
Abstract
Activation of the enzyme indoleamine 2,3-dioxygenase 1 (IDO1) underlies the course of several human pathological conditions and, to date, no efficacious therapeutic IDO inhibitors are available. We proposed to develop a robust screening system based on the use of yeast cells to identify new lead compounds for the pharmacological inhibition of IDO—the BLOCKADE platform. Yeast combines the advantages of a relevant surrogate model for eukaryotic cell processes with the amenity to miniaturization and automation. We brought added value to the system by increasing the stringency of our assay, as the BLOCKADE strain was not deleted for any efflux pump, thus creating additional challenges for test compounds to be identified as hits. Screening of a library of 50 080 small molecules led to the identification of 101 potential IDO inhibitors, a low hit rate of 0.2%, reflecting the stringent assay conditions imposed. Most important, secondary pharmacology assays in mammalian cells confirmed activity for 76% of the hits, whereas hepatotoxicity testing indicated that 87% of them displayed a safe profile. The high predictivity rates obtained using the BLOCKADE platform clearly validate our system as a powerful tool for drug discovery.
Introduction
Indoleamine 2,3-dioxygenase 1 (IDO1) is an intracellular heme-containing enzyme that catalyzes the first and rate-limiting step of the kynurenine pathway, through which 95% of the nondietary tryptophan (trp) is metabolized. 1 Expression of IDO is upregulated by proinflammatory cytokines and can be detected in a variety of tissues, including placenta, spleen, thymus, lung, digestive tract, and central nervous system. 2 Alterations in IDO activity have been implicated in a number of human pathologies, due to the reduction of local trp levels and/or to imbalances in the level of cytotoxic metabolites produced by the kynurenine pathway.3,4 Based on the evidence for a relevant role of IDO in human pathology, inhibition of IDO activity as a therapeutic strategy has been tested in preclinical models of many diseases. The most widely used IDO inhibitor is the tryptophan analogue 1-methyltryptophan (1MT). 5 Treatment with 1MT, alone or in combination with other agents, attenuated disease severity in animal models of arthritis, 6 ischemia-reperfusion injury, 7 endotoxin shock, 8 human immunodeficiency virus (HIV)/simian immunodeficiency virus (SIV) infection, 9 airway inflammation, 10 and cancer, 11 among others. Data obtained from these preclinical studies resulted in strong evidence corroborating the use of pharmacological IDO inhibitors in human patients. Supporting this approach is the fact that IDO-deficient mice are healthy and develop and reproduce normally, 12 indicating that no major adverse physiological effects are expected to result from the inhibition of IDO activity.
1MT is currently being tested in phase I/II clinical trials for the treatment of relapsed or refractory solid tumors (ClinicalTrials.gov identifier: NCT00739609; NCT00617422). Despite presenting good pharmacokinetics and toxicity profiles, 13 1MT is a weak competitive inhibitor of IDO. The majority of the remaining known IDO inhibitors, either competitive or noncompetitive, are also trp analogues that retain the indole ring and are only active at high concentrations. In addition, some natural products derived from different sources were shown to inhibit IDO and to belong to distinct structural classes. 5 Clearly, the development of new lead compounds with increased efficacy is critical to potentiate the success of the pharmacological inhibition of IDO as a therapeutic approach.
Identification of IDO inhibitors so far has been mostly performed by enzyme activity assays using recombinant human IDO expressed in Escherichia coli. 14 Although this is a well-established, fast, and inexpensive assay, the current lack of strong drug candidates for IDO inhibition strongly supports the requirement of more adequate screening systems. In the current work, we developed a yeast-based screening platform for the identification of IDO inhibitors. The advantages of using yeast as a model organism for the study of cellular alterations underlying human diseases are widely accepted and have been proven by major breakthrough discoveries, particularly in the field of neurodegenerative diseases. 15 In the yeast Saccharomyces cerevisiae, tryptophan metabolism through the kynurenine pathway leads to the biosynthesis of NAD+ ( Fig. 1 ). 16 A second pathway, using nicotinate phosphoribosyltransferase (NTP1), also allows synthesis of NAD+, and it has been shown that deletion of genes encoding kynurenine pathway enzymes is co-lethal with deletion of NPT1. 16 The yeast homolog of human IDO is the protein encoded by the BNA2 gene. 16 The goal of our study was to obtain a genetically modified yeast strain whose survival was dependent on the activity of human IDO. Co-deletion of BNA2 and NPT1 abolishes the synthesis of NAD+ and therefore is lethal for the yeast cell. 16 We proved that heterologous expression of human IDO in yeast is capable of complementing this double deletion, thus reverting the lethal phenotype. We then developed a stringent screening system for the identification of human IDO inhibitors—the BLOCKADE yeast platform—which was used for the high-throughput screening (HTS) of a small-molecule library of 50 080 compounds, allowing the identification of 101 hits with potential for further development. Secondary potency and toxicity testing in mammalian cell systems corroborated the predictivity of our yeast system, resulting in residual attrition levels, validating our technology as a powerful tool for the early stages of drug discovery and bioactive identification.
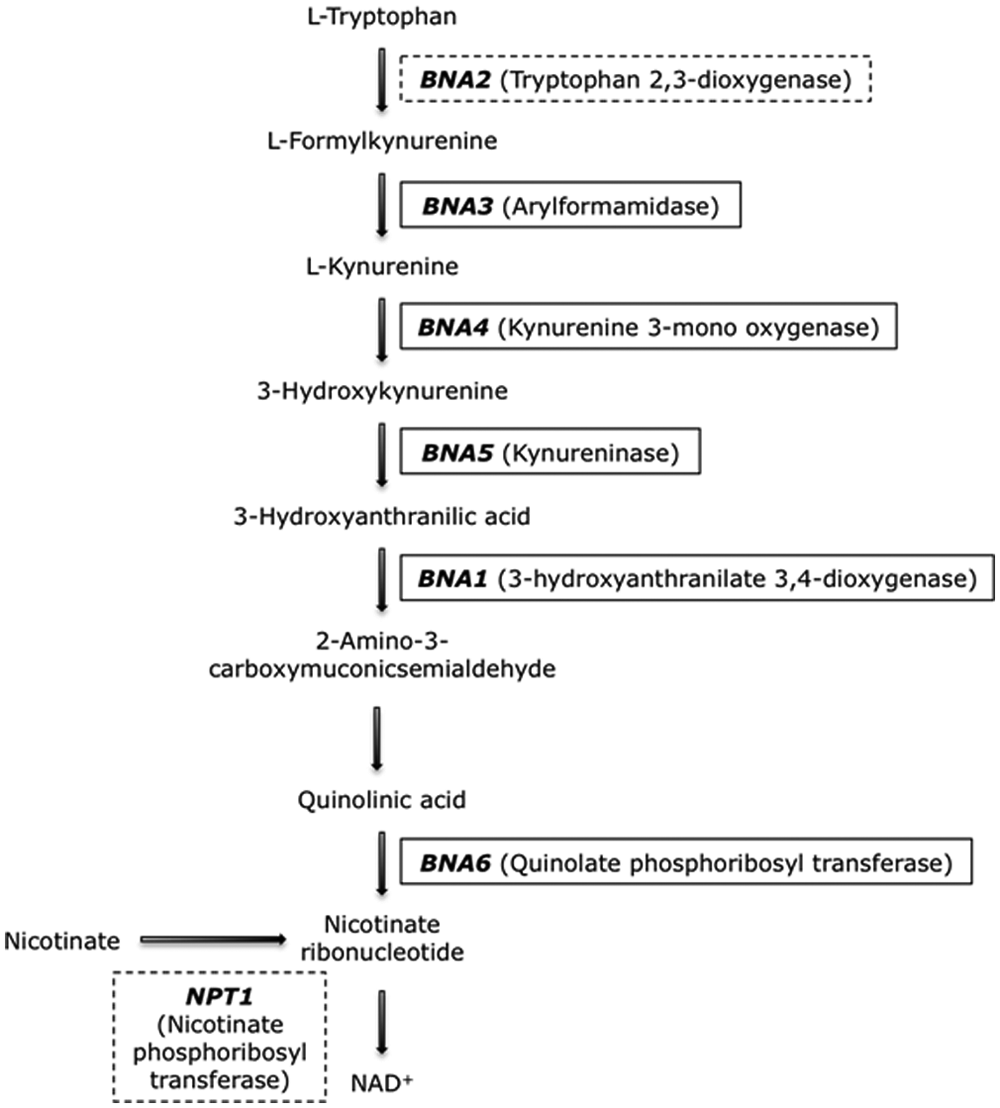
NAD+ synthesis pathways in Saccharomyces cerevisiae cells. If both NPT1 and BNA2 are deleted (dashed squares), NAD+ synthesis stops and the cell cannot survive.
Materials and Methods
Plasmids and Cloning
Taq polymerase and T4 DNA ligase were purchased from Invitrogen (Carlsbad, CA), and the restriction enzymes were purchased from New England Biolabs (Ipswich, MA). The QIAquick Gel Extraction Kit and QIAprep Spin Miniprep Kit, from Qiagen (Valencia, CA), were used to purify DNAs for agarose gel and to purify high molecular grade DNA, respectively. The human IDO coding sequence was purchased from the I.M.A.G.E. Consortium and cloned in the yeast expression episomal plasmid pESC-LEU in the restriction sites SmaI and XhoI, under the control of the GAL1 promoter. For expression in mammalian cells, human IDO coding sequence was cloned in pCDNA3 using the restriction sites EcoRI and XhoI, under the control of the cytomegalovirus (CMV) promoter. Sequence integrity of IDO was confirmed by sequencing.
Yeast Strains and Media
The yeast strains used in this study derive from S. cerevisiae BY4741 strain (MATa; his3Δ1; leu2Δ0; met15Δ0; ura3Δ0). Yeast cells were grown in Yeast Nitrogen Base (YNB) supplemented with uracil and/or leucine. Transfor-mations were performed using the lithium acetate method. 17 A 1500-bp DNA fragment containing 500 bp homology with the BNA2 promoter at the 5′ end and 200 bp homology with the BNA2 terminator sequence at the 3′ end was transformed in the BY4741 strain to promote homologous recombination in the promoter and terminator regions of the BNA2 gene and therefore remove the respective coding sequence. Positive colonies for BNA2 deletion were selected by the PCR colony using the forward primer 5′-ACTCCGCAATTCGTCGTAAAC-3′, located in the BNA2 promoter, and the reverse primer 5′-TTATAATATCT-TTCTGGCCTC-3′, located in the BNA2 terminator (amplicon size 1500 bp). A second confirmation PCR colony was performed to confirm the presence of the DNA fragment used for gene replacement (300 bp). The forward primer used was 5′-GACCCTGAAGTTCATCTGCAC-3′, and the reverse primer was 5′-TAGTTGTACTCCAGCTTGTGC-3′. pESC-LEU-IDO plasmid was then transformed in the BNA2-deleted BY4741 strain. The yeast integrative plasmid YIplac211 containing a 440-bp sequence from the NPT1 promoter followed by a 200-bp sequence from the NPT1 terminator was linearized inside the NPT1 promoter sequence and transformed in the BNA2-deleted/pESC-LEU-IDO BY4741 strain. Integration in the NPT1 promoter locus gave rise to URA3+ cells. Transformants were selected on galactose medium to induce IDO expression and maintain cells alive, as co-deletion of BNA2 and NPT1 is lethal. Treatment of transformants with 5-Fluoroorotic Acid (5-FOA) induced the production of the toxic compound 5-fluorouracil, leading to the expelling (pop-out) of the NPT1 gene by homologous recombination between the regions of homology flanking the URA3 gene 18 and generating the strain designated as BLOCKADE. Both gene deletions were confirmed by PCR. Growth of BLOCKADE strain was assessed on solid media by spot assay with 10-fold serial dilutions of exponentially growing cultures. Samples from each dilution, starting from OD600 = 1, were spotted on selective medium supplemented with 2% galactose or 2% glucose, in triplicate independent experiments.
Yeast Protein Extraction and Western Blotting
Yeast cells were preinoculated on minimum selective media, containing 1.2% glucose and 0.8% galactose, at 30 °C, 200 rpm. After overnight growth, inocula at a starting OD600 of 0.2 were performed in minimum selective media containing 2% galactose and grown until an OD600 of 1. Cells were collected in Y-PER Reagent (Thermo Scientific, Rockford, IL) containing 1× Protease Inhibitor Cocktail (Sigma, St. Louis, MO), according to the manufacturer’s protocol. Samples were diluted with 5× SDS Sample Buffer (60 mM Tris-HCl [pH 6.8], 25% glycerol, 2% sodium dodecyl sulfate [SDS], 0.715 M β-mercaptoethanol, and 0.1% bromophenol blue) followed by incubation in a boiling water bath for 5 min. Equal amounts of protein from each sample were applied on 15% SDS–polyacrylamide electrophoresis gel, transferred to nitrocellulose membrane, and probed with anti-IDO antibody (Chemicon, Temecula, CA) with dilution 1:1000, ON at 4 °C. After washing with Tris-buffered saline with Tween-20 (TBST) 1×, membrane was incubated with monoclonal horseradish peroxidase–conjugated secondary antibody (Bio-Rad, Hercules, CA) with dilution 1:3000 during 45 min. After washing with TBST 1×, the blot was developed using ECL (Amersham Biosciences, GE Healthcare, Little Chalfont Buckinghamshire, England).
Screening Assay and Data Analysis
The DIVERSet compound library (ChemBridge, San Diego, CA) was diluted in DMSO and plated in 96-well plates at a concentration of 10 mM. For screening assays, dilution plates at a final concentration of 1 mM were stored at –20 °C. All liquid-handling steps were performed using a Janus Automated Workstation (PerkinElmer, Waltham, MA). Yeast cells were grown in a Liconic STX40 Automated Incubator, and growth was monitored with a Victor 3V microplate reader (PerkinElmer). Prior to the screening campaign, a frozen batch of BLOCKADE strain was prepared in 1-mL aliquots. For each assay, one aliquot of cells was thawed and pregrown on a mixture of 1.2% glucose and 0.8% galactose until reaching early exponential phase (OD = 1–2). Cells were then washed three times with water and resuspended in fresh selective medium containing 2% galactose to fully activate IDO expression. Cells were dispensed into 96-well plates, and test compounds were added to each well at a final concentration of 10 µM in 2% DMSO. Yeast growth was monitored during 3 days at 30 °C with agitation. The plates were formatted to contain 16 blanks (nontreated BLOCKADE strain) on the outside columns of the 96-well plate. Hits were identified as compounds that inhibited BLOCKADE growth (measurement of final OD) >60%. Hit confirmation was performed in a secondary screening with decreasing concentrations of compound (10, 5, 2.5, and 1 µM). Robustness of the assay for screening was assessed by calculating the Z′ factor. 19 Chemical clustering of hits was performed using Pipeline Pilot Student Edition software (Accelrys, San Diego, CA).
Pharmacology Assay in COS-7 Cells
COS-7 cells were grown in DMEM 1000 mg/mL glucose, with GlutaMAX and pyruvate (Invitrogen, Carlsbad, CA), supplemented with 10% fetal bovine serum (FBS) and 1% nonessential amino acids (NEAAs). Cells were maintained at 80% to 90% confluence at 37 °C and 5% CO2. COS-7 cells in 24-well plates were transiently transfected with pcDNA3-IDO using FuGene HD (Roche Diagnostics, Basel, Switzerland) following the manufacturer’s instructions. Three hours posttransfection, 100, 10, 1, and 0.1 µM of each compound was added to cells and left for 24 h. Transfection efficiency after 24-h compound exposure was assessed by direct observation of the enhanced green fluorescent protein (EGFP) signal, using an inverted Carl Zeiss microscope (Oberkochen, Germany) AxioObserver D1 (excitation = 485/20 nm, emission = 515 nm). Moreover, immunofluorescence in untreated cells transfected with pcDNA3-IDO was performed using the mouse anti-IDO monoclonal antibody at a dilution of 1:100 (Chemicon) detected with Cy3-conjugated AffiniPure F(ab′)2 Fragment Goat Anti-Mouse IgG (H+L) (Jackson ImmunoResearch Laboratories, West Grove, PA) at a dilution of 1:300, according to standard procedures. Observation was made using an inverted Carl Zeiss microscope AxioObserver D1 (excitation = 470/40 nm, emission = 525/50 nm).
COS-7 Protein Extraction and Western Blotting
COS-7 were grown until 80% to 90% confluence and washed twice with phosphate-buffered saline (PBS). Cells were collected directly in 1× sample buffer with 1× benzonase (Sigma) and 1× Protease Inhibitor Cocktail (Sigma). Then, 60 µg of each sample was boiled for 5 min and applied on 15% SDS–polyacrylamide electrophoresis gel, transferred to a nitrocellulose membrane, and incubated with anti-IDO antibody (Chemicon) at 1:1000 dilution. After washing, the membrane was incubated with monoclonal horseradish peroxidase–conjugated secondary antibody (Bio-Rad) at 1:3000 dilution, and detection was performed with ECL (Amersham Biosciences).
IDO Activity Assay
IDO activity was determined by measuring kynurenine concentration in the supernatant by high-performance liquid chromatography (HPLC), as described previously. 20 Briefly, supernatants from cell culture were collected and immediately frozen at –20 °C until analysis. Protein precipitation and kynurenine extraction were performed by addition of trichloroacetic acid (TCA) at a final concentration of 6%. After discarding cell debris by centrifugation, supernatants were injected into the HPLC pump (Model LC-6A; Shimadzu Corporation, Nakagyo-ku, Kyoto, Japan). Separation was performed using a reversed-phase cartridge Aquasil RP18 column (200 mm length, 4.6 µm grain size) from Thermo Scientific. A SPD-6AU UV-VIS spectrophotometric detector (Shimadzu Corporation) in flow stream series connection was used for detection of kynurenine at a wavelength of 360 nm. The elution buffer consisted of a degassed potassium phosphate solution (0.015 mol/L, pH 6.4) containing 27 mL/L acetonitrile. Analysis was carried out at room temperature at a flow rate of 1.2 mL/min.
The half-maximal effective concentration (EC50) was calculated for each compound, related to the maximum kynurenine production observed in each transfection assay, by linear regression. The EC50 values for each compound were later corrected, taking into account the early cytotoxicity data obtained, as the EC50 value of each compound is affected by the number of viable cells present in each well. Compounds were ranked according to their potency in IDO inhibition as weak (EC50 > 100 µM), average (100 µM > EC50 > 10 µM), good (10 µM > EC50 > 1 µM), and very good (1 µM > EC50 > 0 µM).
Cytotoxicity Assay in HuH-7 Cells
HuH-7 cells were grown in RPMI 1640 (Invitrogen) supplemented with 10% FBS and 1% NEAA. Cells were maintained at 80% to 90% confluence at 37 °C and 5% CO2. For the cytotoxicity assays, aliquots from a common batch were thawed and seeded at 14 × 104 cells/well in 96-well plates. Test compounds were added at eight concentrations (125, 100, 75, 50, 25, 10, 1, and 0.1 µM) in triplicate and incubated for 2 h or 24 h. LDH activity (Cytotoxicity Detection Kitplus [LDH]; Roche Diagnostics) was assayed by measuring absorbance at 492 nm with background subtraction at 690 nm and ATP production (ATPlite 1step Assay System; PerkinElmer) by measuring luminescence, according to the manufacturer’s instructions. The microplate reader INFINITE M200 (Tecan Group, Männedorf, Switzerland) was used for measurements and the software Magellan v6.0 (Tecan Group, Männedorf, Switzerland) for data treatment. Dose-response curves of chlorpromazine (CPZ), a reference compound for cytotoxicity, were used to evaluate the performance of each assay and validate the obtained data.
The half-maximal lethal concentration (LC50) was determined for each time point of the LDH and ATP assays by linear regression (R2 values below 0.970 were not accepted). When less than 50% of cells were dead for the highest compound concentration used, the LC50 was considered higher than 125 µM.
These early in vitro hepatotoxicity assays allowed the ranking of hits as high hazard (LC50 < 10 µM), moderate hazard (50 µM < LC50 > 10 µM), weak hazard (125 µM < LC50 > 50 µM), and low hazard (LC50 > 125 µM) for each time and end point. The results obtained were then combined into one classification per time point, and hits were ranked as follows: high hazard (if one classification at either time point was high hazard), moderate hazard (moderate hazard at both time points or moderate hazard at one time point and weak hazard at the other), weak hazard (weak hazard at both time points or low hazard at one time point and moderate hazard at the other), and low hazard (low hazard at both time points or low hazard at one time point and weak hazard at the other).
Results
BLOCKADE Strain Characterization
The initial goal of the current study was to construct a yeast strain dependent on the activity of human IDO. A BNA2-knockout strain was created by disruption of the BNA2 gene. Next, the coding sequence of human IDO was cloned in a galactose-inducible yeast vector and transformed into the BNA2-knockout strain for functional complementation. Subsequently, disruption of the NPT1 gene was promoted to abolish both pathways leading to the synthesis of NAD+ in yeast. The resulting strain was designated as BLOCKADE. Galactose-inducible expression of human IDO by the BLOCKADE strain was confirmed by Western blot analysis ( Fig. 2A ). Survival of the BLOCKADE strain in culture media containing galactose proved that expression of human IDO is sufficient to compensate for BNA2 and NPT1 deletion, indicating that human IDO is able to substitute for BNA2 and to promote tryptophan degradation and initiate the kynurenine pathway in yeast ( Fig. 2B ). Residual growth in glucose was observed ( Fig. 2B ), most probably due to vestigial amounts of intracellular IDO (not detectable by Western blot) produced during the preinoculation phase, which occurs in medium containing 1.2 GLU/0.8% GAL. Despite the strong expression of IDO in GAL ( Fig. 2A ), NAD+ levels obtained from IDO activity in this double-deleted strain are likely to be lower than the ones normally produced by the combined activity of the kynurenine pathway and NPT1, as suggested by the increased lag phase of BLOCKADE when compared with the wild-type (WT) strain ( Fig. 2C ). DMSO tolerance was assessed and concentrations up to 2.5% had no significant effect on the growth of the BLOCKADE strain (data not shown).
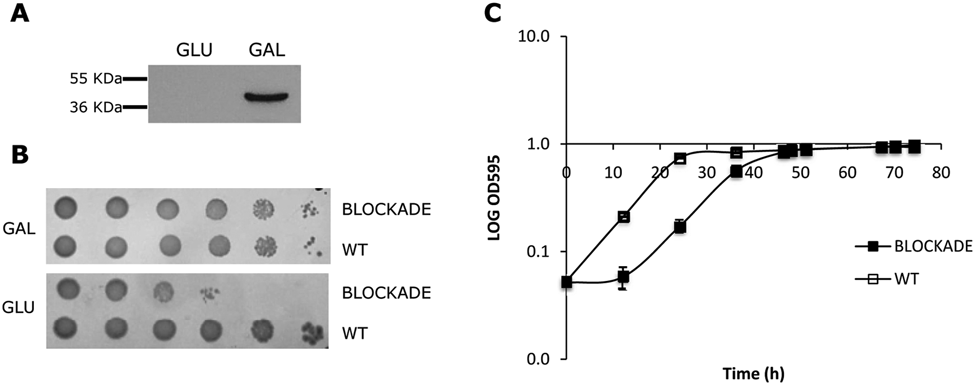
Expression of human indoleamine 2,3-dioxygenase (IDO) in yeast rescued the lethal phenotype caused by BNA2 and NPT1 co-deletion. (
We next tested the effect of known IDO inhibitors on the growth of the BLOCKADE strain as reference compounds for future comparison ( Fig. 3 ). The gold-standard IDO inhibitor is 1MT, whereas 7-methyltryptophan (7MT) and methyl-thiohydantoin-tryptophan (MTH) are two additional trp analogues with known inhibitory effect on IDO activity. 5 To test the effect of 1MT, 7MT, and MTH on the growth of the BLOCKADE strain, cells were preinoculated in 1.2% GLU/0.8% GAL and subsequently inoculated in fresh medium containing 2% galactose to fully activate IDO expression. Cells were dispensed into 96-well plates, and 1MT, 7MT, or MTH was added at final concentrations of 20 µM and 200 µM. Growth at 30 °C under shaking conditions was monitored during 72 h. WT yeast cells were incubated in parallel under identical growth conditions and submitted to the same inhibitor concentrations. An eventual decrease in the growth of the WT strain upon incubation with an IDO inhibitor would indicate a cytotoxic effect of that compound. A similar reduction should also be observed upon incubation of the BLOCKADE strain with the said compound, but that would not reflect specific inhibition of IDO but rather an inhibition of cell growth due to compound toxicity. Comparison of the growth alterations obtained for both the BLOCKADE and the WT strain allows discarding compounds that are cytotoxic, making possible the selection of real IDO inhibitors for further studies.
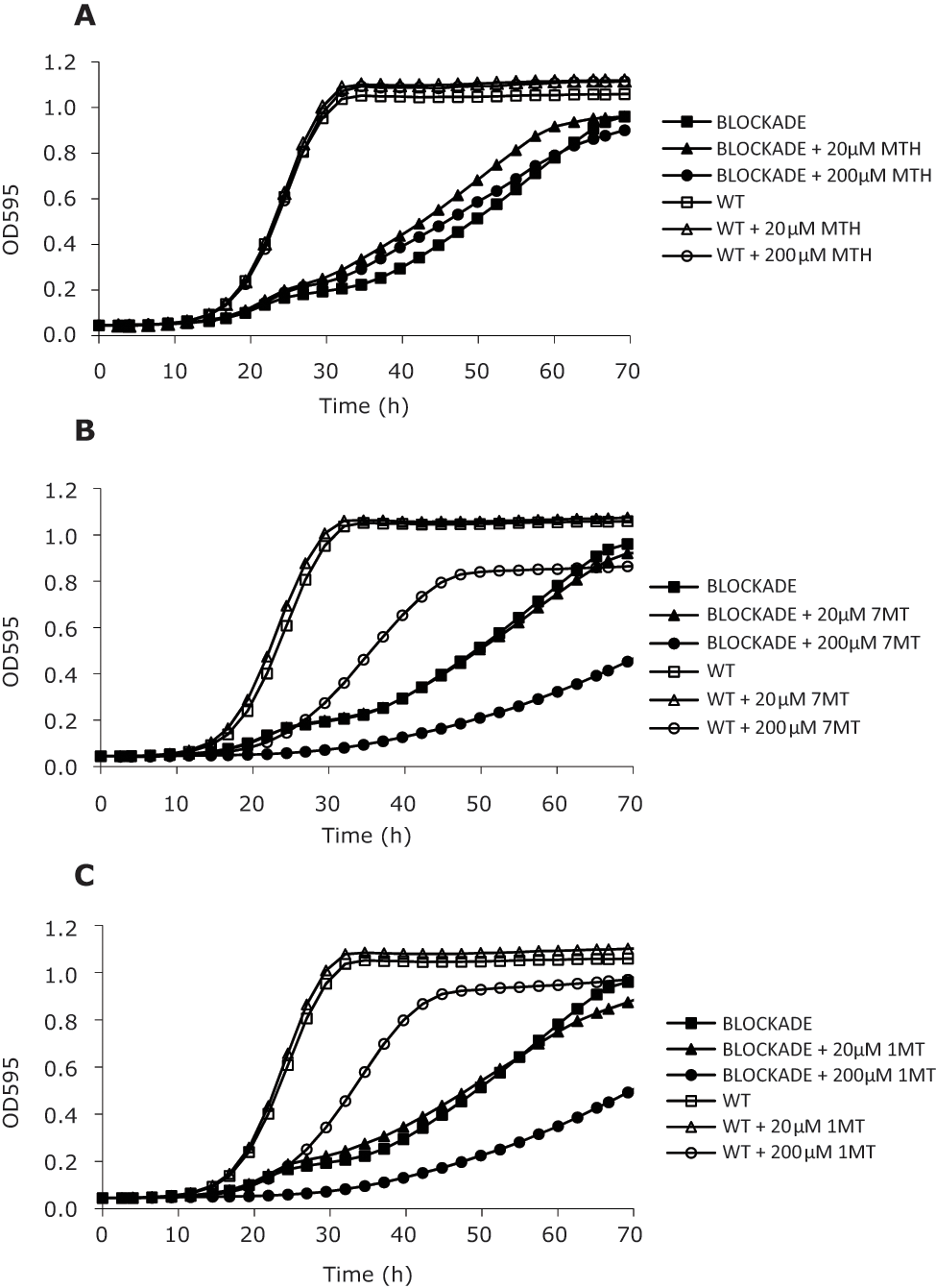
Known indoleamine 2,3-dioxygenase (IDO) inhibitors show a weak effect on the growth of the BLOCKADE strain. BLOCKADE and wild-type (WT) strains were cultured in medium with 2% galactose containing 0, 20, and 200 µM (
Treatment with MTH did not alter the growth profile of the BLOCKADE strain, suggesting that this inhibitor is not potent enough to be active in our yeast system ( Fig. 3A ). In contrast, 7MT and 1MT were able to inhibit the growth of the BLOCKADE strain by 47% and 40%, respectively, but only at very high concentrations (200 µM) ( Fig. 3B,C ). Importantly, treatment of the WT strain with 200 µM of either 7MT or 1MT also resulted in growth inhibition, by 20% and 12%, respectively, indicating a clear influence of these compounds on cell viability. These observations confirmed that the commonly used trp analogues are weak IDO inhibitors and are in accordance with previous results showing a toxic effect of 1MT in yeast. 21 Overall, the results obtained validated the use of the BLOCKADE strain as a stringent assay for the identification of more potent IDO inhibitors, allowing the direct exclusion of compounds with residual inhibitory activity and the identification of cytotoxic liabilities in the earliest stage of compound screening.
Screening Campaign and Hit Identification
The BLOCKADE yeast strain was used for the screening of a small-molecule library of 50 080 compounds (ChemBridge’s DIVERSet). Compounds were tested at an initial concentration of 10 µM. Yeast growth in 96-well plates was monitored during 3 days at 30 °C under agitation, with OD measurements every 2 h. Compounds that inhibited growth of the BLOCKADE strain by more than 60% (measured as a percentage of the final OD) were considered positive. Hits were cherry-picked and confirmed using decreasing compound concentrations (10, 5, 2.5, and 1 µM) to evaluate dose-response correlations and to define potency ranges. Both the BLOCKADE and the WT BY4741 strain were assayed to discard compounds that were false positives or cytotoxic, respectively. Compounds that maintained an inhibitory response at 10 µM, reducing the growth of the BLOCKADE strain by more than 50%, were considered validated hits. Stronger inhibitors induced a 50% reduction in BLOCKADE cell growth at concentrations lower than 10 µM.
Figure 4 displays representative examples of a false-positive compound, a cytotoxic compound, and a hit, as revealed after the secondary screening. As observed, compounds that had no effect on yeast growth were considered false positives ( Fig. 4A ). Compounds that inhibited the growth of both BLOCKADE and WT strains were considered cytotoxic and not selected for further development ( Fig. 4B ). Compounds were considered hits when strong inhibition of IDO activity was observed and a good dose-response correlation pattern was obtained ( Fig. 4C ). Our yeast-based platform was therefore able to identify specific IDO inhibitors, with attractive inhibition profiles and good toxicity patterns.
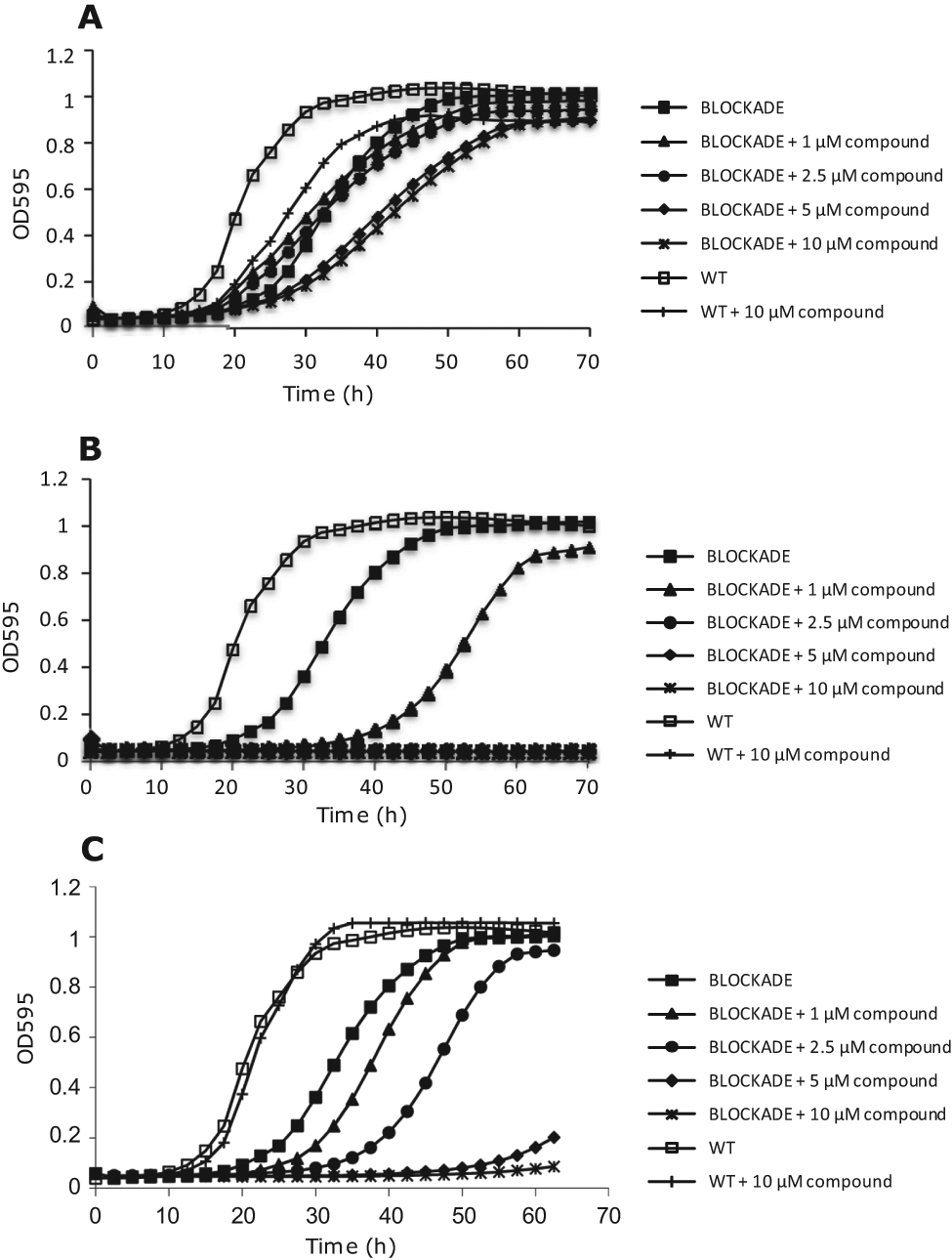
Compounds were classified as false positives (
Of the 291 active compounds identified after the primary screening, 101 were active in the secondary confirmation screening. This resulted in a confirmation rate of 35%, a value higher than ones reported for other screens.22,23 Of the 291 active compounds identified in the primary screening, 44 (15%) were shown to inhibit the growth of WT yeast, hence being discarded as cytotoxic. Confirmation of 101 compounds as hits resulted in a final hit rate of 0.2%. Percentages of inhibition at 10 µM ranged from 50% to 97%, and the majority of the compounds (n = 54) showed inhibition values above 80%. In addition, hits were qualitatively classified based on their capacity to inhibit BLOCKADE growth by more than 50% at concentrations of 1 µM (very good), 2.5 µM (good), 5 µM (average), and 10 µM (weak) (
Robustness of the BLOCKADE assay was verified by calculation of the Z′ factor using the most potent hit, and a value of 0.8 was obtained, clearly validating our system as a potent tool for HTS. To assess the chemical diversity of the 101 hits, we performed a computational assessment to identify structure similarities. Hits were clustered into seven families with distinct chemical profiles, proving that the BLOCKADE assay was able to select groups of structurally unrelated compounds, delivering a diverse set of pharmacophores for further development (data not shown).
Secondary Mammalian Cell Assay
The potency of 99 hits (2 compounds were not available for resupply) from the BLOCKADE screening was validated in a secondary assay using a model of mammalian COS-7 cells transiently transfected with human IDO. Western blotting and immunofluorescence confirmed the expression of IDO ( Fig. 5A,B ). The level of kynurenine in the culture medium is directly proportional to the activity of IDO, and compound inhibition is reflected by a reduction in final kynurenine concentrations. Cells expressing IDO were treated with four different concentrations of compound (0.1, 1, 10, and 100 µM) and after 24 h the amount of kynurenine was measured by HPLC. Figure 5C shows the comparative results obtained for compound BLK2007052 and 1MT. BLK2007052 causes a much stronger inhibition of IDO activity, as kynurenine production is already reduced when cells are treated with low concentrations of compound.
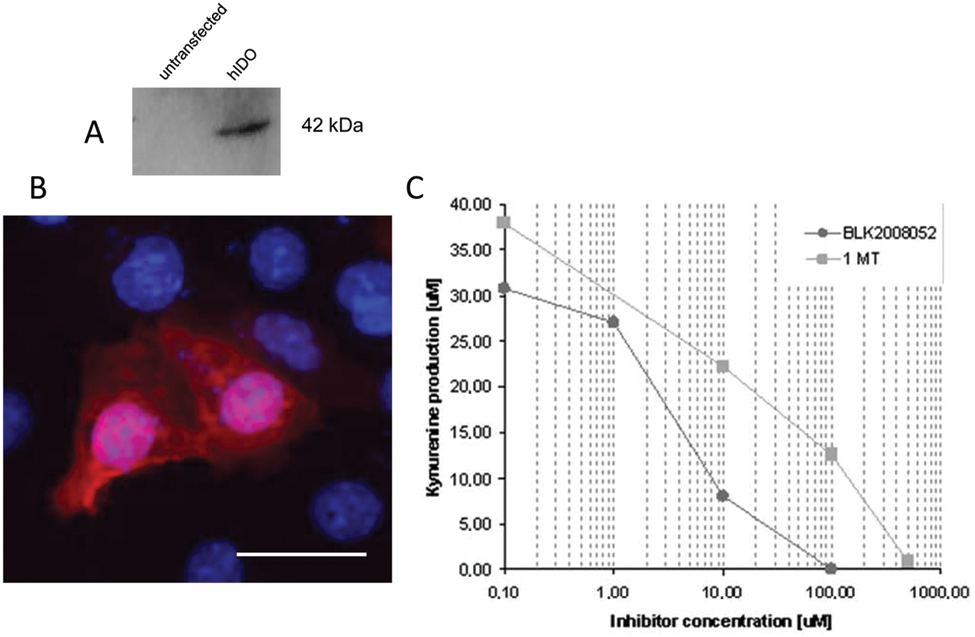
COS-7 cells expressing human indoleamine 2,3-dioxygenase (IDO) were used for a secondary assay. (
The 99 hits were tested in the COS-7 assay, and EC50 was calculated for each. Hits were then ranked according to their potency for IDO inhibition (
Comparison of BLOCKADE Inhibition and COS-7 EC50 Activity
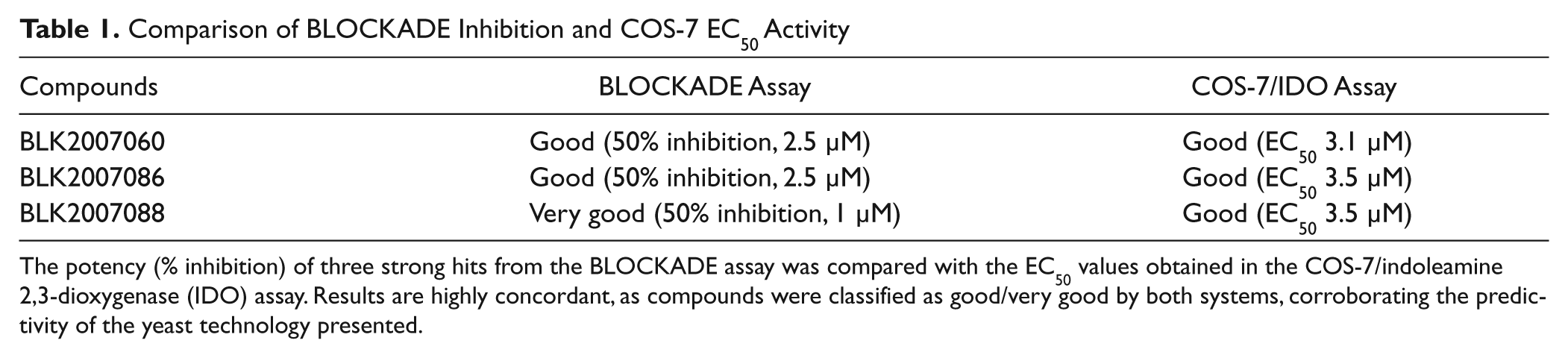
The potency (% inhibition) of three strong hits from the BLOCKADE assay was compared with the EC50 values obtained in the COS-7/indoleamine 2,3-dioxygenase (IDO) assay. Results are highly concordant, as compounds were classified as good/very good by both systems, corroborating the predictivity of the yeast technology presented.
Cytotoxicity Studies
Hit compounds were assayed for cytotoxicity in human hepatoma cells at 2 h and 24 h after treatment. Tests were performed using LDH activity and ATP production, two end points that reflect different biological processes. The LDH assay detects disruption of the cell membrane integrity, a late event in cell death, whereas the ATP assay detects early alterations in cell metabolism. Compounds were tested in six concentrations for the determination of LC50. Low LC50 values correspond to the most hazardous compounds, as low concentrations are enough to induce alterations in cell metabolism and/or cause cell death. Data obtained for the LDH and ATP assays at both time points were combined to generate a final integrated result reflecting the final toxicity of each compound at 2 h and 24 h (
Even after 24 h of incubation, 87% of the identified hits display a safe toxicity profile (low to weak hazard), whereas only 10% of the compounds show a high risk of being hazardous. This high percentage of nontoxic compounds indicates that the use of the BLOCKADE system was able to exclude most cytotoxic compounds, specifically selecting hits that, besides showing a good potency profile in the mammalian cell assay, are expected to be nontoxic for human cells, namely for hepatocytes.
Discussion
The use of IDO inhibitors for human therapeutics has been the focus of much attention in recent years. Current inhibitors show a clear lack of efficacy, as they are weak competitive inhibitors, and new structures are needed to establish pharmacological inhibition of IDO as a treatment. Our goal was to develop a robust yeast-based platform for the identification of new molecules with inhibitory effect toward IDO activity. We generated a humanized yeast strain—BLOCKADE—where both pathways that lead to NAD+ production were abolished by co-deletion of BNA2 and NPT1 genes, making cell survival dependent on the activity of human IDO. Although previous studies suggested that expression of human IDO in yeast was able to promote tryptophan degradation, 21 our work showed for the first time that IDO activity is able to functionally complement disruption of the kynurenine pathway caused by BNA2 deletion. Growth of the BLOCKADE strain was responsive to the level of IDO activity, providing a readout for a life/death assay for the identification of IDO inhibitors.
It is widely recognized that cell-based assays using yeast as a model organism provide greater ease of genetic manipulation and can be screened rapidly at a low cost. We took this concept to an upper level and refined the requisites of the current assay to develop a stringent screening tool that could reduce attrition rates in subsequent phases of drug discovery. Sensitivity of yeast to drugs is increased in strains lacking the two major efflux pumps, PDR5 and SNQ2.24,25 Therefore, most yeast assays are performed using modified strains where pumps are deleted.21,26 On the contrary, the BLOCKADE strain kept all the transporter systems intact, as we proposed to detect only compounds with high specificity. We deliberately expected to lose some potential hits, to ensure we were selecting the most potent compounds. Inhibitory concentrations of hits from the BLOCKADE assay are expected to be higher than the ones reported for other yeast assays, as the BLOCKADE strain is not deleted for the major efflux pumps. Therefore, compounds are being actively pumped out of the cells, and intracellular compound concentration is low and does not reflect the concentration of compound in the medium, which is used for the calculation of potency. Assay conditions are stringent so that only the most strong and selective IDO inhibitors are able to cause a significant growth delay and be identified as hits. Screening of a library of 50 080 small molecules resulted in the identification of 101 compounds with an inhibitory effect in the activity of IDO. This corresponds to a final hit rate of 0.2%, a low value that reflects the strict experimental conditions imposed for this assay, as average hit rates for HTS assays are generally twofold to fourfold higher.27–29
A secondary assay measuring IDO inhibition in mammalian cells confirmed activity for 76% of the hits. To our knowledge, this is the highest predictivity rate reported for a secondary screen of diversity compound libraries,29,30 supporting the strategy of using yeast cells, together with stringent assay conditions, as a valuable tool for the identification of active compounds. In addition, cytotoxicity testing in human hepatocytes demonstrated that 87% of the hits identified displayed a safe toxicity profile (low to weak hazard) after 24 h of incubation. Again, this observation reveals the power of our yeast platform for the selection of compounds with true potential for further development. Data obtained in secondary and toxicity assays strongly validated the robustness of our system, proving that we were able to select compounds with good target specificity and low toxicity profiles, which are less likely to fail in subsequent efficacy tests, thus allowing reduction of attrition rates. It is also relevant that, among the 101 hits, distinct chemical families and pharmacophores were identified, indicating that there was no biased selection toward specific chemotypes and that our system is able to identify chemically diverse molecules. The selected compounds represented a varied source of chemical structures, with seven main families identified. A medicinal chemistry program was initiated to develop new leads with the potential to become future medicines for the treatment of diseases where IDO activation is involved (data not shown).
In summary, our yeast system for the screening of compounds with therapeutic potential offers several benefits over both in vitro enzymatic assays and other cell-based assays commonly used in the initial stages of drug discovery. Unlike enzymatic assays, our system allows targets to be in a physiologically relevant and complex cellular environment, where the activity of compounds is assessed through biological pathways, thus reflecting a true effect on the desired target. Aspects such as membrane permeability, toxicity, and biological stability are immediately addressed, significantly increasing the predictability of the assay and thus reducing attrition rates due to the subsequent identification of undesired liabilities of the initial hit compounds. On the other hand, in opposition to mammalian cell assays, yeast requires very simple and inexpensive growth conditions, presents a fast growth rate, and is extremely amenable to HTS adaptation, which permits the development of screening assays that, besides being extremely informative, are cost-competitive and allow short time frames for hit identification. Finally, the technology described in the present study, based on the example of the BLOCKADE platform, is highly customizable and has already been adapted to different targets relevant to human health and other industry sectors. Screening platforms for compounds targeting Alzheimer disease, tuberculosis, and UV protection, among others, were successfully developed using the same concept and have delivered valuable sets of hits with improved potential for future development due to the selective compound characteristics provided by the use of this stringent yeast-based technology.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. BIOALVO S. A. owns a pending patent on this work, for which several of the authors are inventors (WO2008075991 - yeast platform construction and screening methods).
Funding
This work was supported by the Portuguese SI I&DT–incentive system for technology research and development in companies, Project N. 1413.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.