
Abstract
Wnt/β-catenin signaling has emerged as a central player in pathways implicated in the pathophysiology and treatment of neuropsychiatric disorders. To identify potential novel therapeutics for these disorders, high-throughput screening (HTS) assays reporting on Wnt/β-catenin signaling in disease-relevant contexts are needed. The use of human patient–derived induced pluripotent stem cell (iPSC) models provides ideal disease-relevant context if these stem cell cultures can be adapted for HTS-compatible formats. Here, we describe a sensitive, HTS-compatible Wnt/β-catenin signaling reporter system generated in homogeneous, expandable neural progenitor cells (NPCs) derived from human iPSCs. We validated this system by demonstrating dose-responsive stimulation by several known Wnt/β-catenin signaling pathway modulators, including Wnt3a, a glycogen synthase kinase-3 (GSK3) inhibitor, and the bipolar disorder therapeutic lithium. These responses were robust and reproducible over time across many repeated assays. We then conducted a screen of ~1500 compounds from a library of Food and Drug Administration–approved drugs and known bioactives and confirmed the HTS hits, revealing multiple chemical and biological classes of novel small-molecule probes of Wnt/β-catenin signaling. Generating these type of pathway-selective, cell-based phenotypic assays in human iPSC-derived neural cells will advance the field of human experimental neurobiology toward the goal of identifying and validating targets for neuropsychiatric disorders.
Keywords
Introduction
One of the major obstacles to the identification of therapeutic interventions for central nervous system (CNS) disorders has been the difficulty of studying the step-by-step development of pathophysiology in systems that are amenable to drug and functional genomic screening. 1 Although significant progress has been made toward this goal using immortalized and/or postmortem fetal stem cell–derived neuronal cells,2–4 these studies lack the coverage of a range of patient genotypes, are unable to be coupled to studies on living patients, and lack the ability, in the case of fetal-derived stem cells, to determine disease status later in life. Fortunately, recent advances in the field of stem cell biology and capabilities to create patient-specific human neurons with defined genomes using reprogramming technology provide unprecedented opportunities for both the investigation of the pathogenetic mechanisms of CNS disorders and the discovery of novel therapeutic targets.5,6 Multiple examples now exist of the creation of human-induced pluripotent stem cell (iPSC) models of monogenic CNS disorders (e.g., Fragile X syndrome, Rett syndrome).7,8 There are also growing efforts to address the more challenging case of complex polygenic disorders, including bipolar disorder and schizophrenia.9,10 Although still preliminary, these human iPSC disease models are beginning to provide fundamentally novel insights into the pathogenesis of CNS disorders as well as to allow the functional characterization of disease genes and cellular phenotypes in the context of human neurobiology.
Given these advances, one of the opportunities, and challenges, that exists in the field of human stem cell biology is the creation of a robust and scalable platform for human experimental neurobiology that can support both (1) screening for novel therapeutics using physiologically relevant, defined neural cell subtypes and (2) the identification and validation of novel targets and mechanisms for therapeutic intervention. Given the inherent challenges of direct neural differentiation of iPSCs, including heterogeneity of resulting neural cell types, our strategy to address these needs is to develop robust methods for the routine derivation of expandable homogeneous populations of neural progenitor cells (NPCs) from iPSCs. These NPCs can be maintained as self-renewing, genomically stable cells that can be expanded greatly and subsequently differentiated in vitro to give rise to postmitotic, functional neurons and glial cells on the scale of the millions-billions of cells needed for a large-scale, high-throughput screen (HTS).
Here we describe our initial efforts using this strategy of deriving NPCs from human iPSCs to develop high-throughput, cell-based assays of signaling pathways implicated in a variety of neuropsychiatric diseases with an initial focus on targeting the molecular mechanisms regulating neurogenesis that involve Wnt/β-catenin signaling, a pathway implicated in the response to drugs used to treat bipolar disorder, such as the mood stabilizer lithium, as well as a pathway that has been implicated by genetic factors associated with susceptibility to neuropsychiatric disease.11–14
Materials and Methods
Derivation of Human iPSC-NPCs
iPSCs were reprogrammed from the clinically unaffected human fibroblast cell line, GM08330 (Coriell Institute for Medical Research, Camden, NJ), and characterized as previously described. 7 iPSC clones were maintained on an irradiated mouse embryonic fibroblast (iMEF; GlobalStem, Rockville, MD) feeder layer with daily feeding of iPSC media: 20% Knock-out Serum Replacement (KOSR; Life Technologies, Carlsbad, CA), 1× penicillin/streptomycin (Life Technologies), 1× nonessential amino acids (Life Technologies), additional 1 mM L-glutamine (Life Technologies), 100 µM 2-mercaptoethanol (Bio-Rad, Hercules, CA), 77.5% Dulbecco’s modified Eagle’s medium (DMEM)/F-12 (Life Technologies), and 10 ng/mL basic fibroblast growth factor (bFGF; Stemgent, Boston, MA) in an humidified incubator at 37 °C with 5% CO2. The cells were passaged weekly enzymatically using 1 mg/mL collagenase IV (Life Technologies). The generation of the NPC line was previously described. 7 Briefly, neural differentiation was initiated by transferring one of the iPSC clones (8330-8) from maintenance on an iMEF feeder layer to feeder-free conditions by growing a high density of cells on 1% Matrigel (354277; BD Biosciences, Franklin Lakes, NJ) substrate and feeding with mTeSR1 media (StemCell Technologies, Reston, VA). Within a couple of weeks, neural rosette structures appeared. The neural rosettes were manually isolated, expanded, and maintained in NPC media as described below. After five passages in NPC expansion media, cells were analyzed for Nestin, SOX1, SOX2, and PSA-NCAM expression by immunocytochemistry. The neuronal differentiation potential of NPCs was evaluated by immunostaining for TuJ1, MAP2, SMI312, and glial fibrillary acidic protein (GFAP).
Culturing Human iPSC-Derived Neural Progenitor Cells
All tissue culture ware (T75 flasks, 6-well, 24-well, 96-well, and 384-well plates) used for culturing human iPSC-NPCs was prepared by a double-coating procedure to provide suitable extracellular factors required for adherence and growth of the iPSC-NPCs. Plates or flasks were first coated with 20 µg/mL polyornithine (Sigma, St. Louis, MO) in ddH2O for 2 h and then with 5 µg/mL laminin (Sigma) in phosphate-buffered Saline (PBS 1×; Gibco, Carlsbad, CA). Coated tissue culture ware could be stored at 4 °C in laminin-PBS for a prolonged period (1–2 weeks) before use.
Media used for human iPSC-NPC culture (NPC media) were composed of 70% DMEM (High Glucose 1×; Gibco 11995), 30% Ham’s F12 with L-glutamine (Modified; Cellgro/Mediatech, Manassas, VA), 1× penicillin/streptomycin, and 1× B27 Supplement (50×, Gibco) and supplemented with 20 ng/mL epidermal growth factor (EGF; Sigma, prepared as 20 µg/mL stock in DMEM), 20 ng/mL bFGF (Stemgent; prepared as 20 µg/mL stock in PBS), and 5 µg/mL heparin (Sigma; prepared as 5 mg/mL stock in Ham’s F12 media) just before use.
Human iPSC-NPCs were maintained in complete NPC media at 37 °C with 5% CO2 in a humidified atmosphere and split twice per week. For passaging, confluent cultures in T75 flasks were washed once with 10 mL PBS and then treated with 1 mL TrypLE Select (Invitrogen, Carlsbad, CA) until cells detached. TrypLE treatment was stopped by adding 9 mL NPC media, and cells were gently triturated multiple times to obtain a single-cell suspension followed by centrifugation at 1000 rpm (700× g) for 5 min and then resuspended gently in complete NPC media. For maintenance, cells were routinely passaged at a 1:3 ratio, or 4 × 106 cells were allocated to one T75 flask and 0.4 × 106 cells/well to a six-well plate.
Creation of TCF/LEF Reporter Line in Human iPSC-Derived NPCs
On day 0, NPCs (initially at passage 42) were seeded at 76 000 cells/well in a polyornithine/laminin-coated 24-well plate. The next day (day 1), a Cignal T cell factor/lymphoid enhancer factor (TCF/LEF) luciferase reporter consisting of tandem TCF/LEF binding sites (containing the sequence AGATCAAAGGGGGTA) with a cytomegalovirus (CMV) minimal promoter and a separate human phosphoglycerate kinase (PGK) promoter that constitutively drives the expression of a puromycin resistance gene packaged into a vesicular stomatitis virus G protein (VSV-G) pseudotyped lentivirus (Qiagen, Valencia, CA) was added to the NPCs at a multiplicity of infection (MOI) = 10. After mixing, the NPCs were spun at 2000 rpm (930× g) for 30 min, before placing the plate of cells in the incubator for 24 h. On day 2, the media were replaced with fresh NPC media, and on day 3, the media were replaced with NPC media containing 0.8 µg/mL puromycin (Sigma) to start selection of the transduced cells. The transduced NPCs were fed with fresh 0.8 µg/mL puromycin in NPC media every 3 days until confluent, then passaged into a well of a polyornithine/laminin-coated six-well plate. Subsequently, the stable TCF/LEF reporter NPC line was maintained similarly to naive NPC cells with the exception of inclusion of 0.8 µg/mL puromycin in the NPC media. After expanding for 3 weeks, the stable TCF/LEF reporter NPC line was cryogenically preserved, and frozen stocks were subsequently used for assays.
Production of NPC-Compatible Wnt3a-Conditioned Media
Wnt3a is a ligand that stimulates the Wnt/β-catenin signaling pathway. To stimulate the Wnt/β-catenin signaling pathway in our TCF/LEF reporter assay, we added a low dose of Wnt3a-conditioned media (Wnt3a-CM) before treatment. Wnt3a-CM was prepared from mouse L cells (CRL-2648; American Type Culture Collection [ATCC], Manassas, VA) stably transfected with a Wnt3a-expressing vector, 15 and Wnt3a was secreted into culture media. The L cells were cultured in DMEM, 10% fetal bovine serum (FBS), and 1× penicillin/streptomycin. To prepare large quantities of Wnt3a-CM for use in HTS, we allowed Wnt3a-secreting L cells to grow to 90% confluency in multiple 150-mm tissue culture dishes, at which stage fresh media replaced the old media (day 0). Culture media were collected and replaced with fresh media every 2 days for three collections (days 2, 4, and 6). Collected media were spun at 3000 rpm for 15 min to remove cellular debris. Supernatant was stored at 4 °C until the final batch was collected, when three batches of supernatant were combined, aliquoted, and stored at −80 °C. The activity of collected Wnt3a-CM was measured in the TCF/LEF reporter assay in a dose-response curve from which EC25 was calculated.
TCF/LEF Luciferase Reporter Assay for Wnt/β-Catenin Signaling
White 96-well (Corning 3903; VWR, West Chester, PA) or 384-well (Corning 3707; VWR) microplates were used in the assay. Plates were prepared with the polyornithine/laminin coating procedure described above with robotic handling, including aspiration with a Power Washer PW384 (Tecan US, Inc., Durham, NC) and dispensing with a Matrix WellMate microplate dispenser (Thermo Scientific, Hudson, NH), to provide standardized operation. For 96-well plates and 384-well plates, 50 µL of coating reagents or 20 µL per well was used, respectively. A quick spin step was included after dispensing of each coating reagent and later cell suspension for 384-well plates to ensure that all liquid was brought down to the bottom of wells. Before cell seeding, coating reagent was removed through a wash with NPC media.
For assays, single-cell suspensions were prepared from the human iPSC-NPCs, stably integrated with the TCF/LEF-luciferase reporter, and dispensed into 96- or 384-well plates at the seeding densities of 20 000 or 6000 per well, respectively, using a Matrix WellMate (Thermo Scientific) microplate dispenser. A 96-well plate format was used for measuring Wnt3a and lithium (lithium chloride solution, 10 M; Fluka, St. Louis, MO) dose-response effects, whereas a 384-well plate format was used for all small-molecule probe treatments and chemical screening. The next day, cells were subjected to various 24-h treatments. Just prior to the luminescence reading, the plates were taken out of a 37 °C incubator and equilibrated to room temperature for 30 min before the SteadyGlo reagent (Promega, Madison, WI) was dispensed (volume equal 1:1 SteadyGlo:culture medium). Luminescence was measured after a 10-min incubation using an EnVision multilabel plate reader (PerkinElmer, Waltham, MA). To obtain dose-response curves for Wnt3a-CM or lithium, we treated cells in 96-well plates with a range of concentrations of Wnt3a-CM or lithium. The EC25 concentration of Wnt3a-CM was calculated from the dose-response curve and used for stimulation of Wnt signaling in subsequent experiments. To look at the additive or synergistic effect with lithium, we included 10 mM or 20 mM lithium in combination with other compound treatment. Dose-response tests for small-molecule compounds were performed in 384-well plates with 10 doses in 2-fold dilution series, with or without EC25 Wnt3a-CM stimulation or lithium. One hour after the addition of Wnt3a-CM, lithium, or media, 50 nL of each compound arrayed in compound plates was transferred to cell microplates using the CyBi-Well vario pinning robot (CyBio Corp., Jena, Germany). GraphPad Prism software (GraphPad Software, La Jolla, CA) was used for graphing and EC50 calculations.
Z-Factor Determination
Z-factor 16 calculations were used to determine the robustness and reproducibility of the assay. Z-factors were calculated either by using media as a negative control and Wnt3a-CM as a positive control or based on DMSO (negative control) and 0.625 µM CHIR-99021 (positive control) in the presence of EC25 Wnt3a-CM.
HTS of Bioactive Compounds
Approximately 1500 compounds from a library of Food and Drug Administration (FDA)–approved drugs and known bioactives composed of 1- to 10-mM stock solutions in DMSO were screened to identify novel small molecules capable of increasing TCF/LEF reporter activity. Cells were plated on polyornithine/laminin-coated 384-well plates and incubated overnight. The screen was performed in three conditions: media, EC25 Wnt3a-CM, or 10 mM lithium. Before compound treatment, 10 µL of media, 10 µL of 72% Wnt3a-CM, or 10 µL of 40 mM lithium was dispensed into 30 µL media in each well to achieve a final volume of 40 µL media, a final EC25 concentration of 18% Wnt3a-CM in media, or a final concentration of 10 mM lithium in media, respectively. One hour after the addition of media, Wnt3a-CM, or lithium, 50 nL of each compound from the library was transferred to the cell microplates using the CyBi-Well vario (CyBio Corp.) pinning robot equipped with a 384-pin array. After 24-h treatment with compounds, plates were taken out of the 37 °C incubator and equilibrated to room temperature for 30 min before 15 µL of SteadyGlo reagent was dispensed and luminescence was measured as described above.
Immunocytochemistry
To assess NPC identity and neural differentiation potency, we fixed proliferative NPCs or subsequent terminally differentiated neuronal cultures (after the removal of the mitogens EGF and bFGF for 10 days) in 4% paraformaldehyde and immunostained them with rabbit anti-NESTIN (AB5922; EMD Millipore, Billerica, MA), rabbit anti-SOX1 (AB15766; EMD Millipore), rabbit anti-SOX2 (AB59776; Abcam, Cambridge, UK), or mouse anti-PSA-NCAM (MAB5324; EMD Millipore) for NPCs, as well as mouse anti-TuJ1 (T8660; Sigma), chicken anti-MAP2 (CPCA-MAP2; EnCor Biotechnology, Inc., Gainesville, FL), mouse anti-SMI312 (SMI-312R; Covance, Princeton, NJ), or mouse anti-GFAP (MCA-5C10; EnCor Biotechnology, Inc.) for neuronal cultures. After primary antibody incubation, cells were washed three times with PBS and followed by the incubation with appropriate fluorochrome-conjugated secondary antibodies. After the final three washes with PBS, neural marker staining was evaluated on a Zeiss Axiovert fluorescence microscope with 10× or 20× objectives equipped with a Zeiss Axiocam digital camera (Carl Zeiss, Jena, Germany).
Results
Expandable, Homogeneous Human iPSC-Derived Neural Progenitors as a Platform for HTS
The homogeneous human NPC line, 8330-8, was derived from iPSCs generated from fibroblasts isolated from a healthy human adult male as detailed in our recently published study. 7 The process included reprogramming the human fibroblasts using the four now classical Yamanaka transcription factors (OCT4, SOX2, KLF4, and c-MYC) into pluripotent stem cells,17,18 followed by neural induction and rosette isolation ( Fig. 1A ). The resulting NPC line was immunopositive for Nestin, SOX1, SOX2, and PSA-NCAM, a group of markers indicative of neural progenitor cell fate ( Fig. 1B ). These iPSC-derived NPCs rapidly self-renew and can be expanded and propagated for extended periods (upwards of 60+ passages) without incurring genomic instability. Prior to initiating the generation of stable reporter lines, the ability of the iPSC-NPCs to terminally differentiate into neurons and glia was confirmed by withdrawal of mitogenic factors (EGF, bFGF) and subsequent immunocytochemistry for lineage-specific markers TuJ1 (neural), MAP2 (dendritic), SMI312 (axonal), and GFAP (glial) ( Fig. 1C ). We also found that the NPCs, with prolonged differentiation, give rise to morphologically complex, polarized neurons that are electrically active (data not shown). Therefore, once successfully isolated, these human iPSC-derived NPCs can be expanded significantly to provide the billions of cells needed for a large-scale HTS.
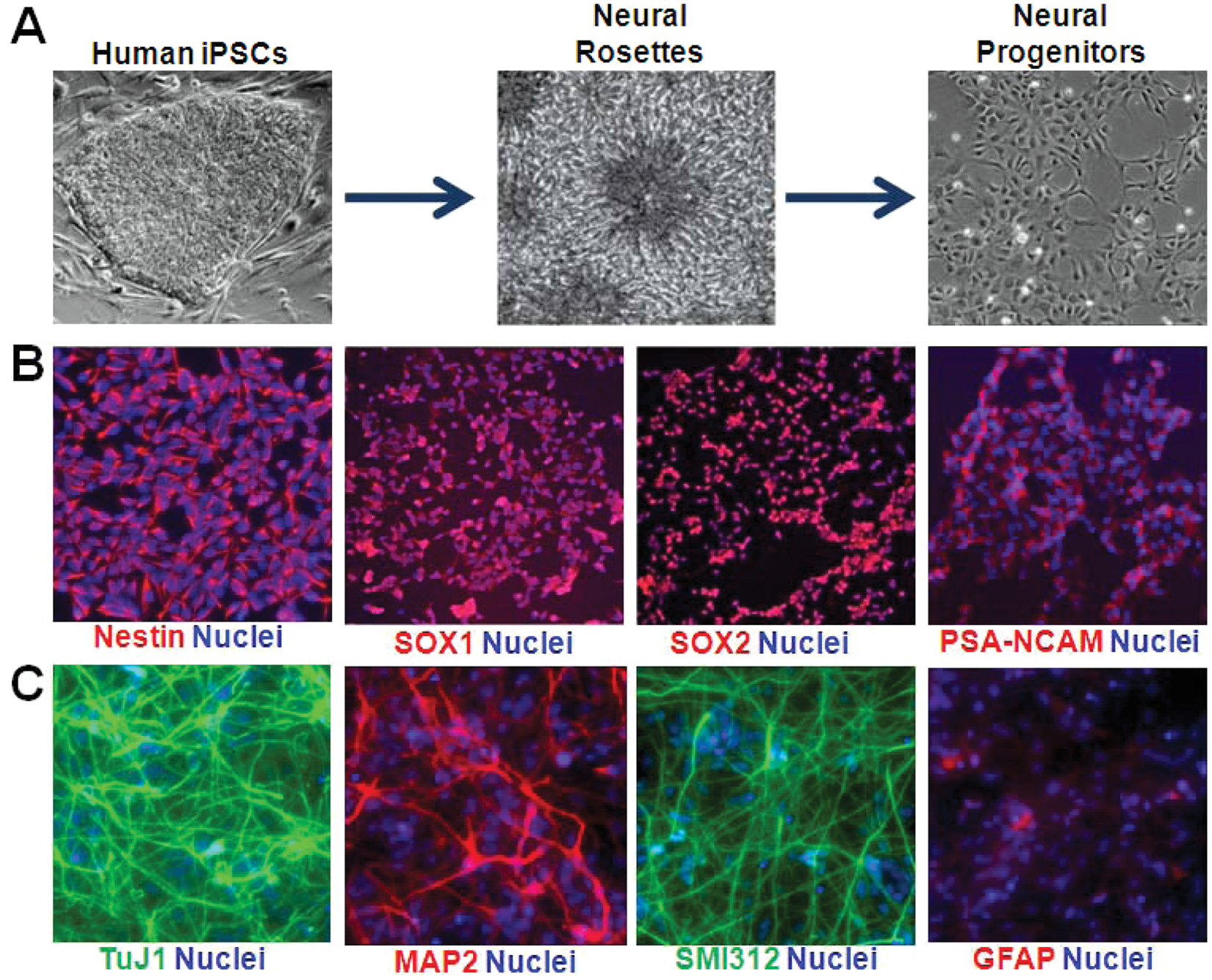
Derivation of homogeneous induced pluripotent stem cell (iPSC)–derived expandable neural progenitor cells (iPSC-NPCs). (
Development of a Wnt/β-Catenin Signaling Reporter Assay in Human iPSC-Derived NPCs
The identification of small-molecule probes that modulate the Wnt/β-catenin signaling pathway in human patient–specific NPCs can facilitate a better understanding of the regulation of neurogenesis and the pathophysiology of neuropsychiatric disorders. Previously, we and others have used luciferase reporters driven by promoters containing LEF/TCF family transcription factor binding sites to successfully identify small molecules affecting the Wnt/β-catenin signaling pathway in tumor/immortalized cell lines in large-scale screens. 19 In an effort to discover small molecules affecting the Wnt/β-catenin signaling pathway in the context of human neural stem cells and neurons, we sought to generate a stable TCF/LEF reporter NPC line derived from iPSCs that could be used in HTS.
As the lentiviral system has been successful in introducing transgenes into many cell types, including cell types that have been difficult to transfect using lipid-mediated methods, our approach was to introduce the TCF/LEF reporter into NPCs using VSV-G pseudotyped lentiviral particles. The TCF/LEF luciferase reporter we used contains the firefly luciferase gene under the control of tandem repeats of the TCF/LEF transcriptional response element, which binds the TCF/LEF family of high-mobility group (HMG) domain-containing transcription factors, together with a CMV minimal promoter. This vector also contains a separate human PGK promoter that constitutively drives the expression of a puromycin resistance gene and thus can be used to select for transduced cells in stable cell line generation. Initially, we tested the sensitivity of the 8330-8 NPCs to various amounts of the lentivirus with or without the addition of polybrene or protamine sulfate, agents used to increase the efficiency of viral transduction (
Assay Sensitivity to Known Wnt/β-Catenin Signaling Pathway Modulators
To validate our NPC TCF/LEF-luciferase reporter system ( Fig. 2A ), we first tested several known Wnt/β-catenin signaling pathway modulators (Wnt3a-CM, lithium, and adenosine triphosphate [ATP]–competitive GSK3β inhibitors) ( Fig. 2 ). From the dose-response curve of Wnt3a-CM ( Fig. 2B ), we calculated the effective concentration that induces a 25% increase of reporter activity (EC25), which was subsequently used in our high-throughput screen. We observed dose-responsive effects from lithium ( Fig. 2C ), although its effect was relatively modest compared with Wnt3a-CM or CHIR-99021, a potent GSK3β inhibitor ( Fig. 2D ). These data indicate that the reporter system we developed provides a suitable system for screening for Wnt/β-catenin signaling modulators.
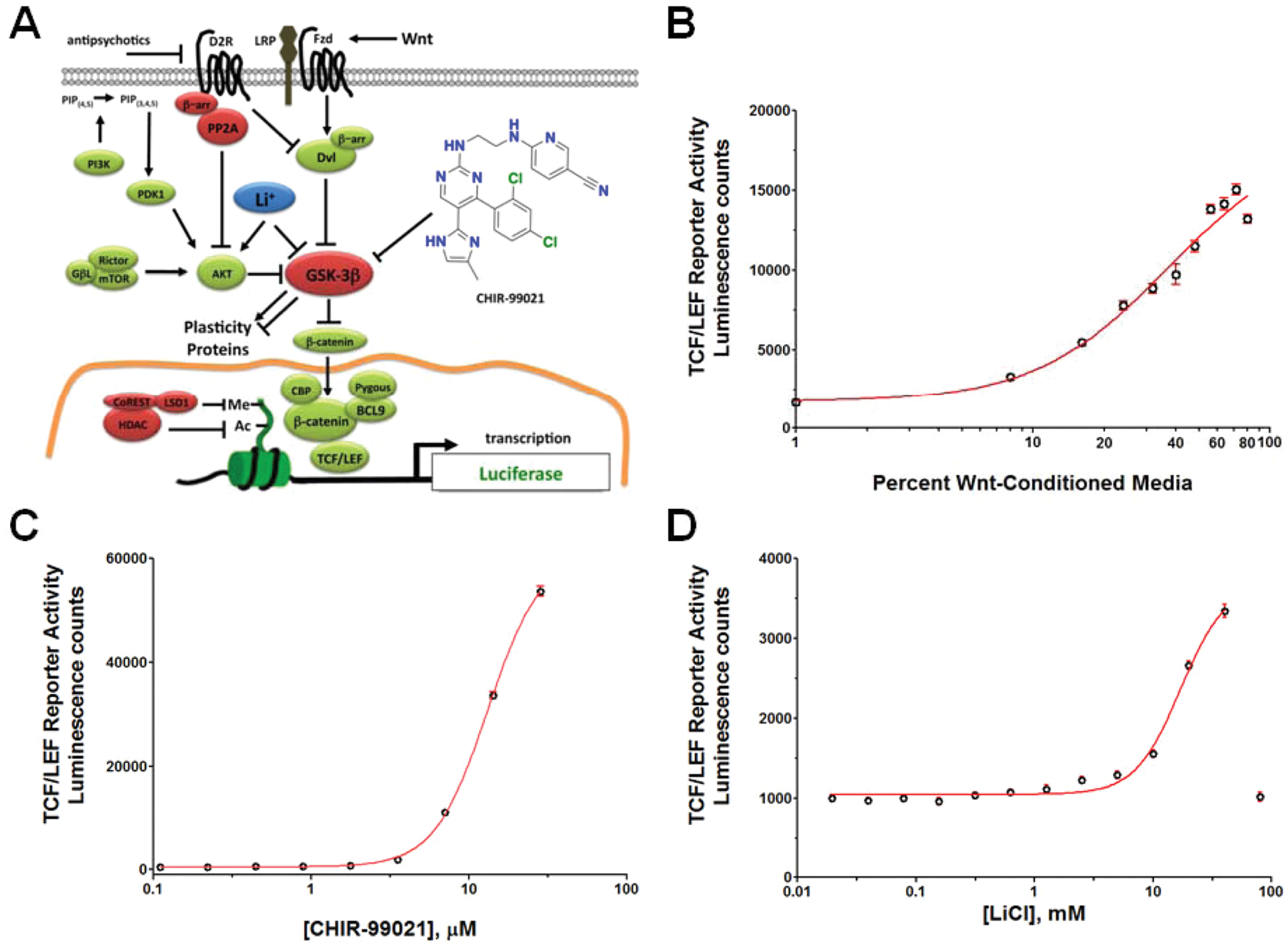
T cell factor/lymphoid enhancer factor (TCF/LEF) luciferase reporter system for Wnt/β-catenin signaling in human neural progenitor cells (NPCs). (
Assay Parameters and Robustness
To determine the robustness and reproducibility of the TCF/LEF reporter assay over time, we treated cells with (1) media alone, (2) 10 mM lithium, or (3) EC25 Wnt3a-CM. Treatment of TCF/LEF reporter cells with 10 mM lithium for 24 h resulted in a 2- to 5-fold increase of TCF/LEF reporter activity, whereas treatment of EC25 Wnt3a-CM produced activation ~50-fold. These values were reproducible in repeated assays conducted over multiple weeks ( Fig. 3A ). Under these conditions, the coefficient of variation observed for the EC25 Wnt3a-CM condition was ~10%, whereas the coefficient of variation was ~20% for the condition of media or 10 mM lithium. If Z-factors are calculated based on DMSO (negative control) and 0.625 µM CHIR-99021 (positive control) under the condition of EC25 Wnt3a-CM, we could routinely observe Z-factor values above 0.75. Using media as a negative control and Wnt3a-CM as a positive control, we consistently observed Z-factor values between 0.55 and 0.75. CHIR-99021, the positive control, showed a further ~5-fold activation in the presence of EC25 Wnt3a-CM. Thus, the assay is highly robust and suitable for HTS to detect Wnt3a potentiators.
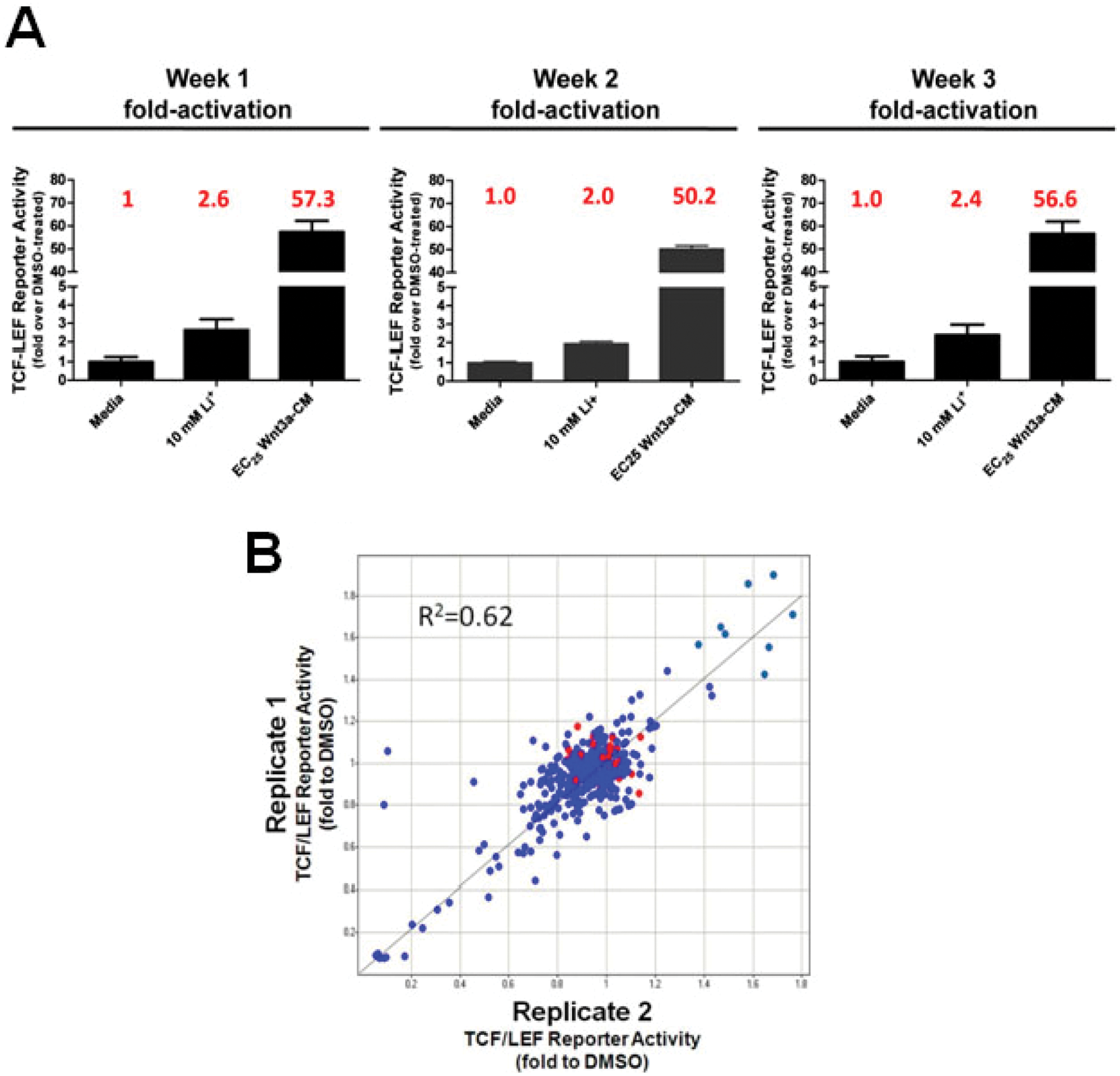
Reproducibility of the T cell factor/lymphoid enhancer factor (TCF/LEF) reporter assay in human neural progenitor cells (NPCs). (
HTS of Known Bioactives
Using our validated TCF/LEF reporter system to perform HTS, we first determined the reproducibility of data generated from two replicates of a 384-well plate with compounds transferred robotically. As shown in Figure 3B , there was overall a correlation of 0.62 between the two replicates.
We next performed a screen of ~1500 compounds from a library of FDA-approved drugs and known bioactives to identify novel small molecules capable of increasing TCF/LEF reporter activity ( Fig. 4 ). Compound activity was assessed in the absence or presence of EC25 Wnt3a-CM or 10 mM lithium. In the EC25 Wnt3a-CM screen using 10 µM CHIR-99021 as the positive control, a Z-factor of 0.75 was obtained. In the presence of EC25 Wnt3a-CM, we found 45 compounds that induced a ≥1.5-fold increase of TCF/LEF reporter activity and 61 compounds that decreased reporter activity to ≥0.5-fold, which corresponded to ± ~3 SD beyond the median of the DMSO values. This high overall hit rate is likely due to the fact that the library is composed of known bioactives. Under the conditions of media alone or 10 mM lithium, a smaller number of compounds were scored as hits, and most were also present in the group identified under the condition of EC25 Wnt3a-CM. Therefore, we proceeded to retest compound dose responses with the hits from the condition of EC25 Wnt3a-CM. Of note, although our screen also identified inhibitors of Wnt/β-catenin signaling, we focused on compounds that activated Wnt/β-catenin signaling, which we retested in a 10-point dose-response series.
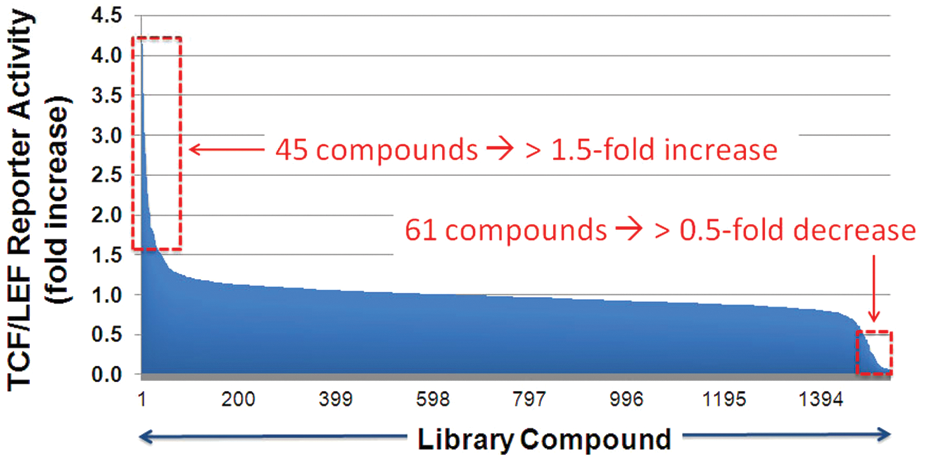
High-throughput screen for Wnt/β-catenin signaling modulators. The screen was conducted in three conditions: media, EC25 Wnt3a-CM (shown here), or 10 mM lithium. TCF/LEF, T cell factor/lymphoid enhancer factor.
Riluzole and mGluR1 Antagonists as Wnt/β-Catenin Signaling Activators
Reassuringly, riluzole, an FDA-approved drug that we had previously identified as a Wnt/β-catenin pathway potentiator in our mouse HT-22 neural progenitor cell line, which has activity that is dependent on intact TCF/LEF binding sites in the reporter gene,
19
was among the 45 compounds that increased TCF/LEF reporter activity in our NPCs. To confirm the result, we retested riluzole in doses under the conditions of both EC25 Wnt3a-CM and 20 mM lithium. Repeatedly, EC25 Wnt3a-CM induced a ~50-fold increase over baseline on the TCF/LEF reporter activity, whereas that for 20 mM lithium was ~4-fold. In our dose-response retest, riluzole potentiated the effects of Wnt and lithium (
Fig. 5A
) with an EC50 of ~5 µM (
Fig. 5B
). As riluzole was reported to enhance Wnt/β-catenin signaling through an indirect antagonism of metabotropic glutamate receptor 1 (mGluR1),
19
we tested four mGluR1 antagonists (LY456236, YM298198, Bay 36-7620, and JNJ16259685) in the TCF/LEF reporter assay (
Fig. 5C
). In the presence of EC25 Wnt3a-CM, we found that two potent mGluR1 antagonists (YM298198 and JNJ16259685) increased TCF/LEF reporter activity in a dose-dependent manner, with an EC50 of ~3 µM or 0.7 µM, respectively (
Fig. 5D
). Although LY456236 and Bay 36-7620 were reported to be active in the TCF/LEF reporter system in melanoma cells,
19
we failed to detect their activities in our reporter system in human iPSC-NPCs, indicating the potential difference between these two different cell types. YM298198 and JNJ16259685 also potentiated lithium activation of the TCF/LEF reporter gene (
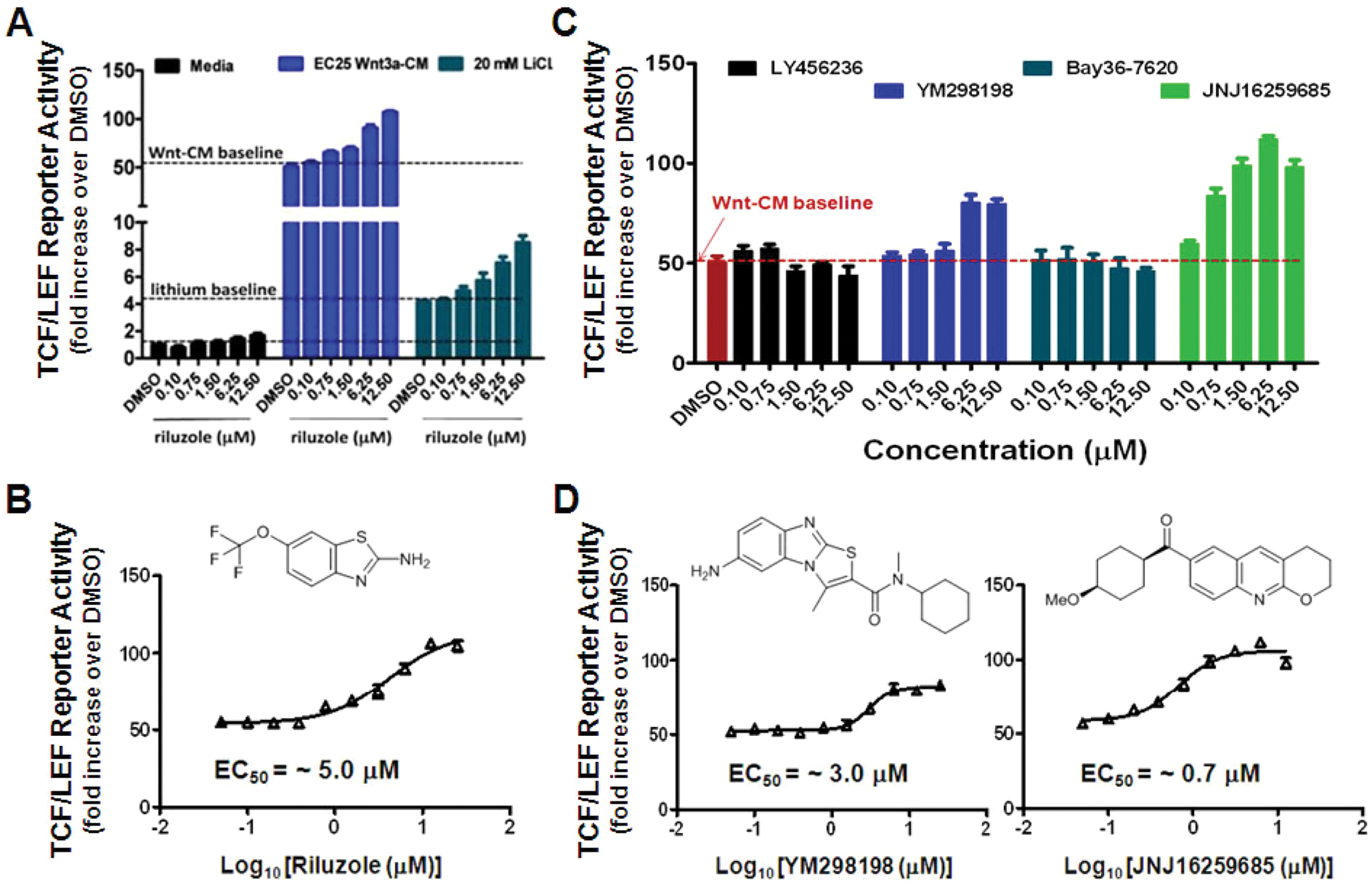
Riluzole and mGluR1 antagonists in the T cell factor/lymphoid enhancer factor (TCF/LEF) reporter assay. Riluzole, a reported Wnt/β-catenin signaling potentiator and mGluR1 antagonist, was among the high-throughput screening hits. (
Retest of Potential Wnt/β-Catenin Signaling Activators
In addition to riluzole, a confirmed Wnt/β-catenin signaling potentiator, we also retested the remainder of the hit candidates in our iPSC-NPC TCF/LEF reporter assay under the conditions of both EC25 Wnt3a-CM and 20 mM lithium. Hit compounds were resupplied as dry powders from commercial vendors and reformatted into 10-dose compound plates. All hit compounds showed additive effects to Wnt3a in a dose-dependent manner, whereas most, but not all, were additive to lithium as well.
Figure 6
shows five representative retested hit compounds, all of which showed a dose-dependent additive effect to Wnt3a, and among which (–)-eseroline was inactive in the presence of 20 mM lithium (
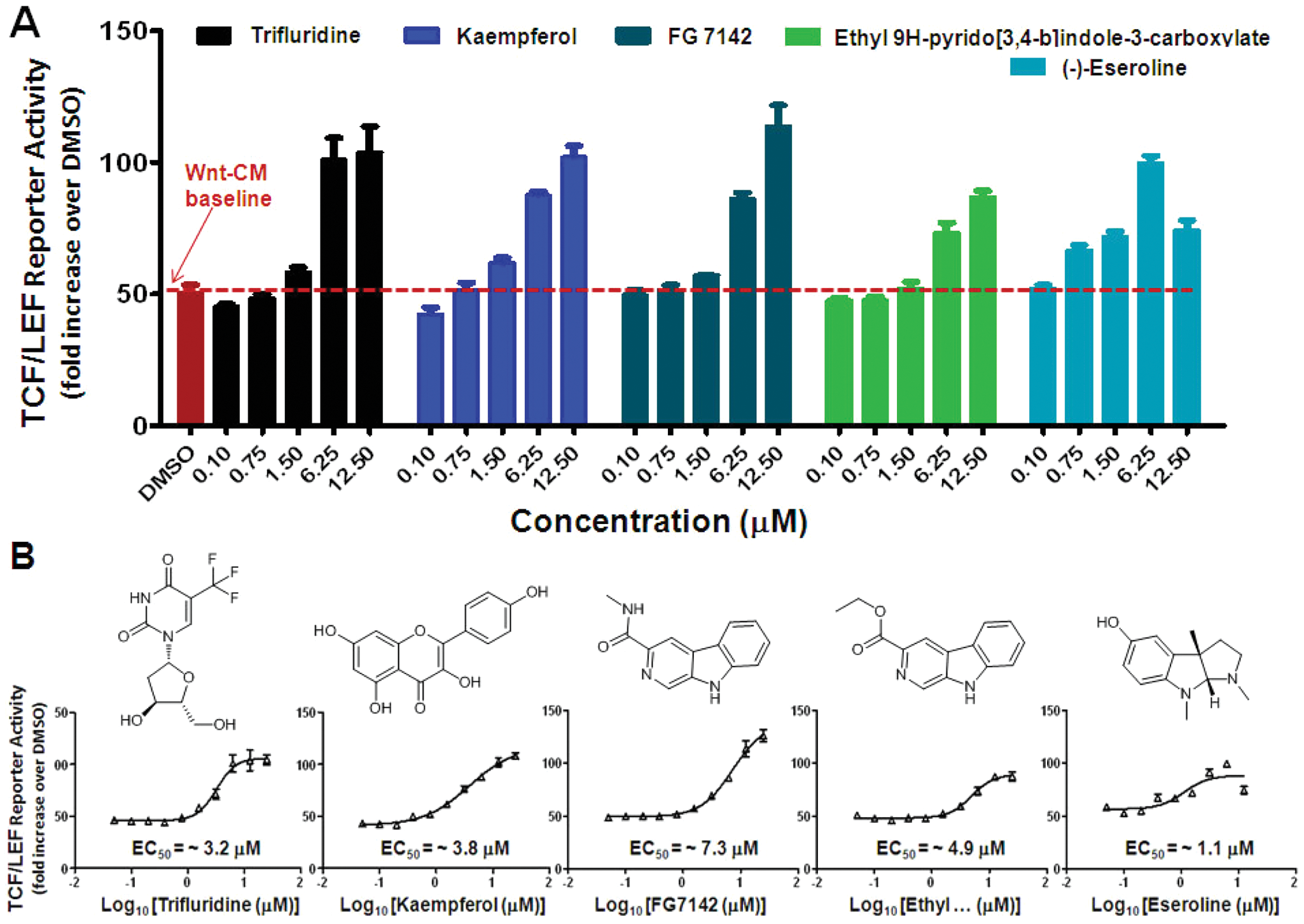
Dose-response retest of representative high-throughput screening hits. (
Discussion
Wnt/β-catenin signaling has been shown to play an important role in the regulation of the proliferation of neural progenitors during embryonic brain development and adult neurogenesis and subsequently in neural differentiation and other aspects of neuroplasticity.12,20,21 Here we show, for the first time, that human iPSC-derived NPCs have an intact functional Wnt/β-catenin signaling pathway capable of responding to Wnt3a, direct GSK3 inhibitors such as CHIR-99021, and a variety of known and novel small molecules, including mGluR1 antagonists, resulting in the transcriptional activation of a synthetic Wnt/β-catenin reporter gene containing TCF/LEF response elements that bind the TCF/LEF family of HMG domain-containing transcription factors.
Several lines of evidence implicate the Wnt/GSK3 pathway in the etiology of neuropsychiatric disorders. First, this pathway is targeted by pharmacological agents used to treat these disorders.22–24 In addition, the Wnt/GSK3 pathway has further been implicated in the etiology of neuropsychiatric disorders by the demonstration of inhibition of GSK3 by the schizophrenia-associated gene DISC1 21 and the recent discovery of genetic variation of TCF4, which is a known target gene of β-catenin, as a susceptibility locus for schizophrenia and a known cause of intellectual disability in the case of Pitt-Hopkins syndrome haploinsufficiency. 25 Accordingly, the identification of small-molecule probes that enhance Wnt/β-catenin signaling and elicit a synergistic/additive effect to lithium would be valuable tool for probing the role of Wnt/β-catenin signaling in neuroplasticity and the pathophysiology of neuropsychiatric disorders.
We note both shared responses of certain compounds (e.g., CHIR-99021, riluzole) between our iPSC-derived NPC TCF/LEF reporter line and previous mouse and nonneural human TCF/LEF reporter lines, 19 as well as different responses, as in the case of certain mGluR1 antagonists. Although we cannot rule out differences in factors such as metabolism or assay sensitivity, it is conceivable that our human NPC reporter system reveals previously unidentified responses intrinsic and novel to this system, thus providing further support for the use of the iPSC-derived NPC model.
Having demonstrated the feasibility of identifying compounds that appear to modulate Wnt/β-catenin signaling in human iPSC-NPCs, we note that a critically important next step in the screening process will be to validate the hits with additional secondary assays addressing the ability of the compounds to modulate endogenous TCF/LEF-mediated transcription and to have downstream consequences on pathways involved in neurogenesis and neurodevelopment. Of particular interest will be to determine if any of the compounds identified as modulators of Wnt/β-catenin signaling are specific to human iPSC-NPCs as compared with other human cell types (e.g., immortalized cell lines or cancer cell lines). Identifying Wnt/β-catenin signaling modulators that are inactive in tumor-derived cells, for example, may point to particularly attractive targets for modulating Wnt/β-catenin signaling in the CNS. Also of interest will be to compare the ability of compounds to activate Wnt/β-catenin signaling in differentiated postmitotic neurons. Here, it may be feasible to identify Wnt/β-catenin modulators only active in the proliferative neural progenitors or, conversely, if the screen is performed on differentiated neurons first, to identify modulators only active on postmitotic neurons.
Since Wnt/β-catenin signaling has been shown to play an important role in regulating mammalian neurogenesis and neurodevelopment, 20 we anticipate that performing large-scale HTS will lead to critically needed probes of the underlying molecular pathways that can be used both in the in vitro studies with human and rodent neural progenitors but also, after further optimization, as probes of these pathways in vivo. Most broadly, through the reversal or compensation of deficits, the modulation of postnatal and adult neurogenesis has been proposed as a potential therapeutic avenue for multiple neuropsychiatric and neurodegenerative disorders, including, but not limited to, bipolar disorder, schizophrenia, major depression, traumatic brain injury, Alzheimer disease, Parkinson disease, and Huntington disease.26,27 For example, recent studies in the context of frontotemporal dementia have implicated dysregulation of Wnt signaling involving the Frizzled 2 (FZD2) receptor 14 as being involved in the underlying pathophysiology, with evidence that potentiators, such as the ones discovered here, may be therapeutically relevant.
Although rosette picking was used as the initial procedure to generate and isolate NPCs from neural differentiated iPSC cultures in our studies, an important consideration is that once isolated and manually passaged more than five passages, the NPC line presented in this study (8330-8) was homogeneous (by morphology and marker staining as presented) and expanded significantly to provide the billions of cells needed for a large-scale HTS. However, we note that there is growing evidence for the existence of multiple types of NPCs within differentiating iPSC cell cultures. 28 As the patterning capability of long-term expandable iPSC-NPCs as currently cultured appears to become restricted to a more ventral, hindbrain fate,29,30 an important avenue for future studies will be to further exploit the plasticity of iPSC model systems to identify and refine culture conditions to enable the continued expansion of both dorsal and ventral telencephalic progenitors. Achieving this objective will provide access to additional progenitor subtypes and ultimately allow generation of functional, subtype-specific neurons en masse that can be used for HTS and a variety of functional genomic studies.
In summary, the use of human, patient-specific iPSC models and the subsequent derivation and expansion of progenitor cells of defined lineages provide a platform scalable in terms of the number of cells, genotypes, disease models, and phenotypes. Once expanded to capture additional patient genetic diversity, as well as other types of CNS progenitors and disease-relevant phenotypes, this platform can support the objectives of the field of human experimental neurobiology for (1) functional characterization of pathophysiological mechanisms; (2) screening for novel therapeutics using physiologically relevant, defined cell subtypes, such as the NPCs described here; and (3) the identification of novel targets and mechanisms for therapeutic intervention, such as mGluR1 antagonists for potentiating Wnt/β-catenin signaling described here. Future studies will aim to systematically assess whether there are differences in the response of iPSC-NPCs from patients to modulators of Wnt/β-catenin signaling, including the panel of iPSCs models that we and others are developing from patients with bipolar disorder, schizophrenia, and Fragile X syndrome (e.g., Sheridan et al. 7 )—disorders in which there exists evidence for dysregulated GSK3 signaling that may be causally involved in the underlying pathophysiology.
Footnotes
Acknowledgements
We thank members of the Haggarty laboratory, the Broad Institute Chemical Biology Platform, Roy Perlis (MGH Department of Psychiatry), and Randall Moon (University of Washington, Seattle) for helpful discussions.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The project described was supported by grant numbers R33MH087896 (SJH) and R01MH091115 (LHT) from the National Institutes of Health (NIH) and National Institute of Mental Health (NIMH). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Mental Health or the National Institutes of Health. Funding support was also provided in part by the FRAXA Research Foundation and Marigold Foundation (SJH) and the Stanley Medical Research Institute (LHT, SJH).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.