
Abstract
The SWI/SNF-like adenosine triphosphate (ATP)–dependent chromatin remodeling complex, esBAF, is both necessary and, in some contexts, sufficient to induce the pluripotent state. Furthermore, mutations in various BAF subunits are associated with cancer. Little is known regarding the precise mechanism(s) by which this complex exerts its activities. Thus, it is unclear which protein interactions would be important to disrupt to isolate a relevant readout of mechanism. To address this, we developed a gene expression–based assay to identify inhibitors of the native esBAF complex. Specifically, a quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) assay was developed in mouse embryonic stem (ES) cells to monitor expression of Bmi1, a developmentally important gene repressed by the esBAF complex. The assay was miniaturized to a 384-well format and used to screen a diverse collection of compounds, including novel products of diversity-oriented synthesis (DOS). Confirmed hits were validated using a knock-in ES cell reporter line in which luciferase is inserted into the Bmi1 locus. Several of the validated hits regulate a panel of target genes in a manner similar to the BAF chromatin-remodeling complex. Together these data indicate that expression-based screening using qRT-PCR is a successful approach to identify compounds targeting the regulation of key developmental genes in ES cells.
Introduction
An essential component of the embryonic stem (ES) cell core pluripotency transcriptional network is the SWI/SNF-like adenosine triphosphate (ATP)–dependent chromatin-remodeling complex 1 ; several of the subunits have a genetically dominant role in ES cell development and formation of the inner cell mass.2,3 In addition, the complex is necessary and, in certain contexts, sufficient for the induction of pluripotency. 4 Mammalian SWI/SNF complexes are composed of a central ATPase (BRG1 or BRM) and 10 to 12 subunits (referred to as BAFs for BRG1 or BRM-associated factors). The 11 members are encoded by 20 genes, which exhibit combinatorial assembly for an astounding 288 predicted combinations. Indeed, ES cells express a unique assembly of subunits, called esBAF, which cannot be functionally rescued by the expression of alternate, homologous subunits ( Fig. 1 ). The combination of BAF subunits is cell type specific, and subunit switching is an important determinant of differentiation. 5
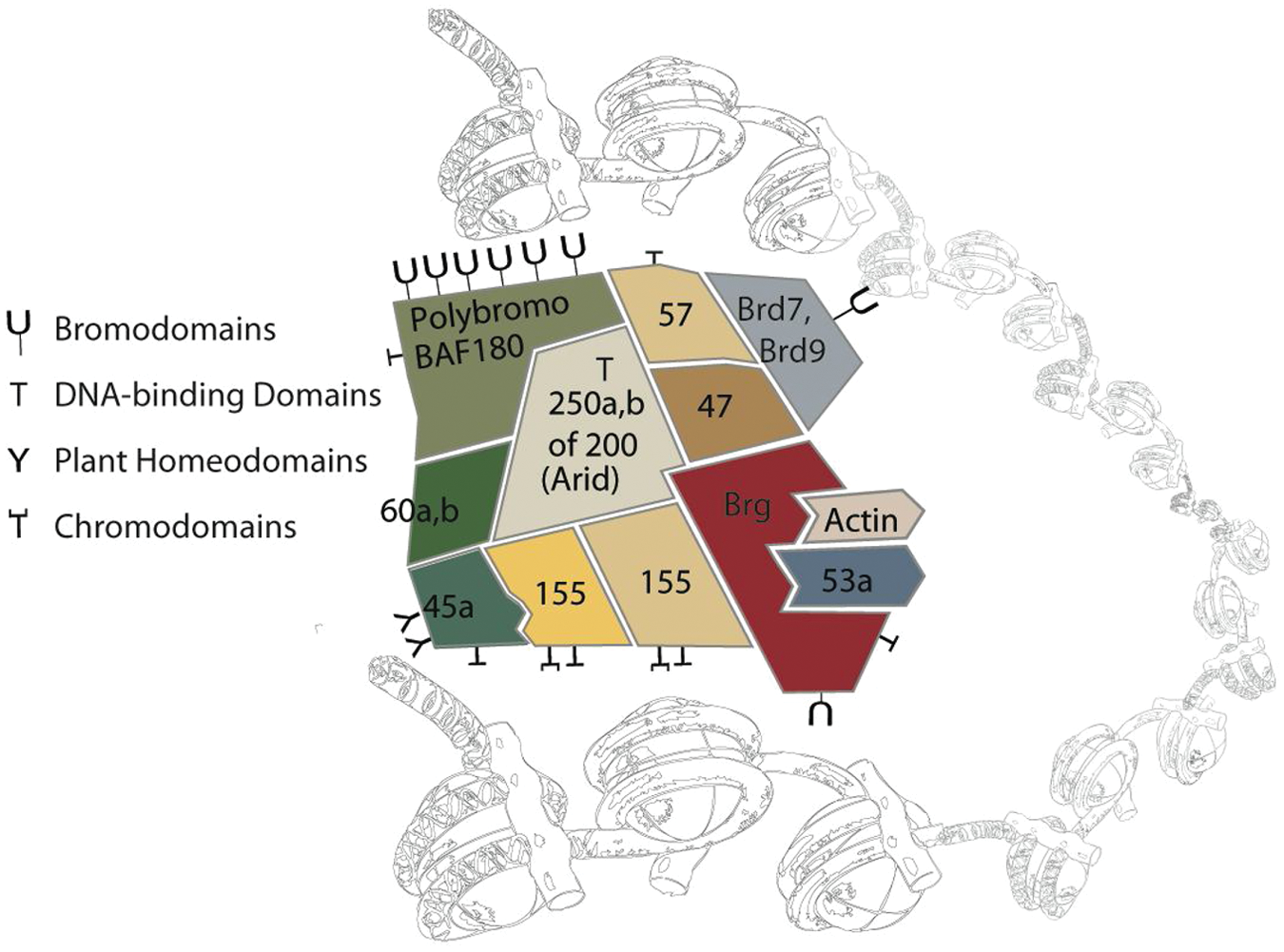
The esBAF complex associated with chromatin. The specificity imparted by the particular composition of BAF subunits in embryonic stem (ES) cells is in part through the combinatorial assembly of chromatin targeting motifs, including DNA-binding domains, bromodomains, chromodomains, and plant homeodomains.
In addition to the essential role of BAF in pluripotency and development, several BAF subunits have been confirmed as tumor suppressors in humans. As of now, malignancy-causing BAF subunit mutations include BAF47 (hSNF5, INI1) in nearly 100% of human malignant rhabdoid tumors 6 ; BAF250a (Arid1a) in ovarian clear cell carcinoma, gastric cancer, and colorectal cancer7,8; BAF180 (polybromo) in renal clear cell carcinoma 9 ; BAF200 (Arid2) in hepatocellular carcinoma 10 ; and Brg1 mutations in meduloblastoma. 11 Cancer-associated perturbations in expression have been reported for Brm, BAF155, BAF60, BAF57, BAF45d, and BAF53a, although, as of yet, they have not been characterized as tumor suppressors. 12 Considering the evidence linking misregulation of the BAF complex to cancer initiation, progression, and therapeutic resistance, further understanding of the mechanism of the BAF complex is crucial for understanding the link between chromatin remodeling and tumor biology. 12
Although SWI/SNF complexes in Saccharomyces cerevisiae exclusively activate genes, microarray studies in mouse ES cells reveal a predominantly repressive role for esBAF (approximately 70% of genes directly regulated by esBAF are repressed). To further complicate matters, esBAF predominantly represses targets at sites distal to the promoters; only 12% of esBAF binding occurs at promoters. 1 It is unclear how the esBAF complex can be acting in such a manner, and previous mechanistic studies have only revealed small glimpses of the whole picture.13–15 Selective and specific small-molecule probes of esBAF activity will be invaluable tools with which to elucidate the mechanism(s) by which this complex chromatin regulator functions; none currently exist.
In vitro, the BAF complex displays DNA-stimulated ATPase activity and can mobilize nucleosomes on a nucleosomal template. The ATPase subunit, BRG1 or BRM, is sufficient for remodeling activity, and the addition of core subunits, BAF47, BAF155, and BAF170, increases remodeling activity to a level observed for the whole complex. 16 The subunits not required for ATP-dependent remodeling in vitro, however, are essential for all of the activities of the complex in vivo, indicating activities for individual subunits beyond nucleosome remodeling. Previous evidence supported the model that the BAF complex mobilized nucleosomes at promoters to create open regions of chromatin for active transcription. 17 However, our evidence supports a more repressive function for the esBAF complex, from a location distal to the promoter. These data indicate that BAF complexes play more complex roles in ES cell gene regulation than previously thought and act via currently unknown mechanisms. We plan to use small-molecule inhibitors to identify and order the series of reactions catalyzed by the esBAF complex. We hypothesize that the temporal control that small-molecule inhibitors provide will be critical in deciphering the elusive mechanism of the BAF complex.
No compound has yet been identified as an inhibitor of an ATP-dependent chromatin-remodeling complex. A number of approaches could be developed for high-throughput screening (HTS) of small-molecule libraries to identify inhibitors of esBAF activity, including both cell-based and biochemical strategies. To maximize the physiological relevance of any hits, we chose to develop a gene expression–based quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) assay as a primary screen for compounds that inhibit esBAF-mediated repression in ES cells. We used the Ambion Cells-to-Ct kit (Life Technologies, Carlsbad, CA) in a 384-well format to multiplex the expression of Bmi1, a developmentally important BAF-repressed gene, with actin in mouse ES cells. We validated confirmed hits using a mouse ES cell knock-in luciferase reporter line for Bmi1 expression. Last, we used qRT-PCR to test the compounds’ ability to regulate the expression level of a panel of esBAF target genes. By screening a library of ~30 000 small molecules, including both novel and pharmacologically active compounds, we identified 20 compounds that transcriptionally mimic the esBAF knockout. Studies are under way to determine whether the compounds act directly on the BAF complex or if they inhibit important transcriptional regulators that act in concert with the BAF complex. Either possibility will lead to deeper understanding of the actions of this chromatin-remodeling complex, which plays an essential role in pluripotency, human tumor suppression, and cellular senescence.
Materials and Methods
Culture of Mouse ES Cells for qRT-PCR Screen
The feeder-free mouse ES cell line, E14, was maintained on gelatin-pretreated tissue culture plates in ES media consisting of high-glucose Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 15% fetal bovine serum (FBS; ES cell qualified; Applied Stem Cell, Menlo Park, CA), 100 µM 2-mercaptoethanol (Invitrogen), 1% minimum essential medium (MEM) nonessential amino acids (Invitrogen), 1 mM Hepes (Invitrogen), 100 U/mL penicillin/streptomycin (Invitrogen), 2 mM GlutaMAX (Invitrogen), 1 mM sodium pyruvate (Invitrogen), and 1 U/mL LIF (Millipore, Billerica, MA).
Development and Culture of Bmi-luc Reporter ES Cell Line
The Bmi-luc reporter line was developed as published with modifications for incorporation of the firefly luciferase gene. 18 Briefly, ES cells derived from TC1 mice were electroporated with an 8-kb construct consisting of homology arms flanking the ATG transcription start site of Bmi1, where the firefly luciferase gene and neomycin were inserted. After selection, knock-in lines were confirmed using Southern blot analysis, and neomycin was removed by transfection with Cre recombinase. Renilla luciferase was inserted into the genome using a lentiviral vector and selection with blastomycin, followed by clonal isolation and propagation to create the final Bmi-luc ES cell reporter line. Bmi-luc ES cells were maintained on a layer of irradiated mouse embryonic fibroblasts (MEFs) and were plated on gelatin-coated 384-well plates for screening.
Compound Library and Screening
White opaque tissue culture–treated 384-well plates (Corning, Corning, NY) were pretreated for 30 min with 0.1% gelatin in PBS (Millipore). The gelatin was removed by aspiration, and 5000 ES cells were plated per well in in a total volume of 50 µL ES media. After plating, the ES cells were incubated for 24 h at 37 °C and 5% CO2. Test compounds (100 nL) were added to ~10 µM final concentration by pin transfer; each compound was tested in duplicate. The compound library comprised ~30 000 compounds from diverse sources, including bioactive compounds (including Food and Drug Administration [FDA]–approved drugs), commercially available drug-like molecules, targeted collections (biased for kinases, chromatin modifiers, etc.), stereochemically diverse compounds, and purified natural products. After compound addition, cells were incubated for 18 h at 37 °C and 5% CO2. The media were aspirated and cells were washed with 100 µL phosphate-buffered saline (PBS). Cells-to-Ct lysis buffer containing DNaseI (10 µL) was added, and plates were agitated for 5 min at room temperature. Cells-to-Ct stop solution was added and plates were incubated for 2 min at room temp (19–25 °C). Meanwhile, Cells-to-Ct reverse transcriptase master mix (8 µL) was dispensed into new PCR plates. Cell lysate (2 µL) was transferred to the PCR plate containing RT master mix, and the plates were incubated at 37 °C for 60 min and then at 95 °C for 5 min in a PCR block. qPCR master mix (4 µL), consisting of Roche Taqman master mix, 40× FAM Bmi1 Taqman probe, and 40× VIC actin Taqman probe (Applied Biosystems, Life Technologies, Carlsbad, CA), was dispensed into 384-well qPCR plates. cDNA from the RT reaction (1 µL) was dispensed into the qPCR plate alongside cDNA isolated from Brg1-depleted ESE14 cells (to serve as a positive control for elevation of Bmi1 transcript levels). qPCR was performed on the Roche Lightcycler (Roche Applied Sciences, Indianapolis, IN) with a 10-min incubation at 95 °C to activate the enzyme, followed by 40 cycles of 1 min at 60 °C and 15 s at 95 °C. Calculations are based on the 2–ΔΔCT calculations described by Livak and Schmittgen, 19 where –ΔΔCT = (CT,Bmi1 – CT, Actin)Treated – (CT,Bmi1 – CT, Actin)DMSO control. The “–ΔΔCT” values were calculated on a per plate basis using 32 neutral control (DMSO-treated) wells and the derived CP values obtained from the Roche Lightcycler, which are akin to CT values. –ΔΔCT was calculated for all compounds, and the −1.33 value for −ΔΔCT was established as a cutoff for hit calling and cherry-picking purposes (this value corresponds to a ~2.5-fold increase in Bmi1 expression). For an in-depth protocol for implementation of high-throughput RT-PCR for small-molecule screening assays, including data analysis, see Phelan et al. 16 and Bittker. 20
Analysis of Bmi1 Induction Using Luciferase Reporter ES Cell Line
Bmi1 luciferase knock-in ES cells were plated in 384-well white opaque tissue culture plates coated with gelatin at a density of 5000 cells per well in 30 µL media. Cells were grown for 24 h at 37 °C and 5% CO2 and pinned with 100 nL test compound. The compound-treated cells were grown for an additional 24 h at 37 °C and 5% CO2 and then equilibrated to room temperature for 1 h. Dual-Glo (Promega, Madison, WI) luciferase reagent (25 µL) was added and plates were incubated for 45 min at room temperature. Firefly luciferase levels were read on an EnVision plate reader (PerkinElmer, Waltham, MA), and 25 µL Dual-Glo renilla luciferase reagent was added. Plates were incubated for 15 min at room temperature, and renilla luciferase levels were read. Measurements were calculated as a ratio of firefly and renilla luciferase levels.
Additional Analysis of Brg1 Targets by qRT-PCR
cDNA (1 µL) obtained by RT reactions from lysates of ES cells treated with hit compounds (used at the concentration that gave the maximal Bmi1 induction during the concentration-response confirmation studies) was screened using SYBR Green for Bmi1, as well as additional BRG1 targets, Phox2b, Fgf4, Bmp4, Socs3, Cbx7, Ring1a, and Eed. Gapdh was used as the housekeeping gene. Melting curves and sequencing were performed for each set of primers to confirm specificity.
List of SYBR Green qPCR Primers
Brg1: Forward: CGGTTGTGAGTGACGATGAC; reverse: TCTGGTGCTACCCATCACTG
Bmi1: Forward: TACCATGAATGGAACCAGCA; reverse: AAAGGAAGCAAACTGGACGA
Phox2b: Forward: CAGGGACCAGAGCAGT; reverse: CTGCTTGCGCTTCTCGTTGA
Actin: Forward: TTGCTGACAGGATGCAGAAG; reverse: CCACCGATCCACACAGAGTA
Gapdh: Forward: TGCACCACCAACTGCTTAG; reverse: GGATGCAGGGATGATGTTT
FGF4: Forward: GGGTGTGGTGAGCATCTTCGGA; reverse: GGTATGCGTAGGACTCGTAGGGC
Cbx7: Forward: TCTCAGGGCAGTCCTTGTCT; reverse: CGTCCTCAGGTAGGAAGCTG
Ring1a: Forward: CCTGGACATGCTGAAGAACA; reverse: TCCCGGCTAGGGTAGATTTT
Eed: Forward: CTGGCAAAATGGAGGATGAT; reverse: GGTCAGTGTTGTGCATTTGG
Bmp4: Forward: ACAATGTGACACGGTGGGA-AAC; reverse: TGTGGGTGATGCTTGGGACTAC
Socs3: Forward: ATTTCGCTTCGGGACTAGC; reverse: AACTTGCTGTGGGTGACCAT
shBrg1 construct: Open Biosystems PLKO.1 vector: TRCN0000071386; hairpin sequence: CCGGCG-CCCGACACATTATTGAGAACTCGAGTTCTCAATAATGTGTCGGGCGTTTTTG targeted to CGCCCGACACATTATTGAGAA
Results
Selection of a Reporter Gene for esBAF Activity
Using ChIP-seq and microarray data from a conditional Brg1 knockout ES cell line, our lab has determined that the esBAF complex is enriched both at highly expressed ES cell-specific genes and at repressed developmental or lineage-specific genes.1,21 A major group of genes directly repressed by esBAF are members of the Polycomb repressive complex, including Bmi1, Cbx7, Eed, Ring1a, Phc1, Phc2, and Suz12. Of these, we chose to further validate Bmi1 as a reporter of esBAF activity. Bmi1 is a particularly interesting target due to its essential role in the maintenance and self-renewal of hematopoietic and neural stem cells, as well as its role as an oncogene. 22 Thus, although our primary focus is to identify inhibitors of the BAF complex, compounds that regulate the expression of Bmi1 independent of the BAF complex will also be of interest and will be pursued separately. Using qRT-PCR, we observed a 10-fold increase of Bmi1 expression upon tamoxifen-induced Cre-mediated Brg1 deletion from a conditional knockout ES cells line, Brg1f/f, actin-CreER ( Fig. 2A ). 21 There was no delay between the decrease in Brg1 and the subsequent increase in Bmi1, providing additional evidence that Bmi1 is a direct target of the esBAF complex ( Fig. 2B ). We confirmed the regulation of Bmi1 transcription in ES cells by the BAF complex by testing a short-hairpin RNA (shRNA) against Brg1 in E14 ES cells, a commonly used feeder-free ES cell line ( Fig. 2C ). Knockdown of Brg1 for 72 h displays a similar robust increase in the transcript of Bmi1 as the Brg1 knockout (KO) cell line ( Fig. 2D ). Taken together, these data validate Bmi1 as a reporter for esBAF activity.
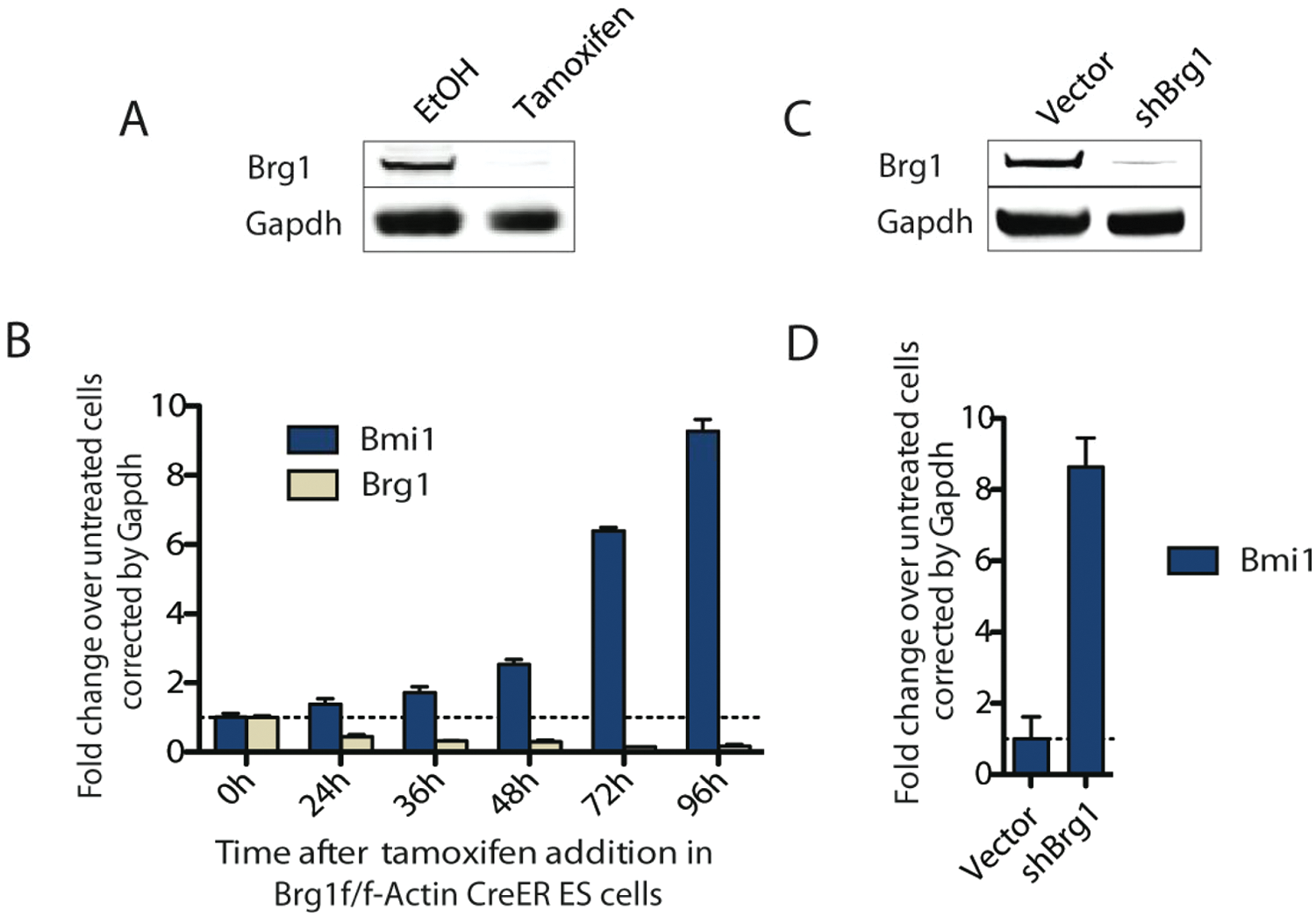
Depletion of BRG1 results in increased Bmi1 expression. (
Development and Validation of a Bmi1 Expression-Based Assay
We developed a robust qRT-PCR expression-based assay to measure Bmi1 transcript levels in mouse ES cells as a reporter of esBAF-mediated repression. Using mouse E14 cells and Ambion’s Cells-to-Ct system with Applied Biosystem’s Taqman probes, we optimized individual assay parameters (cell number, incubation period prior to adding compound, incubation period with compound, qPCR master mix type, housekeeping gene selection, and Taqman probe concentration) for successful assay execution in a 384-well format. Since esBAF is necessary for ES cell viability and pluripotency, esBAF inhibitors may cause a decrease in cell number that occludes any increase in Bmi1 expression. Therefore, to correct for cell number and compound toxicity, we felt it important to multiplex Bmi1 and actin expression using compatible Taqman probes. We screened a number of housekeeping genes with cDNA from Brg1f/f and actin-CreER ES cells, including GAPDH, actin, and HSP90. We found that all the housekeeping genes tested gave very similar results (
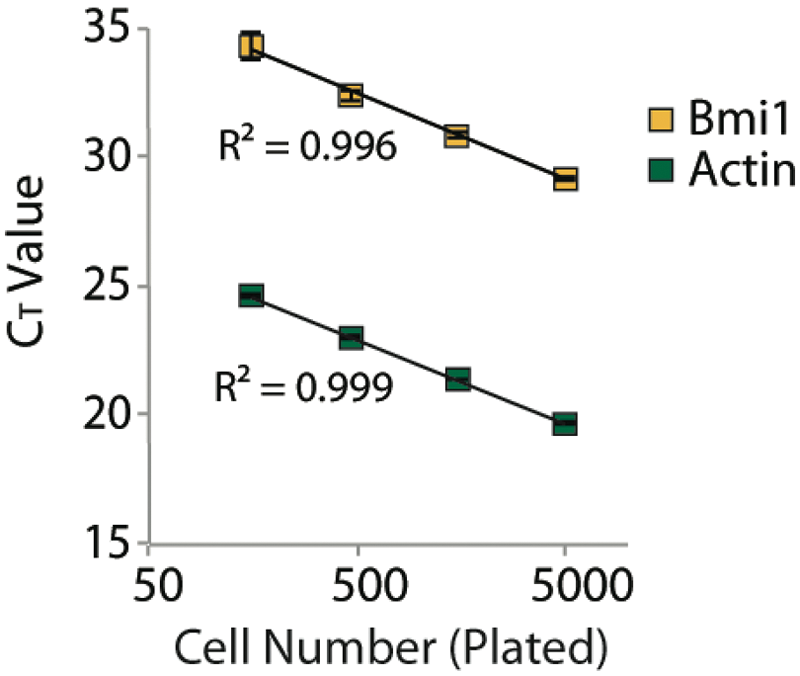
CT value as a function of number of cells plated per well of a 384-well plate. The direct relationship between Bmi1 and actin CT values means that the ΔΔCT value provides an accurate normalization of Bmi1 transcript levels by actin, even at low densities of cells.
High-Throughput qRT-PCR Screen and Validation of Hits
Having demonstrated that the qRT-PCR assay was robust and capable of detecting hits, we initiated a screen of an additional 28 000 compounds (see
As an orthogonal approach to assessing esBAF activity, we developed a mouse ES cell line with a luciferase reporter knocked into the Bmi1 locus. 18 We found a reproducible induction of luciferase upon RNAi-based depletion of Brg1, when corrected for cell number ( Fig. 5 ). Since the esBAF complex is necessary for both pluripotency and self-renewal in ES cells, depletion of Brg1 by RNAi causes a decrease in cell proliferation. Similarly, inhibitors of esBAF would likely also cause a reduction in cell viability that would confound the ability to monitor an increase in Bmi1 expression. Therefore, we desired a way to correct for cell number. We created an internal control in the knock-in cell line by infecting it with a lentivirus containing the renilla luciferase gene under the EF-1a promoter. Luciferase from Renilla reniformis uses a different substrate than firefly luciferase, allowing for multiplexing of readouts using commercially available kits. We optimized and validated the ES reporter cell line using Promega’s Dual-Glo luciferase reagent and tested all 82 “cherry-picked” hits. Forty-three compounds showed dose-dependent behavior in the secondary luciferase reporter screen (a representative concentration-response curve is shown in Fig. 4C ). Comparison of the performance of hits in both the primary qRT-PCR assay and the secondary luciferase reporter assay identified 34 compounds that showed dose-dependent responses in both assays. Compounds determined to be hits in the qPCR screen but not the luciferase screen were not followed up on due to the possibility that they may only appear to be hits by downregulating actin transcript levels instead of upregulating Bmi1 levels. Many of the hits show a decrease in actin levels at high concentrations due to a loss of cell viability (as we would expect for esBAF inhibitors) that would be indistinguishable from a compound that interferes with actin transcription. Thus, the luciferase secondary screen was important for eliminating these potential artifacts. The high overlap between the two complementary screens (80%–90%) validates our approach for identifying compounds that upregulate Bmi1 expression in ES cells.
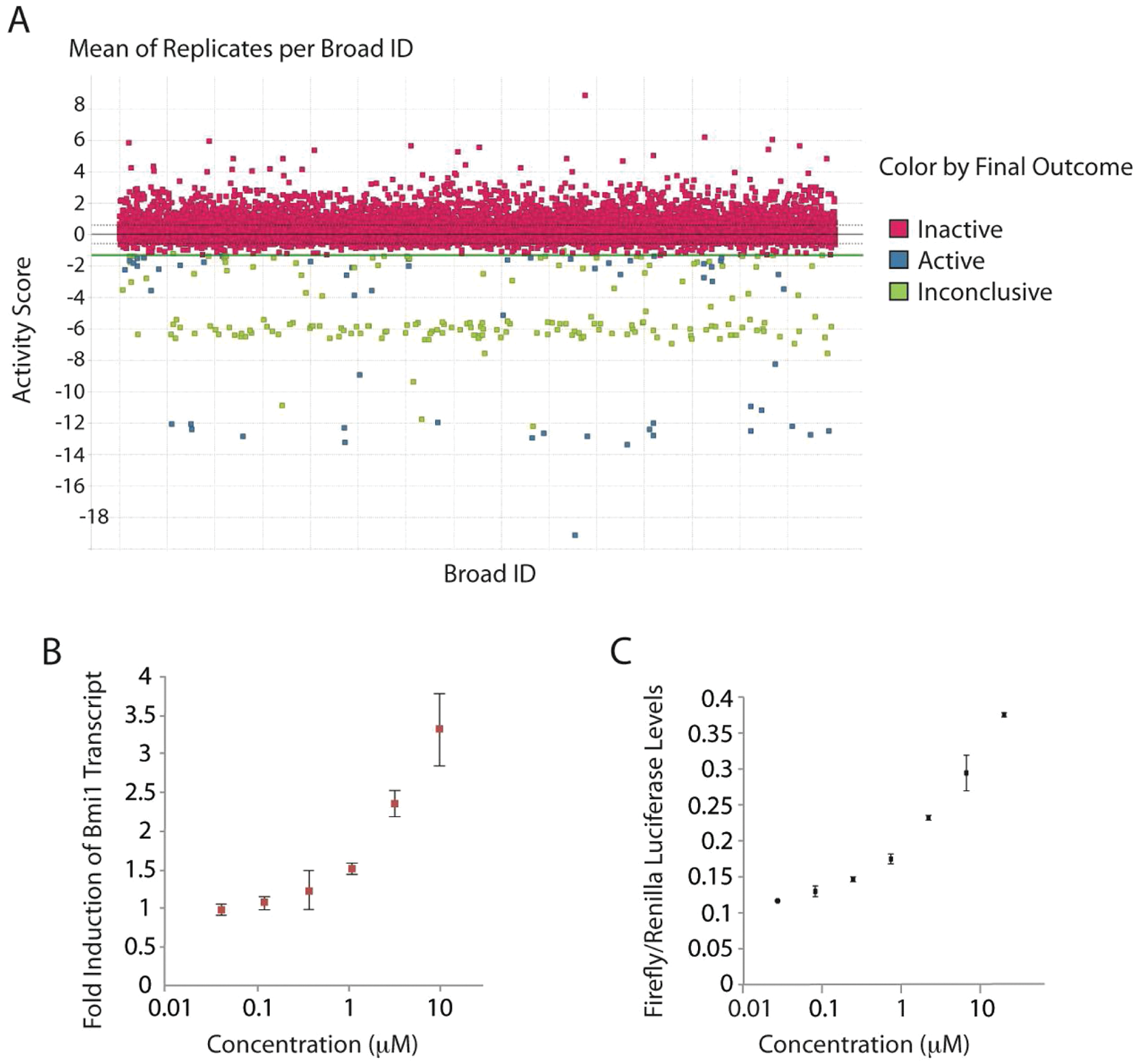
(
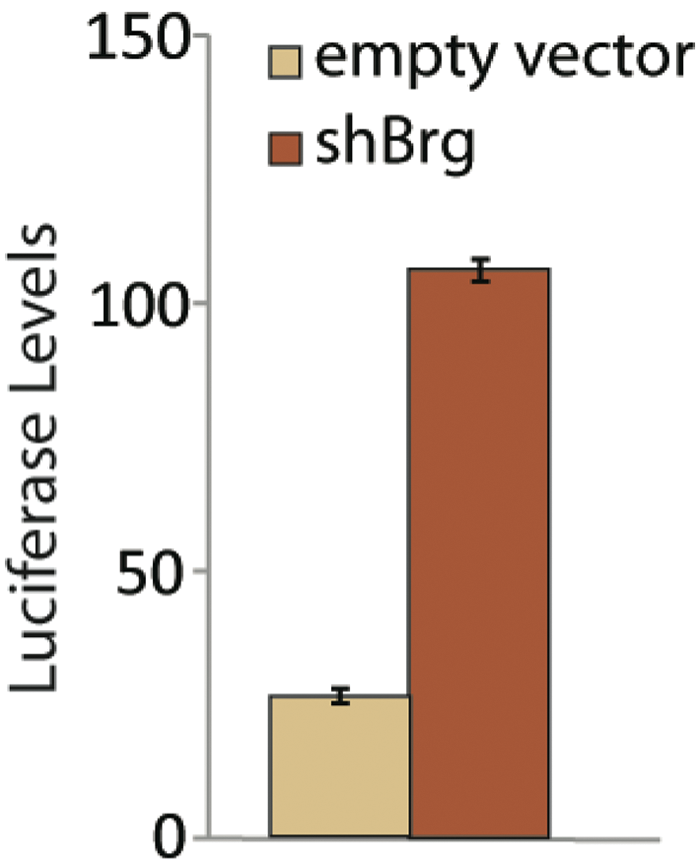
Increase in Bmi1 levels upon BRG1 depletion as indicated by the luciferase reporter. Data are shown at 72 h after viral infection with shRNA against Brg1.
In addition to confirming a compound’s ability to induce Bmi1 transcription, we used the cDNA obtained during the rescreen of hits to perform qRT-PCR for additional developmentally important esBAF targets identified from microarray and ChIP-seq experiments. The ability to resample the cDNA samples obtained during the rescreen of hits for multiple relevant targets is an advantage of the qRT-PCR approach. We initially used SYBR Green to examine the effects of hits on the transcriptional regulation of Phox2b and Fgf4, in addition to Bmi1. 1 To rule out any potential artifacts obtained from using actin as a housekeeping gene, we tested both actin and GAPDH as housekeeping genes in SYBR Green studies. We found similar results with both housekeeping genes. For all SYBR Green primers tested, melting curves were generated and the products were validated with sequencing. This first screen of Bmi1, Phox2b, and FGF4 levels served to eliminate compounds that may have general repressive activities or may be acting on a separate target or pathway responsible for Bmi1 regulation. Of the 34 confirmed and validated hits tested, 22 compounds regulated these targets in a manner similar to esBAF ( Fig. 6 ). Following analysis of mRNA expression levels of five additional esBAF transcriptional targets (Eed, Cbx7, Ring1a, Bmp4, and Socs3), another 2 compounds were eliminated, leaving 20 compounds for follow-up studies focusing on determining the mechanism of action. These studies are currently under way.
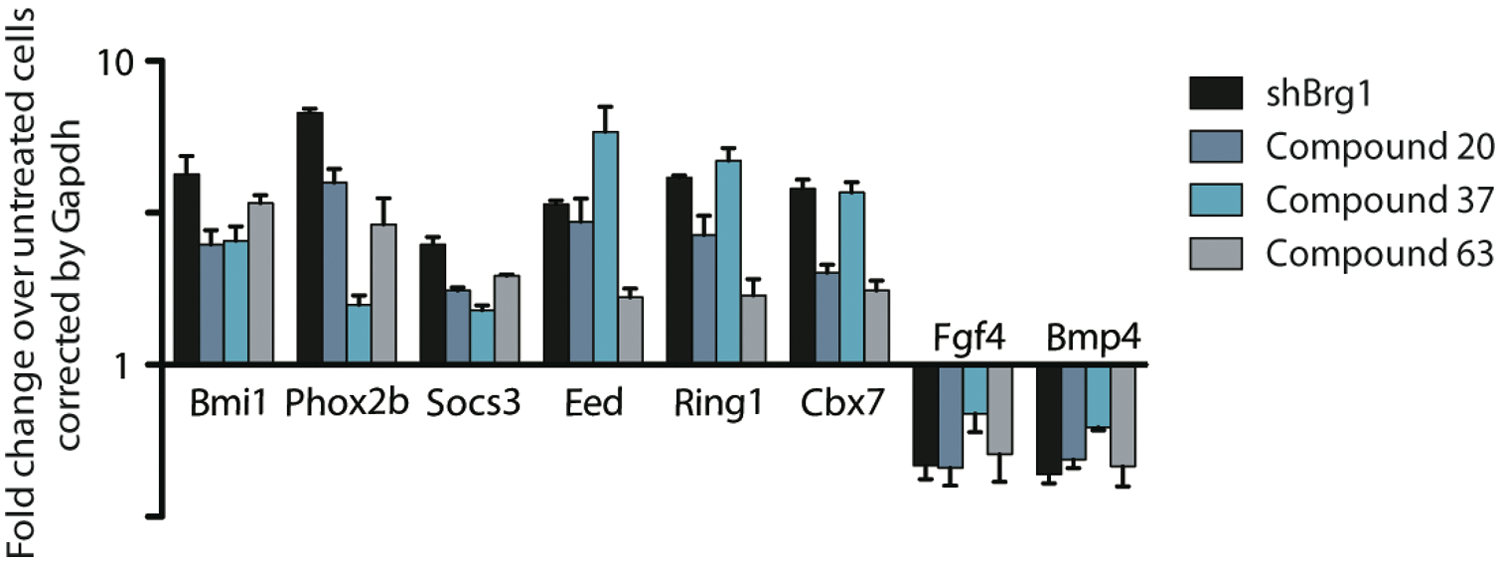
Transcriptional profiles of E14 ES cells treated with shBrg1 (Brg1 KD) and three representative hits. The fold change was calculated as comparison to DMSO-treated embryonic stem (ES) cells using Gapdh as the housekeeping gene. Twenty compounds out of ~30 000 screened produced a transcriptional profile similar to Brg1-deficient cells.
Discussion
To identify compounds that inhibit the esBAF complex, we developed a gene expression–based screening system using mouse ES cells. By monitoring mRNA expression levels of Bmi1, a developmentally important esBAF-repressed target gene, we were able to identify a series of compounds that mimic the transcriptional action of the BAF complex in mouse ES cells. Measurement of the endogenous Bmi1 mRNA expression levels by qRT-PCR afforded several advantages as a primary screening strategy. It provided a straightforward way to screen for transcriptional activity in ES cells without genetic manipulation, thus reducing the risk of assay artifacts. In addition, we found there to be minimal optimization required to produce an assay with low variability and high reproducibility. Both of these points are concerns with standard reporter gene assays (e.g., luciferase, β-galactosidase reporters). Although there are limits on the number of compounds that can be screened with this method (primarily due to cost and throughput factors), we have determined that gene expression–based screening is a physiologically relevant, robust method for screening a medium-sized library of compounds in ES cells.
Although unconventional, we found this screening method to be an advantageous way to screen for inhibitors of novel transcriptional regulators for which very little is known mechanistically and for which there are no known inhibitors. We were able to use the qRT-PCR screen to validate the target and to identify a panel of potential esBAF inhibitors for follow-up studies. The qRT-PCR screen was very robust and reproducible, making it fairly quick to identify and confirm hits with a minimal number of false positives. Furthermore, storage of the cDNA isolated from compound-treated cells enabled rapid interrogation of additional targets to validate the biological relevance of confirmed hits. From this work, we identified a panel of 20 compounds that transcriptionally mimic the Brg1 knockout phenotype in mouse ES cells. Several factors, including cost, throughput, and ES cell-specific considerations, will likely make a qRT-PCR approach less feasible when screening larger collections (hundreds of thousands to millions) of compounds. Efforts are currently under way to miniaturize the assay into a 1536-well format to expand the feasibility to larger screens.
What still remains to determine is the mechanism of action for these validated hit compounds. As we still do not understand the mechanism of BAF-mediated repression, this remains a significant challenge. Our first step will be to identify compounds that bind directly to the esBAF complex using mass spectrometry–based binding assays. 23 BAF is known to catalyze the movement and displacement of nucleosomes. Since the ATPase function of BRG1 is necessary for this function, our next step will be to characterize any BAF binders in an in vitro ATPase assay and an in vitro nucleosome remodeling assay.13,14 For compounds that bind to esBAF but do not inhibit remodeling activity in vitro, we will determine the subunit targeted by the small molecules using seven established embryonic stem cell lines that have been created in our lab with null mutations in subunits of the complex. Finding inhibitors that target certain domains, such as plant homeodomain fingers, chromodomains, or bromodomains, will help elucidate the roles of individual subunits in esBAF targeting and may explain how different combinations of BAF subunits promote cell-specific transcriptional programs.24,25 We will investigate how these compounds affect the known in vivo effects of the BAF complex. As mentioned earlier, only a few subunits are required for nucleosome remodeling activity in vitro, whereas many other subunits are required for the in vivo activities of BAF. We will investigate the effect of compounds on the activities, which include but are not limited to nucleosome positioning, 26 changes in histone variants and histone modifications, DNase I hypersensitivity sites associated with open chromatin in the promoter regions of actively transcribed genes,15,27 looping and higher-order chromatin structure,28,29 transcription factor occupancy, 1 and the cellular effects related to loss of pluripotency.
Using a transcriptional reporter of esBAF repression not only allows us to identify compounds that target the esBAF complex directly but will also enable us to identify small-molecule inhibitors of proteins that act on or with the BAF complex. Future work will focus on determining if the validated hits inhibit players in putative ES cell-specific signaling pathways that regulate esBAF function or inhibit proteins that act in concert with the esBAF complex, including transcription factors and chromatin-modifying enzymes required for esBAF binding or activity. For compounds that do not bind directly to the complex, we plan to use established assays for target identification, 30 with the goal of identifying novel protein partners required for BAF activity in ES cells.
Small-molecule inhibitors will be used as tools to provide the precision and temporal control needed to tease out the precise functions and mechanisms of the BAF complex. These compounds will prove useful to researchers studying stem cells, chromatin, and cancer. These compounds may even one day be used for a therapeutic function, as an increase in Brg1 expression in adult cardiomyocytes has recently been shown to be a major player in hypertrophy. 31
Footnotes
Acknowledgements
We thank Stuart L. Schreiber for his generous support and guidance (GM38627, S.L.S.), Naoki Hosen for his generous gift of the Bmi1-GFP construct, P. J. Aspesi and Vihren Kolev for technical advice, and Lili Wang and Josh Bittker for critical reading of the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institute of General Medical Sciences (GM38627), the National Cancer Institute (N01-CO-12400), the National Institute of Neurological Disorders and Stroke (NS046789), and the American Cancer Society (121535-PF-11-145-01-DMC). The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Service, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. government.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.