDLa modules contribute to receptor activation.8,9
Multiple reports have indicated that expression of RXFP1 in human embryonic kidney cells (HEK293T) is able to signal through the classical Gαs-coupled mechanism to activate AC, resulting in elevation of intracellular and extracellular cAMP concentrations.3,4 We have developed a homogeneous cAMP assay using HEK293T cells stably expressing human RXFP1 receptor to screen for small-molecule agonists of human RXFP1. The assay was successfully miniaturized and used to screen a 365 677-compound library in quantitative high-throughput screening (qHTS) format. Following confirmation and counterscreening, we have identified small-molecule agonists that are >100-fold selective toward RXFP1 over RXFP2, making this the first report of a series of small-molecule agonists for the human RXFP1 receptor.
Materials and Methods
Materials
Ro 20-1724 and forskolin were purchased from Sigma-Aldrich (St. Louis, MO). HTRF cAMP HiRange kit was purchased from CisBio (Bedford, MA), and the cAMP-Screen enzyme-linked immunosorbent assay (ELISA) assay kit from Life Technologies (Carlsbad, CA). Porcine relaxin 10 was a gift from Dr. O. David Sherwood (University of Illinois at Urbana-Champaign).
Cell Line Generation
The HEK293T cells stably transfected with human RXFP1 (HEK293-RXFP1) 11 or RXFP2 (HEK293-RXFP2) 3 expression constructs were kindly provided by Drs. András Kern and Gillian D. Bryant-Greenwood (University of Hawaii) and Dr. Sheau Yu Teddy Hsu (Stanford University). HEK293T cells stably transfected with V1b (HEK293-V1b) was purchased from PerkinElmer (Waltham, MA).
Cell Culture
HEK293-V1b cells were grown in DMEM medium with 10% fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin at 37 °C in 5% CO2. HEK293-RXFP1 and HEK293-RXFP2 cells were grown in the same conditions with the addition of 500 µg/mL geneticin for HEK293-RXFP1 cells and 100 µg/mL zeocin for HEK293-RXFP2 cells. All cell lines were grown at 37 °C in the presence of 5% CO2 and 95% humidity. Cells were passaged once they reach 80% confluence at a split ratio of 1:8.
HTRF cAMP Assay
The initial assay optimization and all follow-up assays were conducted in 384-well format. For 384-well format assays, cells were seeded as 8000 cells/well in 30 µL/well media with a MultiDrop Combi dispenser (Thermo Scientific, Waltham, MA) and allowed to attach overnight at 37 °C in 5% CO2. Following the incubation, 2 µL/well of 1.6 mM Ro 20-1724 in PBS+ (Dulbecco’s phosphate-buffered saline [DPBS], 1 mM CaCl2, 0.5 mM MgCl2, 0.05% bovine serum albumin [BSA], 0.005% Tween 20) was dispensed using a BioRAPTR FRD dispenser (Beckman Coulter, Brea, CA). Subsequently, 0.25 µL/well of compound solutions in DMSO was dispensed with CyBi-well dispenser (CyBio, Jena, Germany). The cells were stimulated with compounds for 30 min at 37 °C in 5% CO2, after which 8 µL/well of each HTRF cAMP HiRange kit (CisBio, Bedford, MA) detection reagent was dispensed with a BioRAPTR FRD dispenser. The detection reagents were diluted as such: K-anti-cAMP antibody at 1:20 and cAMP-d2 at 1:18 in HTRF lysis buffer (supplied by the assay kit). The plates were incubated for 30 min at room temperature before the signal was read on an Envision plate reader (PerkinElmer, Waltham, MA).
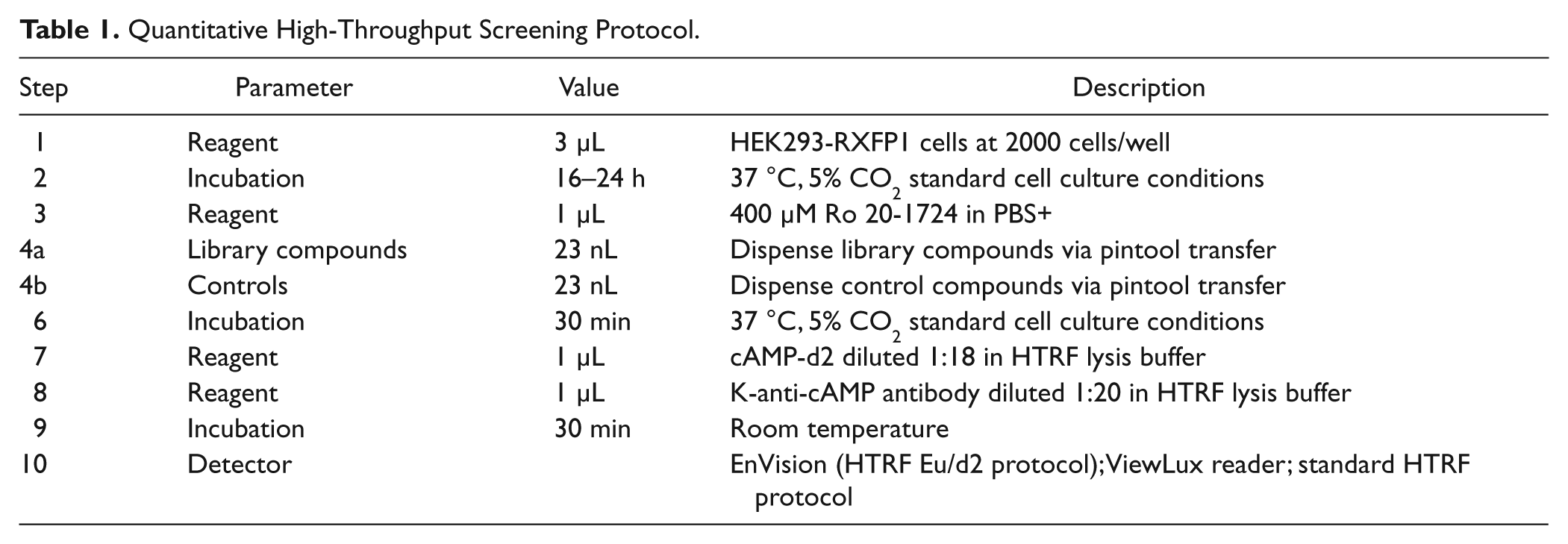
The primary screen was done in a 1536-well format ( Table 1 ). Briefly, HEK293-RXFP1 cells were seeded at 2000 cells/well in 3 µL/well growth media with a MultiDrop Combi dispenser (Thermo Scientific, Waltham, MA) and allowed to attach overnight at 37 °C in 5% CO2. Following the incubation, 1 µL/well 400 µM Ro 20-1724 in PBS+ (DPBS, 1 mM CaCl2, 0.5 mM MgCl2, 0.05% BSA, 0.005% Tween 20) was dispensed using a BioRAPTR FRD dispenser (Beckman Coulter). Then, 23 nL/well of compound solutions in DMSO was dispensed via pintool transfer (Kalypsys, San Diego, CA). The cells were stimulated with compounds for 30 min at 37 °C in 5% CO2, after which 1 µL/well of each HTRF detection reagent was dispensed with a BioRAPTR FRD dispenser. The detection reagents were diluted as such: K-anti-cAMP antibody at 1:20 and cAMP-d2 at 1:18 in HTRF lysis buffer (supplied by the assay kit). The plates were incubated for 30 min at room temperature, and then the signal was read on a ViewLux plate reader (PerkinElmer).
Quantitative High-Throughput Screening Protocol.
ELISA cAMP Assay
The ELISA cAMP assay was performed in a 96-well format. Cells were seeded at 16 000 cells/well in 95 µL/well media and allowed to attach overnight at 37 °C in 5% CO2. Following the incubation, 30 µL/well of 417 µM Ro 20-1724 in PBS+ was added, and immediately following that, 1.6 µL/well of compound solutions in DMSO was added with a multichannel pipette. The cells were stimulated with compounds for 30 min at 37 °C in 5% CO2, after which the cells were lysed and cAMP levels in the lysates were assayed according to the cAMP-Screen ELISA assay kit (Life Technologies, Carlsbad, CA). The luminescence signal was detected with a ViewLux plate reader (PerkinElmer).
Compound Library Preparation and qHTS
The library of pharmacologically active compounds (LOPAC1280) consists of a collection of small molecules with characterized biological activities. The LOPAC1280 library has been extensively used for HTS assay validations12,13 and was purchased from Sigma-Aldrich. The Molecular Libraries-Small Molecule Repository (ML-SMR) includes a library of structurally diverse compounds (http://mli.nih.gov/mli/compound-repository/mlsmr-compounds/), which consisted of 365 677 compounds at the time of screening. Compounds from both libraries were serially diluted 1:5 in DMSO in 384-well plates to yield four concentrations (from 0.64 µM to 10 mM) and formatted into 1536-well plates at 7 µL/well. 14 A qHTS was performed using a fully automated robotic screening system (Wako, San Diego, CA), as described previously. 15 Final compound concentrations during cell incubation ranged from 0.46 µM to 57.5 µM.
Data Analysis
Primary screen data were analyzed with customized software developed internally. The maximal response (100% activity) was determined by the response of 28.75 nM relaxin, and the basal signal (0% activity) was measured by the 0.58% DMSO control in the HEK293-RXFP1 cells. All follow-up assays were conducted with three cell lines (HEK293-RXFP1, HEK293-RXFP2, and HEK293-V1b). Because relaxin is less potent in stimulating cAMP production in the HEK293-RXFP2 cell and does not stimulate HEK293-V1b cells, 57.5 µM forskolin treatment was used as the maximal response for data normalization. The concentration responses of all the compounds were analyzed using methods described by Inglese et al, 15 and structural clustering of active compounds was performed using Leadscope Hosted Client (Leadscope Inc., Columbus, OH). The EC50 values of compounds in the confirmation and follow-up experiments were calculated from the dose-response curves by nonlinear regression analysis using Prism software (GraphPad Software, San Diego, CA).
Results
Assay Miniaturization and Validation
The RXFP1 receptor signals through cAMP second messenger. To screen for agonists of the receptor, we used human embryonic kidney cells stably transfected with human RXFP1 receptor (HEK293-RXFP1), which have previously been shown to constitute a functional cell-based model for the RXFP1-cAMP signaling pathway.3,4,16 To amplify the cAMP signal, the assay was conducted in the presence of a PDE4 inhibitor, Ro 20-1724. 17 cAMP levels were detected using the HTRF cAMP assay kit. 18 The detection kit consists of a europium cryptate labeled anti-cAMP antibody (K-anti-cAMP) and d2 dye labeled cAMP (cAMP-d2) as tracers in a time-resolved fluorescence energy transfer (TR-FRET) detection system. The TR-FRET between the K-anti-cAMP and cAMP-d2 is disrupted by cAMP in cell lysates, thus allowing detection of the second messenger in a homogenous format that is suitable for qHTS. The HTRF cAMP assay was initially developed in 384-well format and then miniaturized to 1536-well format with an assay volume of 4 µL. The functionality of the HEK293-RXFP1 cells was tested by examining the cAMP response to porcine relaxin, which has previously been shown to activate RXFP1 with similar potency of H2 human relaxin, 10 and forskolin, an adenylate cyclase activator. The concentration-response curves of relaxin and forskolin in 384-well and 1536-well formats showed good correlation, with EC50 values of 203 pM and 494 pM for relaxin and 164 nM and 193 nM for forskolin in 384-well and 1536-well formats, respectively ( Fig. 1a , b ). The tight correlation between the two plate formats, along with the fact that the potency of relaxin in the HTRF assay also matches that of previously published results in HEK293-RXFP1 cells,7,16,19 indicates that this assay adequately detects RXFP1 activation in the miniaturized format. The signal for maximal stimulation was found to be similar between relaxin and forskolin in the HTRF cAMP assay, with mean ratios from 12 treatment wells in 384-well plate format being 0.78 for relaxin and 0.54 for forskolin treatments, compared with 4.9 for DMSO treatment ( Fig. 1c ). Although previous reports have noted that maximal relaxin stimulation is lower than that of maximal forskolin stimulation, 4 differences in RXFP1 expression levels and/or inclusion of the PDE4 inhibitor Ro 20-1724 might have resulted in the more robust cAMP response.
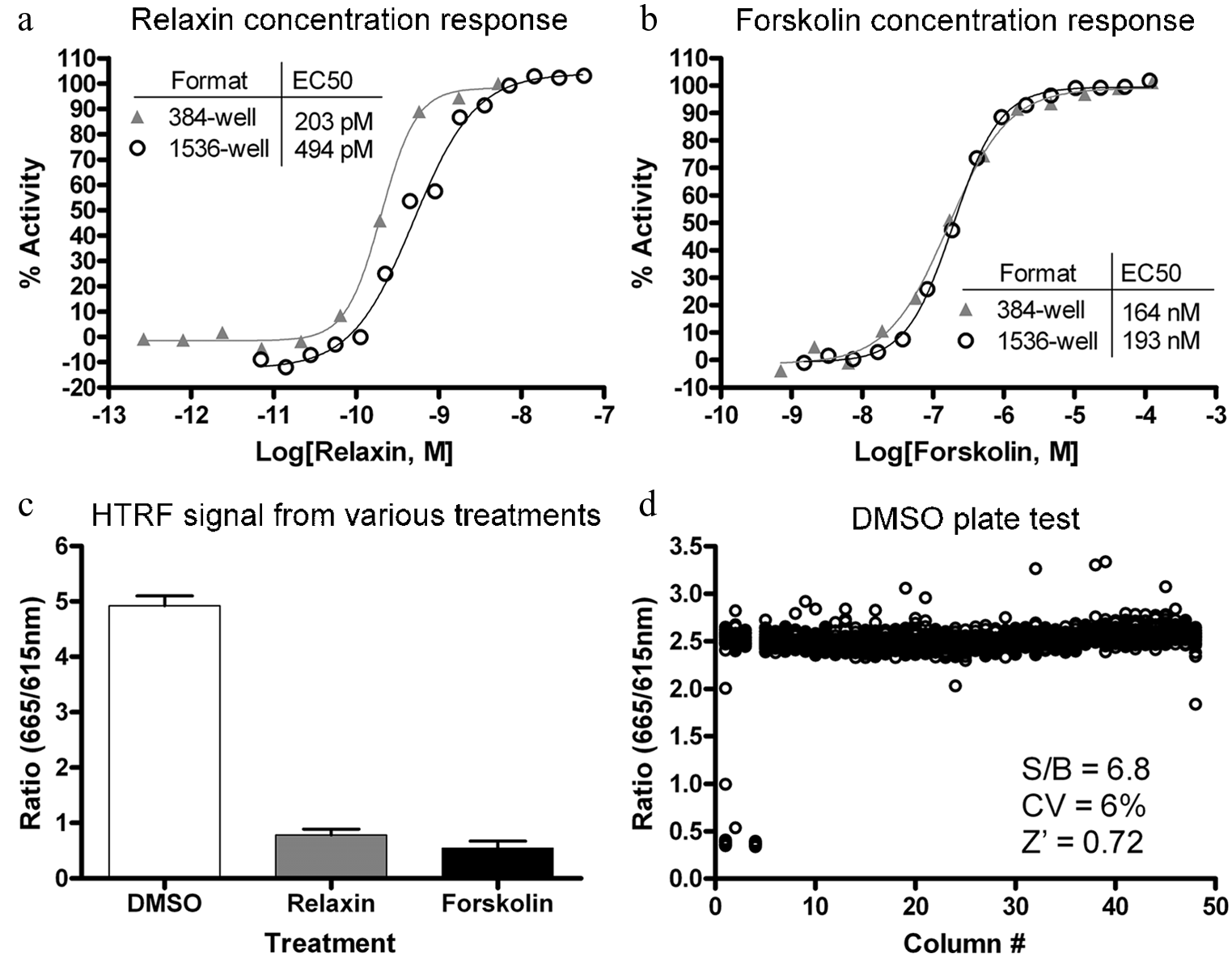
Assay miniaturization and validation. Concentration responses for (
Assay parameters were then assessed in miniaturized plate format against a DMSO control plate, as a solvent control for the compound library. The signal-to-basal ratio (S/B), the coefficient of variation (CV), and Z factor values were 6.8, 6%, and 0.72, respectively ( Fig. 1d ). As a further validation, the assay was used to screen against the LOPAC1280 library of bioactive compounds. The LOPAC1280 library was screened in qHTS format at five concentrations ranging from 92 nM to 57.5 µM. The assay showed robust and clean performance with average S/B, CV, and Z factor values of 6.8, 8.3%, and 0.78, respectively. This preliminary screen identified 13 compounds that showed EC50 values less than 10 µM with efficacy greater than 50%. Upon closer examination, these compounds included the positive control compound, forskolin, as well as various agonists of Gs-coupled GPCRs, such as adenosine and adrenergic receptors, which are known to be expressed in HEK293T cells. The fact that the validation screen was able to isolate known GPCR agonists and forskolin indicated that the assay was effective at identifying upregulators of cAMP. Expectantly, when these 13 hits were counterscreened in an HEK293T cell line stably expressing the vasopressin 1b receptor (HEK293-V1b), they were found to be active at similar potencies and efficacies. Therefore, although the LOPAC1280 screen failed to find true agonists of the human RXFP1 receptor, the results validated both the primary assay and counterscreening strategy.
Quantitative High-Throughput Screen
Upon failing to isolate human RXFP1 agonists from the LOPAC1280 library, the HTRF cAMP assay was used to screen the entire ML-SMR collection of 365 677 compounds using a fully automated screening system in qHTS format. 15 All compounds were assayed at four concentrations of 57.5 µM, 11.5 µM, 2.3 µM, and 92 nM. The response to control agonists (relaxin and forskolin) was measured in each plate and was stable across the entire screen.
During screening, freshly prepared cells were dispensed into 1536-well assay plates as 3 µL/well at a rate of 100 plates per hour and incubated at 37 °C with 5% CO2 for 16 to 24 h prior to the robotic screening. The screen was initiated by addition of 1 µL/well PDE inhibitor Ro 20-1724, 17 followed by 23 nL/well compound or control addition via pin transfer. Subsequently, the cells were incubated for 30 min at 37 °C before 1 µL/well of each HTRF detection reagent was dispensed, and the signal was read on a ViewLux plate reader. The assay showed robust performance with an average S/B of 3.1 and Z′ factor of 0.66 over 1140 plates screened. The concentration response of compounds and the structure-activity relationship were analyzed based on primary screen data. A total of 434 compounds were cherry-picked for follow-up studies, for an initial hit rate of 0.1%. Compounds were considered hits if they showed high potency (<10 µM EC50) and efficacy (>60% of forskolin activation) irrespective of donor channel effects or low potency (>50% activation at 57.5 µM) without donor channel effects.
Follow-up Studies
The cherry-picked compounds were tested in 12-point intraplate 1:3 titration in 384-well plate format to generate more detailed concentration-response curves. Hit confirmation was carried out with HEK293-RXFP1 cells, counterscreening was carried out with HEK293-V1b cells, and hit selectivity was tested with HEK293T cells stably expressing human relaxin family receptor 2 (HEK293-RXFP2; Fig. 2 ). Although forskolin was able to stimulate cAMP production in all three cell lines with similar potencies (EC50 of 56.2 nM–167 nM), relaxin showed >30-fold selectivity toward the RXFP1 over the RXFP2 receptor and showed no activity toward the V1b receptor ( Fig. 3a , b ). The activity of relaxin against RXFP1 and RXFP2 agrees with previously published values.3,4,16
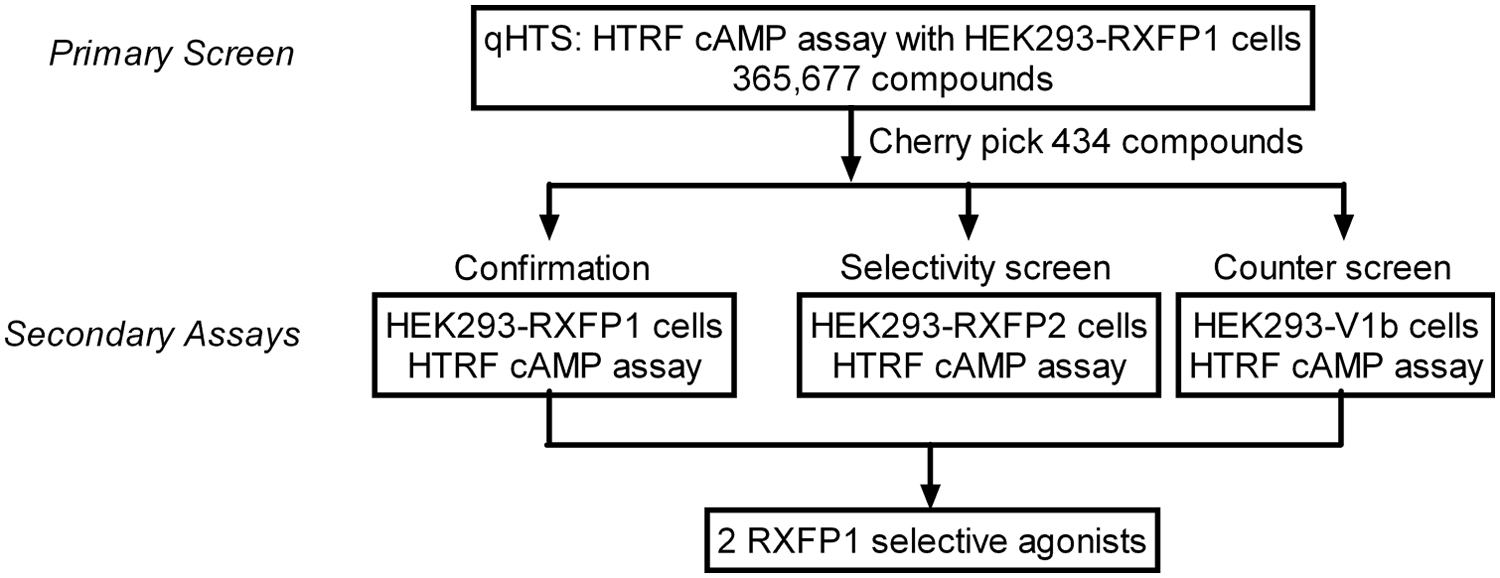
Summary of quantitative high-throughput screening and follow-up assays.
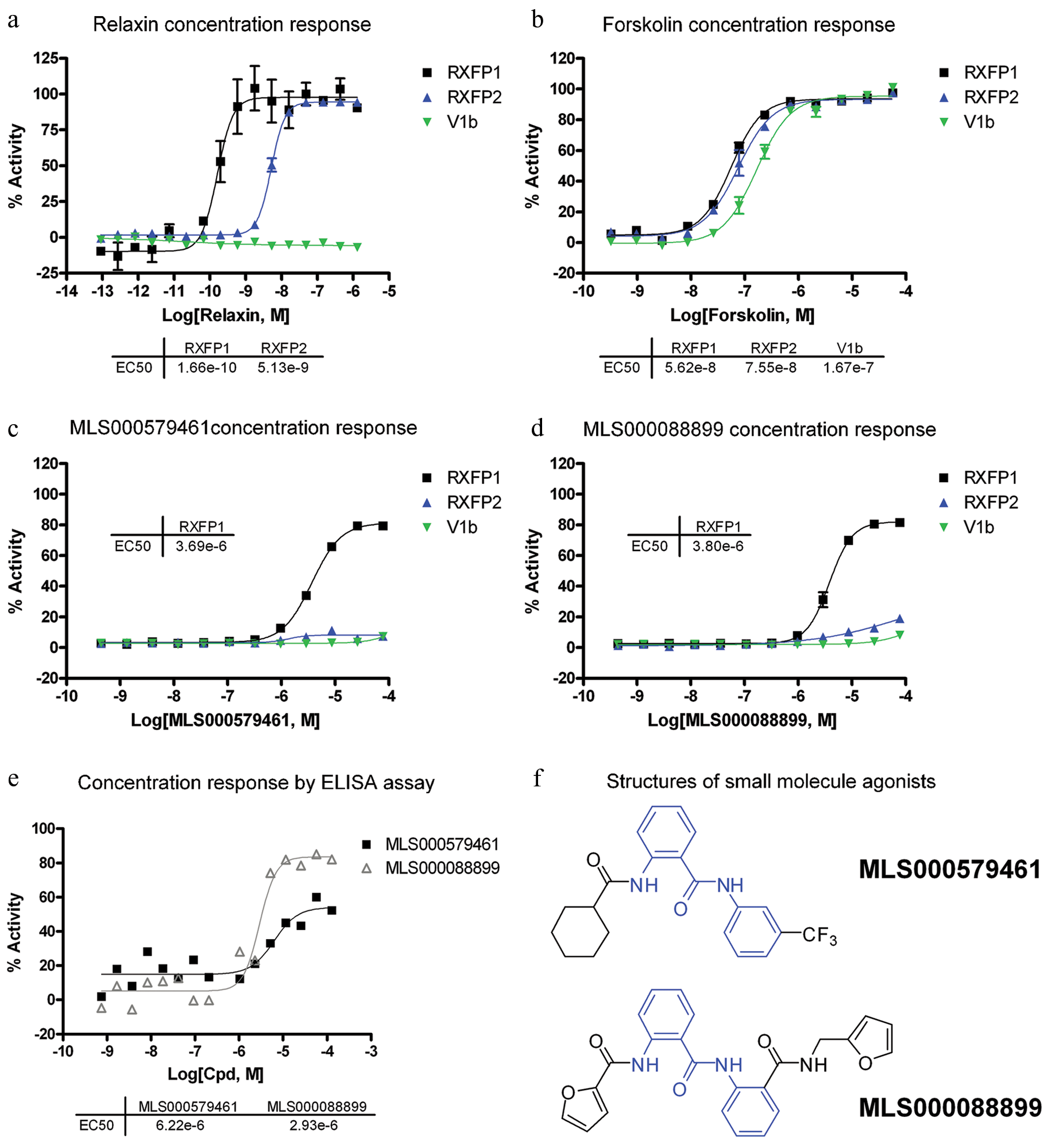
Concentration response of human RXFP1 agonists. HTRF cyclic AMP (cAMP) response in HEK293-RXFP1 (black), HEK293-RXFP2 (blue), and HEK293-V1b (green) cell lines and percentage viability (gray) for (
Of the 434 hits, only two compounds, MLS000579461 and MLS000088899, were found to be selective toward the RXFP1 receptor ( Fig. 3c , d ). The two compounds show similar potencies (3.7 µM and 3.8 µM) and efficacies (80%). Furthermore, both compounds belong to the same chemical scaffold as they contain a 2-acetamido-N-phenylbenzamide core motif ( Fig. 3f , in blue).
The activities of the two hits were further confirmed with an ELISA assay for cAMP detection as an orthogonal assay that is not subjected to the same detection format. MLS000579461 and MLS000088899 showed EC50 values of 6.2 µM and 2.9 µM and efficacies of 60% and 80%, respectively ( Fig. 3e ). These values correlate well with those obtained from the HTRF cAMP assays.
Discussion
We report here the first series of small-molecule agonists of the relaxin receptor RXFP1. The miniaturized HTRF cAMP assay performed well in both assay validation and qHTS. The validation screen against the LOPAC1280 library of bioactive compounds did not find any RXFP1 agonists. Based on this, a low hit rate was expected from qHTS. Thus, a relatively lenient cherry-picking criterion was implemented. Compounds were considered a hit if they showed high potency (<10 µM EC50) and efficacy (>60% maximal activation) irrespective of donor channel effects or had lower potency (>50% activation at 57.5 µM) without donor channel effects. Because the HTRF cAMP assay uses a ratiometric measurement, compounds that affect the donor channel fluorescence can appear as false-positives. However, potent compounds affecting donor channel fluorescence were cherry-picked because the false-positives could be screened out with the counterscreen assay.
Upon filtering the 434 hits through the confirmation, counterscreen, and selectivity assays, only two compounds showed selective cAMP activation in HEK293-RXFP1 cells. It is perhaps not surprising that the screen yielded so few true hits because the natural physiological agonist is a peptide hormone that is 6 kDa in size. Full receptor binding and activation require agonist binding at two distinct sites on the receptor, with the A chain of the hormone binding to transmembrane exoloops of the receptor and the B chain binding to the several sites of extracellular LRR domain.7,16 Furthermore, the amino-terminal LDLa domain has also been shown to be essential to receptor activation, whereas the mechanism is still unclear.8,9 Therefore, it would be difficult for a small molecule to fully mimic the large size of the natural agonist and multibinding site mode of activation. Nevertheless, we have identified the first novel series of small-molecule RXFP1 agonists with >100-fold selectivity over RXFP2 receptor through screening of a 365 677-compound library in qHTS format. The fact that the two lead compounds share a common molecular structure indicates that this scaffold is an ideal starting point for initiation of medicinal chemistry to improve potency toward RXFP1.
Footnotes
Acknowledgements
The authors thank Drs. András Kern and Gillian D. Bryant-Greenwood (University of Hawaii) and Dr. Sheau Yu Teddy Hsu (Stanford University) for HEK293-RXFP1 and HEK293-RXFP2 cell lines and Dr. O. David Sherwood (University of Illinois at Urbana-Champaign) for porcine relaxin.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Molecular Libraries Initiative of the NIH Roadmap for Medical Research U54MH084681 and R03 MH085705 (to A.I.A.) and the Intramural Research Program of the National Human Genome Research Institute and National Center for Advancing Translational Sciences, National Institutes of Health.