
Abstract
The phosphatidylinositol 3-kinase (PI3K) pathway couples receptor-mediated signaling to essential cellular functions by generating the lipid second messenger phosphatidylinositol-3,4,5-trisphosphate. This pathway is implicated in multiple aspects of oncogenesis. A low-cost bioassay that readily measures PI3K inhibition in vivo would serve as a valuable tool for research in this field. Using heterologous expression, we have previously reconstituted the PI3K pathway in the model organism Saccharomyces cerevisiae. On the basis of the fact that the overproduction of PI3K is toxic in yeast, we tested the ability of commercial PI3K inhibitors to rescue cell growth. All compounds tested counteracted the PI3K-induced toxicity. Among them, 15e and PI-103 were the most active. Strategies to raise the intracellular drug concentration, specifically the use of 0.003% sodium dodecyl sulfate and the elimination of the Snq2 detoxification pump, optimized the bioassay by enhancing its sensitivity. The humanized yeast-based assay was then tested on a pilot scale for high-throughput screening (HTS) purposes using a collection of natural products of microbial origin. From 9600 extracts tested, 0.6% led to a recovery of yeast growth reproducibly, selectively, and in a dose-dependent manner. Cumulatively, we show that the developed PI3K inhibition bioassay is robust and applicable to large-scale HTS.
Keywords
Introduction
Phosphatidylinositol 3-kinases (PI3Ks) are enzymes that catalyze the phosphorylation of the 3′ hydroxyl group within the inositide ring of phosphatidylinositol. These proteins are involved in the regulation of various essential processes in eukaryotic cells, such as mitosis, cellular growth, survival, and apoptosis. Three classes of PI3Ks exist—class I, II, and III—which are defined by structure and specificity. Class I PI3Ks are heterodimers that consist of a catalytic subunit and a regulatory subunit that phosphorylate phosphatidylinositol-4,5-bisphosphate (PIP2) to produce phosphatidylinositol-3,4,5-trisphosphate (PIP3). Two types of class I PI3Ks have been shown to exist. Class IA PI3Ks are activated by tyrosine kinase receptors (TKRs) and are composed of a 110-kDa catalytic subunit (p110α, p110δ, or p110β isoforms), which binds to a regulatory subunit (p85, p55, or p50 isoforms). Class IB PI3Ks are stimulated by G-protein-coupled receptors (GPCRs), and there is only one member of class IB, p110γ, which associates with the adaptor protein p101/p87PIKAP. The class I PI3K catalytic subunit has an N-terminal domain that binds to the plasma membrane, a Ras-binding domain that attaches to the product of the H-Ras oncogene, a helicoidal domain, and a kinase domain. 1
Upon activation of the cellular receptors by an external signal, the PI3K catalytic subunit translocates to the plasma membrane. Here, it produces the second messenger PIP3, which recruits proteins that contain a Pleckstrin homology (PH) domain, such as protein kinase B, which is also known as Akt. Once Akt is recruited to the membrane, it is activated by the phosphorylation of two different residues, which correspond to Thr308 and Ser473 in the Akt1 isoform. Akt1 regulates the activity of several downstream targets that in turn modulate key cellular processes, including cell growth, proliferation, and apoptosis. 2 Consequently, changes in the levels of PIP3 generated by the class I PI3Ks are directly related to the development of several pathologies, including cancer, which is the most well studied. 3 In fact, the main negative regulator of the PI3K pathway, the PIP3-phosphatase PTEN (phosphatase and tensin homolog), is a tumor suppressor that is often mutated in human malignancies. 4 The role of the PI3Kα isoform in cancer is well established. In addition, the hyperactivation of other PI3K isoforms has also been found in certain types of tumors such as leukemia. 5 In addition to the role of the PI3Ks in tumorigenesis, the PI3K pathway is also known to be involved in many other common pathologies, including allergic reactions, chronic inflammation in asthma patients, and glomerulonephritis. The δ and γ isoforms of PI3K have been reported to regulate mast cell degranulation and lymphocyte chemotaxis. 6 Moreover, PI3K activation has also been implicated in autoimmune diseases, atherosclerosis, and cardiovascular disease. 6 Metabolic disorders, such as diabetes, may also be caused by aberrant PI3K signaling.
Due to the involvement of PI3K in multiple pathologies, it has been given a great amount of attention as a potential pharmacological target in recent years.3,7 This effort has led to the development of several classes of PI3K inhibitors, including small molecules that competitively and reversibly block the adenosine triphosphate (ATP)–binding pocket within the kinase domain. 8 Some of these inhibitors have been reported to be selective for particular α, β, δ, or γ isoforms, whereas others have been shown to serve as pan–class I PI3K inhibitors. Another group of PI3K inhibitors has also been reported to inhibit the structurally related mTOR kinase, which acts downstream of PI3K in the regulation of cell proliferation and inhibition of apoptosis. 9 Many of these molecules, mostly the pan–class I PI3K inhibitors, have been started in clinical trials. Some PI3K inhibitors are currently progressing into phase II clinical trials.8,10 However, until these compounds demonstrate clinical utility, screens for new, structurally diverse PI3K inhibitors are needed to increase the chances of developing potent and effective drugs to face novel therapeutic challenges. In this context, natural products, which have a higher complexity compared with synthetic compounds, may offer a powerful source of chemical diversity for the discovery of novel lead compounds. 11
The model organism Saccharomyces cerevisiae has several features that allow for its use as a tool to study proteins involved in human diseases. For instance, “humanized” recombinant yeast can be devised by cloning a particular human cDNA into a yeast expression vector, and this technique has been widely used.12–14 If the expression of a given heterologous protein in yeast causes a measurable effect, multiple experimental possibilities result, including simple in vivo pharmacological bioassays. We previously reported the development of a humanized yeast system in which we reconstituted the main elements of the PI3K pathway by coexpressing PI3K, PTEN, and Akt1.15,16 Using this system, we demonstrated that the expression of wild-type p110α did not affect yeast growth, but the fusion of this subunit to a prenylation signal from H-Ras (-CAAX box) caused a complete inhibition of yeast growth that was fully dependent on PI3K catalytic activity. Moreover, the expression of the oncogenic gain-of-function mutant p110α (H1047R) was also toxic in yeast cells. These effects resulted from the conversion of essential PIP2 pools to PIP3 and thus were efficiently counteracted by the coexpression of the PTEN phosphatase. 15 In contrast, the Akt1 kinase did not affect yeast growth by itself, but the in vivo activation of Akt1 by the coexpression of wild-type p110α led to an inhibition of yeast growth, which was dependent on Akt1 catalytic activity. 17 In this work, we use the PI3K-dependent cell growth inhibition in yeast to evaluate the performance of competitive PI3K inhibitors in an in vivo experimental setting. To demonstrate that the bioassay provides a readily scalable platform that can be used to screen for novel PI3K inhibitors, we report the application of this assay for a pilot high-throughput screen on a collection of extracts from a microbial origin.
Materials and Methods
Strains and culture conditions
The S. cerevisiae strains used in this work are BY4741 (MATa; his3∆1; leu2∆; met15∆; ura3∆), the isogenic mutant Y03951 (MATa; his3∆1; leu2∆; met15∆; ura3∆, snq2∆::kanMX4), and Y02409 (MATa; his3∆1; leu2∆; met15∆; ura3∆, pdr5∆:: kanMX4), which were obtained from EUROSCARF (Frankfurt, Germany).
The general culture medium used for S. cerevisiae was YPD (1% yeast extract, 2% peptone, and 2% glucose) either in liquid or solid form. For maintaining plasmid selection, synthetic medium (SD; 0.17% nitrogen base for yeast without amino acids, 0.5% ammonium sulfate, and 2% glucose supplemented with the corresponding amino acids) was used. SG and SR media contained 2% galactose or 1.5% raffinose, respectively, instead of glucose. To induce the protein expression of genes regulated by the GAL1 promoter, the yeast cells were incubated in liquid SR medium until mid-log phase was reached; then, 2% galactose was added to the cultures. The yeast strains were incubated at 30 °C.
Quantitative inhibition assays in liquid medium
The yeast transformants carrying the GAL1-driven expression plasmids, YCpLG (empty vector), YCpLG-PI3Kα-CAAX, YCpLG-PI3Kα(K802R)-CAAX, 15 YCpLG-PI3Kα, or YCpLG-PI3Kα(H1047R), 16 either alone or in combination with pYES2-GFP-Akt115 or the empty pYES2 vector (Invitrogen, Life Technologies Ltd, Paisley, UK), were cultured in the appropriate selective SR or SD medium until log phase. From these preinocula, the cultures were then diluted into fresh SR at an OD600 of 0.05 for PI3K inhibition assays or 0.1 for Akt1 inhibition assays. Then, 5 µL of the inocula was seeded into 96-well microplates containing 100 µL SG to induce the expression of the heterologous genes. The LY294002 (Echelon, Salt Lake City, UT; Sigma, St. Louis, MO), 15e (PI3Kα inhibitor II; Echelon), PI103 (Echelon), ZSTK474 (Selleckchem, Houston, TX), and GDC-0941 (Selleckchem) compounds were added in twofold serial dilutions to the plates. To enhance the permeability of the yeast cells, sodium dodecyl sulfate (SDS; Duchefa Biochemie, Haarlem, the Netherlands), 4-nonylphenyl-polyethylene glycol (NP-40) (Fluka, Milwaukee, WI), and Triton X-100 (GE Healthcare, Piscataway, NJ) were added to the medium at the required concentrations. The plates were then incubated at 30 °C. The applied concentration range had been optimized previously by determining the minimum inhibitory concentration (MIC) in the BY4741 wild-type strain. The OD600 was measured at 0, 14, 18, 20, 24, 40, and 48 h in a spectrophotometer for 96-well plates (model 680; Bio-Rad, Hercules, CA). The ratio of growth recovery for each inhibitor at every condition was determined with the following formula: OD600 ai. – OD600 bi. C = X/OD600 ai. – OD600 bi. C = 0, where the numerator is the subtraction of the OD600 after the incubation (ai.) minus the OD600 before the incubation (bi.) of the cells expressing PI3K-CAAX at each inhibitor concentration, and the denominator is the subtraction of the OD600 after the incubation minus the OD600 before the incubation of the cells expressing PI3K-CAAX in the absence of the inhibitor.
Reverse halo assays on solid media
The yeast transformants carrying the expression plasmids encoding the heterologous target proteins were incubated in SR media overnight and then diluted in fresh medium to an OD600 of 0.5. Subsequently, the suspensions were used as inocula and spread on SG agar medium with a sterile cotton swab. A 6-mm disk of nitrocellulose loaded with 15 µL of the compound from a concentrated DMSO stock was placed on the center of the yeast lawn and allowed to diffuse. The plates were incubated at 30 °C for 72 h, and then, the halo of growth around the disk was measured. To enhance the cellular permeability, the detergents SDS (Duchefa Biochemie), NP-40 (Fluka), and Triton X-100 (GE Healthcare) were added to the medium at the required concentrations.
Pilot-scale high-throughput screening
The pilot-scale high-throughput screening (HTS) was performed in 96-well plates using a multichannel pipetting system (Aquarius; TECAN, Männedorf, Switzerland) to add 5 µL per well of each extract from a stock of 2× Whole Broth Equivalent (WBE) in 20% DMSO. As an internal positive control, the PI3K inhibitor LY294002 was added to specific wells at a final concentration of 125, 62.5, 31.25, or 16 µM in each plate.
For the primary screen, an overnight culture of yeast snq2∆ mutant cells (Y03951 strain) transformed with the YCpLG-PI3Kα-CAAX plasmid 15 was diluted to a final OD600 of 0.02 with SG medium containing 0.003% SDS. Then, 100 µL of this culture was added to each well of the plate using an automatic multi dispenser (Multidrop; Thermo, Waltham, MA), except for four wells, which were used as negative controls for cell growth. As a positive control for cell growth, yeast cells transformed with the YCpLG-PI3KK802R plasmid that expressed a catalytically inactive version of PI3K-CAAX 15 were seeded in four wells of each plate in the absence of extract. For the PI3K-induced toxicity control, yeast transformants with YCpLG-PI3Kα-CAAX expressing PI3K-CAAX were also added to four wells of each plate in the absence of extract. For the specificity assay, the same procedure was followed using yeast cells expressing bacterial SigD/SopB from the YCpLG-SigD 18 plasmid instead of PI3K. For the dose-response assay, twofold serial dilutions in 20% DMSO of the PI3K inhibitory extracts were incubated in the presence of yeast expressing PI3K-CAAX. In every assay, the OD600 was measured using a Multi-Detection Microplate Reader Ultra Evolution (TECAN) both before and after a 30-h incubation at 30 °C.
The Genedata Screener software (Genedata, Inc., Basel, Switzerland) was used to process and analyze the screen data. The activity of the extracts was expressed as a percentage of growth recovery, where 100% represented the growth of the yeast that expressed the catalytically inactive PI3K and 0% represented the growth of the yeast that expressed either the active PI3K or SigD/SopB, depending on the assay. The extracts showing ≥75% inhibition of PI3K were considered to be a positive result in the primary and confirmatory assays. The positive extracts that displayed ≤50% activity against SigD/SopB were considered to be specific.
For a linear regression analysis of the assay reproducibility, the coefficient of determination (r2) was calculated. The dose-response curves were fit to the data using the Condoseo application of the Genedata Screener. The curve-fit quality was determined by the goodness of fit (R2). The Z′ and RZ′ factors were calculated as described by Zhang et al. 19 and Kümmel et al. 20
Results
Treatment with PI3K Inhibitors Counteracts PI3K-Dependent Toxicity in Yeast
To optimize a bioassay for the testing of inhibitors of the PI3K/Akt pathway, we analyzed the ability of serial dilutions of the pan–class I selective PI3K inhibitor LY29400221 to rescue the growth of yeast cells that expressed various inhibitory combinations of the heterologous proto-oncogenic kinases. Specifically, we used p110α-CAAX alone as well as combinations of Akt1 with p110α-CAAX, the oncogenic p110α(H1047R) mutant version or wild-type p110α. As we expected, according to the results of our previous work,15–17 the different combinations tested led to growth inhibition, as shown by the low OD600 achieved by these cultures in the absence of the LY294002 inhibitor ( Fig. 1A ). Serial dilutions of the inhibitor within a 50- to 400-µM range were able to partially rescue the growth of the p110α-CAAX-expressing cells; however, no growth rescue was observed for any of the cells coexpressing Akt1 with the different versions of p110α. This result could be because in vivo Akt1 activation in yeast causes alterations in the plasma membrane (M. I. Rodríguez-Escudero, M. Molina, and V. J. Cid, unpublished data), and these alterations could negatively affect the permeability of the cells to the inhibitor.
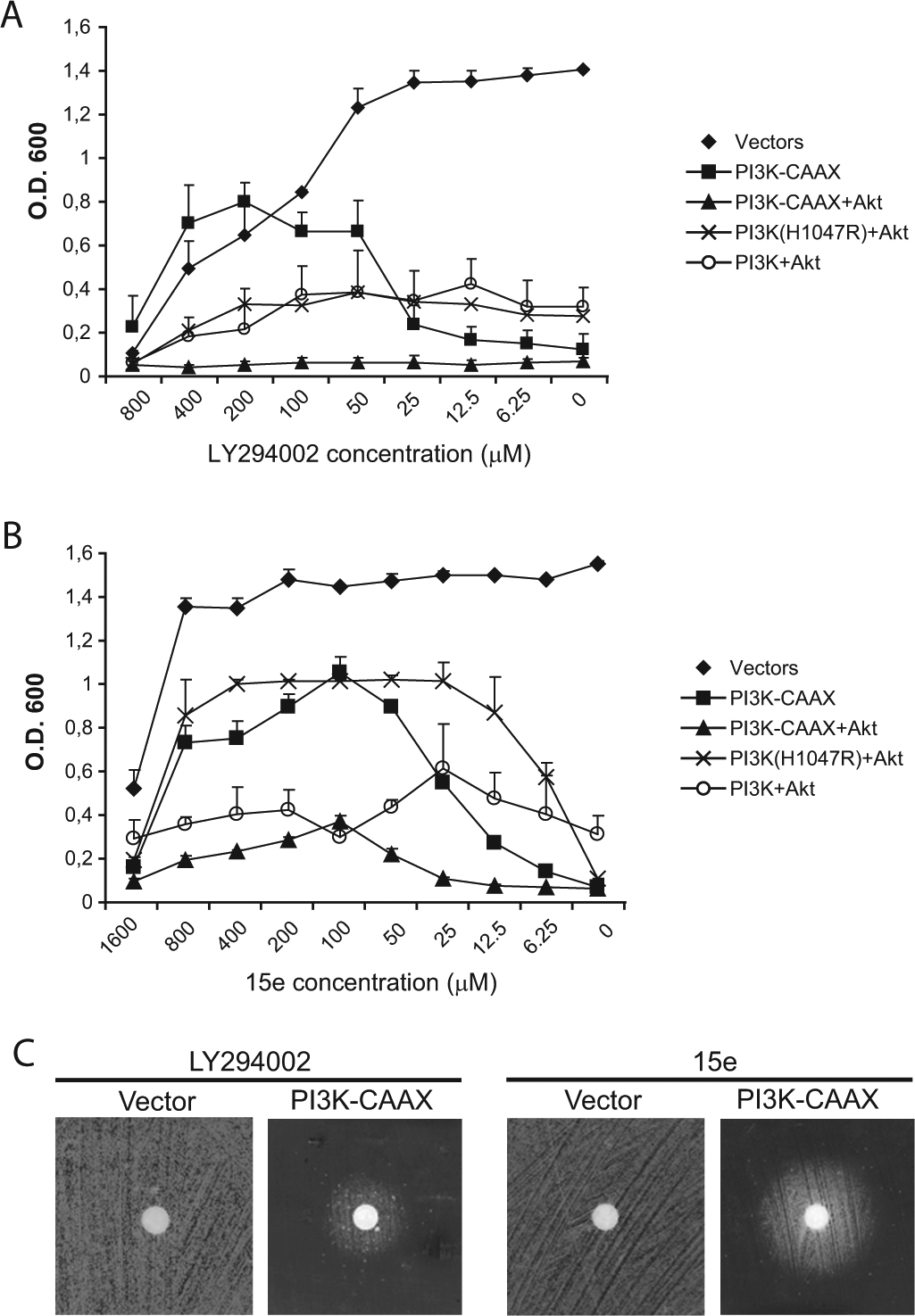
The effect of twofold decreasing concentrations of LY294002 (
We then performed the same assay for the compound 15e, a 4-morpholino-2-phenylquinazoline with a fair selectivity for the p110α isoform of PI3K. 22 As shown in Figure 1B , concentrations of 15e in the range of 25 to 800 µM were able to significantly rescue the growth of cells expressing p110α-CAAX or coexpressing p110α (H1047R) and Akt1; however, treatment with 15e was not able to rescue the growth of cells coexpressing p110α-CAAX with Akt1, which was the most toxic combination due to the additive effects of hyperactive PI3K and in situ–activated Akt1. The lack of growth rescue by both PI3K inhibitors in cells expressing the combination of p110α-CAAX with Akt1 could reflect an enhanced toxicity caused by residual PI3K activity on Akt1. Overall, the concentrations of 15e required to achieve growth rescue by PI3K inhibition were lower than those required for LY294002, and the effective concentration range was also wider for 15e than for LY294002.
Zunder et al 23 previously adapted our yeast model for a qualitative “reverse halo assay” that readily allowed the visualization of PI3K inhibition. Depending on the potency, stability, diffusion ability, and toxicity of a PI3K inhibitor in S. cerevisiae, a halo of growth will appear around a cellulose disk that contains the inhibitor placed upon a lawn of yeast cells expressing p110α-CAAX. As shown in Figure 1C , the growth halo around the disk loaded with 5 mM 15e was larger than the halo obtained with 10 mM LY294002 despite the lower concentration of 15e. Neither of these compounds nor the solvent (DMSO) were toxic for the yeast cells at working concentrations, which was determined by the lack of a halo in the control plates transformed with the empty expression vector ( Fig. 1C ). Therefore, we concluded that 15e is a potent PI3K inhibitor in vivo, as previously described, 22 and our humanized yeast assay is suitable for the detection and analysis of PI3K inhibitors. In light of these results, we decided to use the p110α-CAAX construct for all subsequent experiments because it was very restrictive for yeast growth but readily allowed the rescue of cell growth by both inhibitors tested across a wide concentration range.
The Elimination of the Plasma Membrane ABC Transporter Snq2 Improves the Sensitivity of the PI3K Inhibition Assay
S. cerevisiae is known to express plasma membrane detoxification pumps that efficiently release compounds from the intracellular medium into the extracellular compartment. Commonly, yeast researchers enhance intracellular drug concentrations by using plasma membrane ABC transporter pump mutants, such as pdr1, pdr3, pdr5, and snq2.13,24 We wished to evaluate the influence of these mutations in our bioassay. We found that pdr5 and snq2 mutants grew as efficiently as the isogenic wild-type strains; therefore, we decided to study the performance of these mutants in the PI3K inhibition assay. We found that the presence of the snq2 mutation provided no advantage over the wild-type strain with LY294002 treatment, but the snq2 mutation significantly increased the diameter of haloes with 15e treatment ( Fig. 2 ). We also used a third compound, PI-103, a dual PI3K/mTOR inhibitor, 25 at a 10-mM concentration. With PI-103 treatment, the halo on the wild-type strain was barely detectable but was clearly visible in the snq2∆ mutant ( Fig. 2 ). In contrast, the pdr5 mutant did not enhance the growth halos compared with the wild-type strain (data not shown). Therefore, we concluded that the use of the snq2∆ yeast mutant was advantageous for the PI3K inhibition assay.
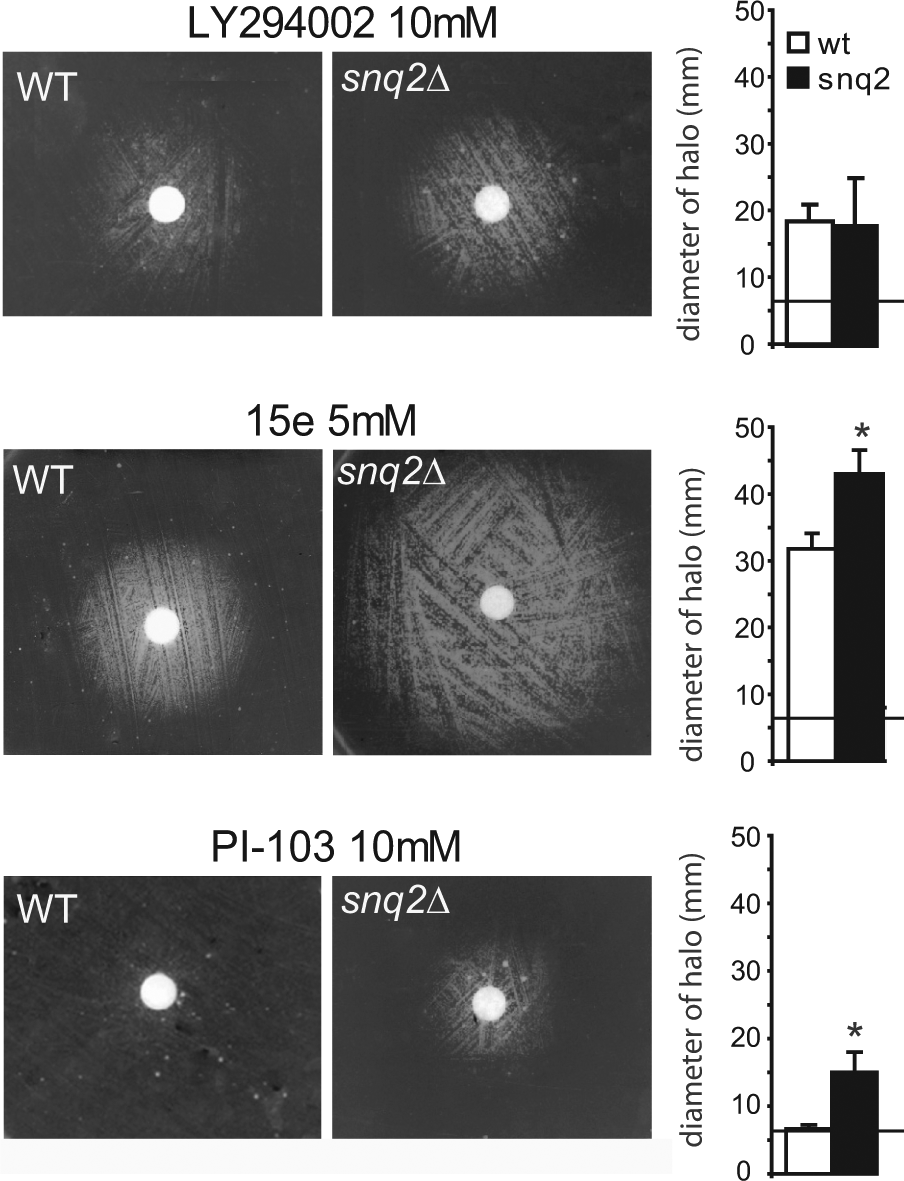
Wild-type BY4741 yeast (WT) and the isogenic snq2∆ Y03951 mutant strain were transformed with YCpLG-PI3Kα-CAAX and spread on SG expression medium in the presence of a disk loaded with 10 mM LY294002, 5 mM 15e, or 10 mM PI-103. The graphs show the average halo diameters obtained from three experimental replicates, and the error bars represent the standard deviation. The diameter of the nitrocellulose disk was 6 mm and is marked with a horizontal line. An asterisk (*) represents a statistically significant difference obtained using a Student t test (p < 0.05).
The Use of Low Concentrations of SDS Increased the Permeability of the Cells to the PI3K Inhibitors
With the aim of further optimizing our system, we hypothesized that the addition of sublethal concentrations of surface-active molecules, such as detergents, to the growth medium should enhance the permeability of the cells to drugs. To determine the optimal conditions for improved cell permeability, we chose three detergents—SDS, NP-40, and Triton X-100—and determined the performance of the bioassay at different detergent concentrations by measuring the halo diameter in the presence and absence of detergent. The addition of 0.001% to 0.005% w/v SDS increased the halo diameters of the snq2∆ strain treated with 10-mM LY294002 disks. The most remarkable increase, which was almost twofold larger than the diameter of the control, was obtained with a 0.003% w/v SDS concentration ( Fig. 3 ). In contrast, the other two detergents did not provide significant advantages at equivalent concentrations. In summary, treatment with SDS allowed a higher intracellular concentration of the compound in yeast cells, which enhanced the sensitivity of the assay.
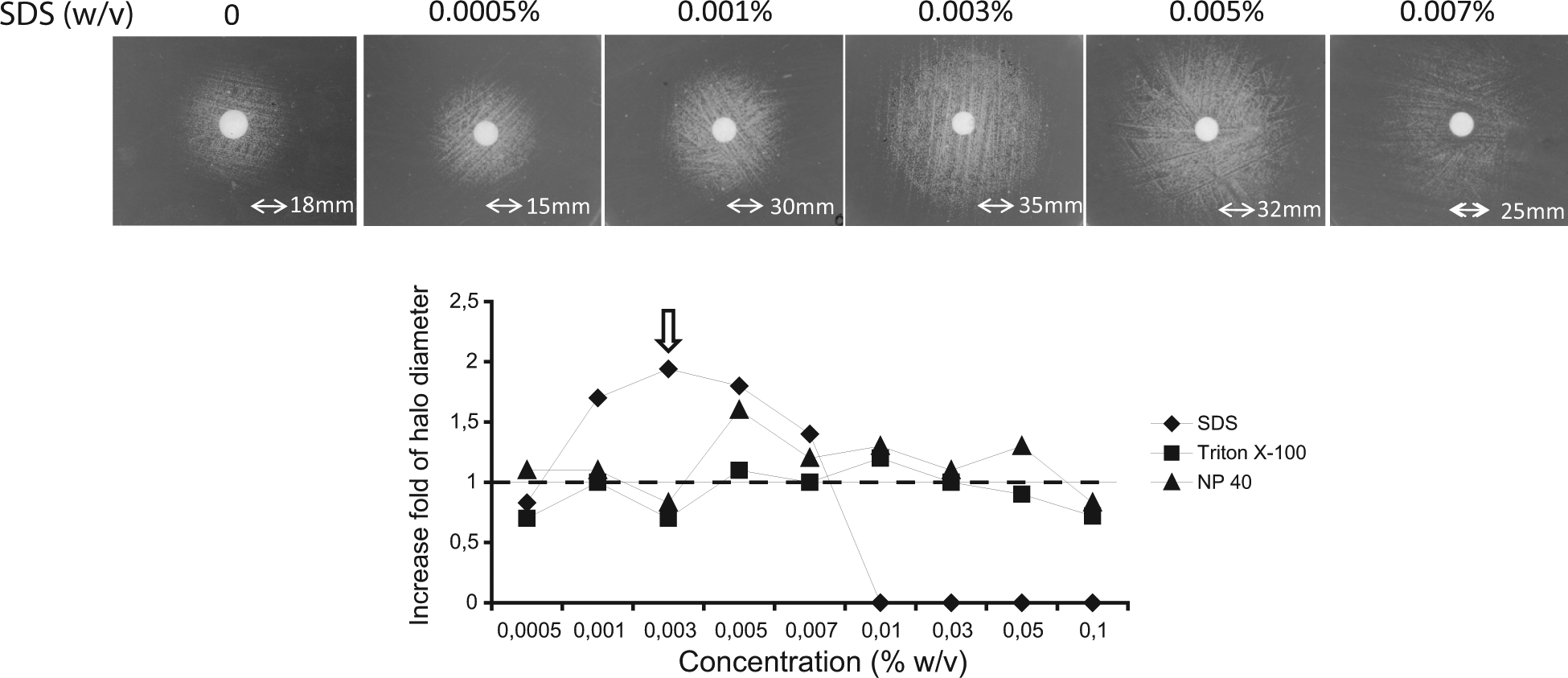
Reverse halo assays performed on the yeast snq2∆ Y03951 mutant strain that expressed PI3K-CAAX from the YCpLG-PI3Kα-CAAX plasmid spread on SG medium containing different concentrations (0.0005%, 0.001%, 0.003%, 0.005%, or 0.007% w/v) of sodium dodecyl sulfate (SDS), Triton X-100, or 4-nonylphenyl-polyethylene glycol (NP-40) in the presence of a disk loaded with 10 mM LY294002. The pictures shown correspond to the experiment performed with SDS (upper panel). In the graph (lower panel), the results are expressed as the fold increase of the halo diameters in the presence of detergent relative to the control halo obtained in the absence of detergent.
A Comparative Assessment of the Optimized Yeast-Based Bioassay with Different PI3K Inhibitors
To confirm the results described above and test the versatility of the bioassay, we used various competitive PI3K inhibitors to evaluate the advantage of the addition of 0.003% SDS to the yeast growth media both in the absence and presence of the snq2 deletion. We performed these experiments with both reverse halo ( Fig. 4 ) and microplate assays ( Fig. 5 ) in parallel. For LY294002 treatment, the effect of SDS addition on the halo enlargement was equivalent in the wild-type and the snq2∆ mutant strains ( Fig. 4A ). In liquid medium, the presence of SDS enhanced the growth rescue in the wild-type strain, and such enhancement was even more effective in the snq2∆ mutant strain ( Fig. 5A ). The growth recovery of the snq2∆ mutant strain with SDS treatment was noticeably higher with a wide range of sublethal concentrations of LY294002 (4-125 µM) than any other versions of the assay ( Fig. 5A ).
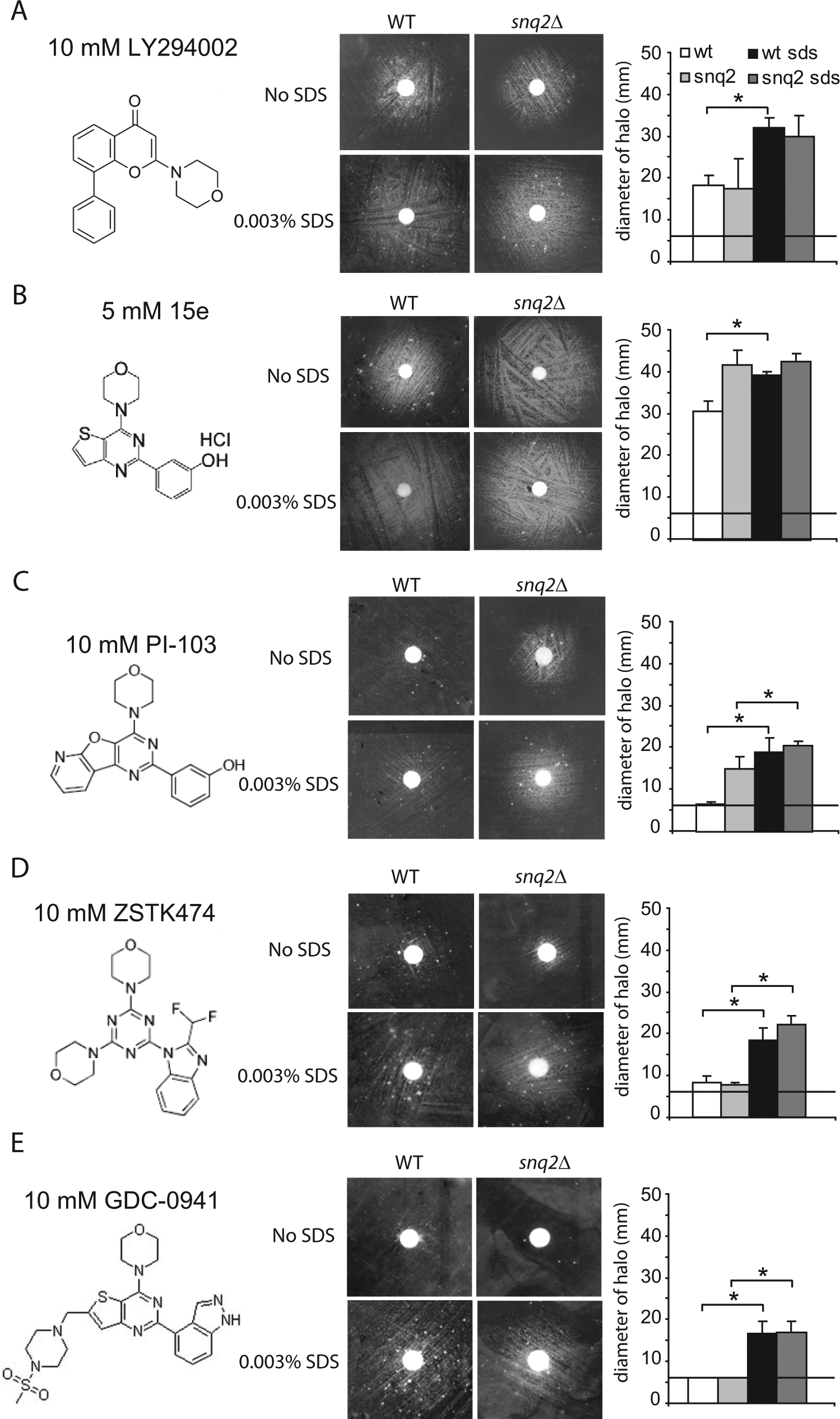
A comparative study of the indicated PI3K inhibitors using the reverse halo assay with wild-type BY4741 yeast (WT) and the isogenic snq2∆ Y03951 mutant strain in the presence or absence of 0.003% sodium dodecyl sulfate (SDS) in SG expression medium. The experiments were performed as described in
Figure 2
. The graphs (
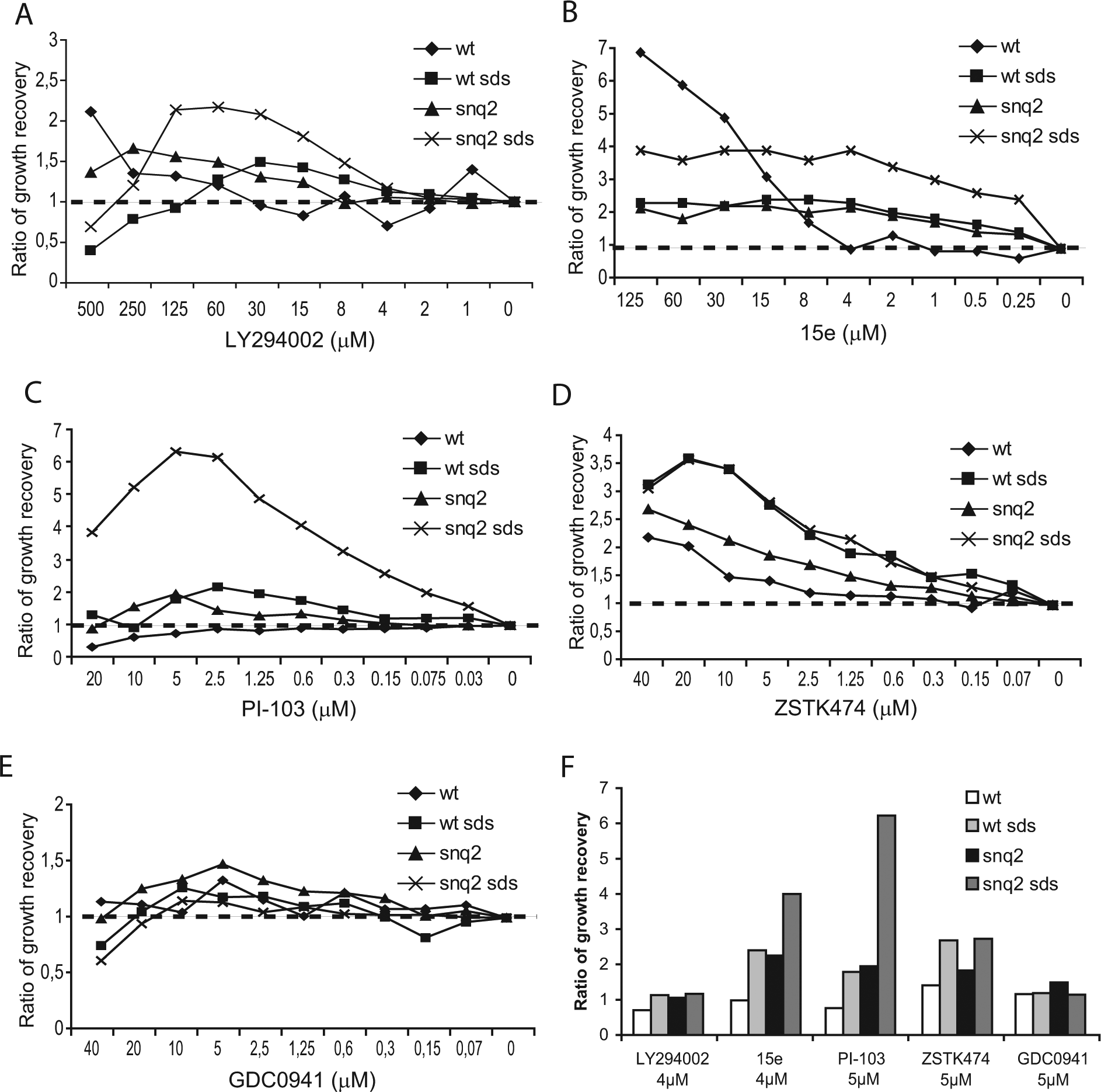
A comparative assessment of the yeast growth recovery obtained by PI3K inhibition in liquid medium. (
For the 15e inhibitor, which rescued the growth of PI3K-expressing yeast very efficiently, the addition of SDS in the reverse halo assay led to halo enlargement in the wild- type strain but did not show an additive effect with the snq2∆ mutant strain, which already had a very large halo ( Fig. 4B ). When the assay was performed in liquid media with a concentration range between 0.25 and 15 µM of 15e, the ratio of growth recovery in the snq2∆ mutant strain treated with SDS was again the highest. These results were closely followed by the ratio of growth recovery in the wild-type strain with SDS treatment and the snq2∆ mutant strain without SDS treatment ( Fig. 5B ) and were again in agreement with the halo results. Thus, both the addition of SDS and the use of the snq2∆ mutant increased the sensitivity of the assay against the 15e compound by two orders of magnitude.
PI-103 and the two pan–class I selective PI3K inhibitory compounds, ZSTK474 and GDC-0941, which are a triazine and thieno[3,2-d]pyrimidine derivative, respectively,26,27 were also assayed in these conditions. GDC-0941 is currently in a phase I clinical trial. 8 With PI-103 treatment, the presence of SDS significantly enlarged the diameter of the haloes ( Fig. 4C ), and in liquid medium, the ratio of growth recovery for the snq2∆ mutant strain with SDS treatment greatly exceeded the recovery ratios for the other conditions tested ( Fig. 5C ). When we tested ZSTK474 with the solid assay in the absence of SDS, the haloes were small and faint; however, the addition of detergent led to clear growth haloes for both the wild-type and snq2∆ yeast strains ( Fig. 4D ). In the liquid assay, the ratio of growth recovery steadily and gradually increased to the same extent over a nM to µM range for both strains in the presence of SDS ( Fig. 5D ). Finally, we also tested GDC-0941, which led to visible haloes in both strains only with SDS-supplemented media, and no differences between the snq2∆ and wild-type strains were observed ( Fig. 4E ). In the serial dilution assay, the growth recovery was poor for all assay conditions tested ( Fig. 5E ), which was likely due to the limited solubility of GDC-0941 in the liquid medium. Figure 5F provides a comparative picture of the potency of the five compounds tested in our yeast bioassay in liquid medium at similar concentrations (4–5 µM) and demonstrates that 15e and PI-103 treatment led to the most efficient inhibition of PI3K. From these results, we concluded that the addition of 0.003% SDS and the use of the snq2 mutant strain improved the sensitivity of the bioassay. Furthermore, we concluded that this system could potentially be useful for screening chemical or natural libraries for PI3K inhibitors with specificities and potencies similar to the known compounds tested here.
Adaptation of the Yeast Bioassay to HTS: A Pilot Assay on Microbial Extracts
On the basis of the optimized yeast bioassay for PI3K inhibition, we decided to develop a pilot HTS using a collection of extracts obtained from fungal and actinomycete cultures.
28
First, the high-throughput conditions were validated with an initial group of 2560 extracts in 34 plates. The samples for the assay validation were chosen randomly from the extract collection. From this initial screen, we identified 80 extracts that showed a significant yeast growth recovery (
In light of these results, we decided to scale up the high-throughput screen to include 9600 extracts. In addition, we introduced further steps in the screening routine to test the specificity and dose dependence of the identified hits. A summary of the results from this pilot screen is shown in
Discussion
PI3K is a prominent target for drug discovery in multiple pathologies, including cancer. Although some PI3K inhibitors are now showing promising results in phase II clinical trials, novel lead compounds are still needed. 8 Therefore, considerable effort is currently being invested in the establishment of screening platforms to identify and characterize PI3K inhibitors of potential pharmacological interest. Experimental setups have been described for this purpose using both in vitro and in vivo assays. The strategies for the in vitro assessment of PI3K activity are specific and allow very precise and reliable quantitative measurements. Recent advances in these methods include the development of fluorescence-based assays to replace radioactive methods 29 and the optimization of these methods using state-of-the-art techniques such as ratiometric fluorescence superquenching. 30 The advantages of the latter method include a high sensitivity and broad application to any PI3K isoform. In vivo PI3K assays have also been devised, including the use of Chinese hamster ovary (CHO) cells cotransfected with the human insulin receptor and an AKT1-GFP reporter to monitor the redistribution of Akt1. 31 Although in vitro assays are a shortcut in the search for compounds that are specific for the target under investigation, they have several disadvantages over in vivo bioassays. First, the cost of reagents required for these assays (e.g., purified enzyme, substrates, and sensing molecules) is very high, which is an important drawback for the application of these assays to screen large compound libraries. Second, the cellular environment is overlooked in an in vitro experimental setting. Therefore, a hit from an in vivo screen proves that the compound is able to interact with the target within a biological system and is one step closer to potential therapeutic use.
In this study, we propose an alternative in vivo assay that is based on the heterologous expression of PI3K in a ready-to-use yeast system. Similar work with other mammalian targets expressed in S. cerevisiae have greatly contributed to drug discovery research. 12 For example, using a yeast system, Zhang et al. 32 identified a new inhibitor of protein aggregation in Huntington disease, and Tribouillard et al. 33 discovered novel antiprionic molecules. Major advantages of our yeast-based bioassay over the currently available methods for the HTS of PI3K inhibitors include the ease of manipulation and a very low cost. The activity of the inhibitor in our yeast system is simply monitored by observing cell growth. Thus, our assay overcomes the requirements for purified PI3K, fluorescence-labeled phosphoinositides, or other reagents required for in vitro assays. Moreover, yeast cultures are easier to handle and less expensive than mammalian cell cultures. Based on these features, our bioassay has great potential as an efficient primary HTS that can be used to prioritize compounds for subsequent, more costly screens. These secondary analyses would discriminate specific kinase inhibitors from nonspecific preselected hits, such as compounds that may interfere with the yeast prenylation machinery that is required for the toxicity of PI3K-CAAX.
The validation of the humanized yeast system as a PI3K inhibition bioassay was performed using known commercial PI3K inhibitors. The optimization of the assay involved finding restrictive conditions in which the yeast growth was severely affected by the expression of the heterologous target and, subsequently, enhancing the intracellular concentration of the compounds. Our experimental system allows the evaluation of PI3K inhibitors either on solid media by measuring the growth haloes around compound-loaded disks or in liquid media at a microtiter scale by determining the nontoxic PI3K-inhibitory concentrations able to recover yeast growth. The former method, called the “reverse halo assay,” is simple, fast, visual, and easy to interpret. In contrast, the smaller amounts of inhibitors required and a higher sensitivity make the liquid media assay more convenient for HTS. Growth interference yeast-based assays have several advantages. First, compounds that effectively inhibit the target protein restore growth, and thus, molecules that are generally toxic to cells are not selected. 24 Second, the specificity of the assay should provide insight into the mechanism of action of the potential inhibitors on the heterologous target being investigated. Inhibitors that do not act directly or selectively against the expressed target should not be detected in the yeast bioassay. For example, resveratrol did not inhibit PI3K activity in our assay (data not shown), which suggests that the resveratrol-mediated downregulation of the PI3K pathway 34 is not through a direct inhibition of the PI3K catalytic subunit.
Because the effect of a compound on a particular target can be influenced by other factors that are only present in mammalian cells, S. cerevisiae may not always be the most appropriate environment to evaluate the activity of an inhibitor. Currently, since other PI3K isoforms do not give rise to the same degree of toxicity when expressed in yeast, the application of our bioassay is restricted to the study of the α isoform of p110. 16 Enhancing the expression by using more efficient yeast promoters should facilitate the extension of our system to other PI3K isoforms. In addition, S. cerevisiae possesses several plasma membrane pumps that release toxic compounds from the intracellular compartment. Yeast strains with deletions of the genes encoding these detoxification pumps have been widely used to overcome this issue.23,24 Here, we report that in our particular assay, the use of the snq2∆ mutant strain, which lacks a plasma membrane transporter involved in detoxification, enhances the PI3K inhibition in vivo compared with the isogenic wild-type strain. In addition, we found that the use of 0.003% SDS to increase cell permeability was also beneficial for the growth rescue achieved by all of the inhibitors tested in both the wild-type and snq2∆ strains. The combination of SDS with the snq2 strain yielded the best assay sensitivity. Also, our results show that the expression of PI3K versions that cause a strong effect on S. cerevisiae growth, such as the membrane-directed p110α-CAAX, are more adequate for the bioassay than the PI3K versions that cause a mild (e.g., PI3K/Akt1) or drastic growth effect (e.g., p110α-CAAX/Akt1).
The validation of our system with known PI3K inhibitors demonstrated the varied potencies of the tested drugs. The compounds derived from 1,4-morpholino-2-phenylquinazolines (i.e., 15e and PI-103) 22 resulted in the most effective PI3K inhibition. The p110α selective inhibitor, 15e, caused the largest halo diameter in the solid assays, whereas the growth recovery in liquid medium was higher for the dual PI3K/TOR inhibitor PI-103 at lower concentrations. Both 15e and PI-103 treatments were more potent than the pan–class I PI3K inhibitor LY294002, which is in agreement with the literature.7,21
To demonstrate that our bioassay was suitable for HTS, we implemented a pilot-scale screen using a collection of natural extracts of microbial origin. We chose this compound library because traditional chemical libraries have already been thoroughly screened and have resulted in a plethora of encouraging candidates. After this extensive chemical mining, we hypothesized that lead compounds with alternative structures and novel mechanisms are more likely to be found in natural sources, which offer a different display of chemical diversity. 11 Although the characterization of the hits identified in this study will require further experimentation, the results from our pilot-scale HTS demonstrate that the yeast PI3K inhibition bioassay is robust enough for extensive HTS. Of the 9600 extracts analyzed, 55 (0.6%) passed the reproducibility, selectivity, and dose dependence criteria. This screening hit rate is within the range that has been published for primary screening processes using natural compounds. 35 Additional in vitro PI3K inhibition assays and antiproliferation assays in PI3K-mutated breast cancer cell lines are under way to test the specificity of our preselected hits. Our preliminary results on a subset of 20 positive extracts have demonstrated that 5 of these extracts have activity in subsequent assays (data not shown), which provides additional evidence that yeast primary screens positively select specific hits. The activity of the positive extracts could be due to a single compound in the extract or a synergistic effect of the mixture. It is hoped that future work will allow the isolation and characterization of the molecules in these extracts responsible for the PI3K inhibition.
In conclusion, we have established optimal conditions for a yeast-based PI3K inhibition assay, in which the balance of potency and toxicity allows for an effective and selective inhibition of the target. Furthermore, we proved our system to be robust and reproducible in a pilot-scale HTS for the search of new PI3K inhibitory compounds, which may eventually contribute to anti-PI3K-based chemotherapy.
Footnotes
Acknowledgements
We are grateful to our colleagues from Unit 3 at the Universidad Complutense de Madrid for help and discussion and from Fundación Medina in Granada, especially Mercedes de la Cruz, for technical assistance.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant (BIO2010-22369-C02-01) from the Ministerio de Ciencia e Innovación (Spain) and the Program for UCM Research Groups (920628) from BSCH-UCM to M.M. and by Fundación MEDINA, a public-private partnership of Merck Sharp & Dohme de España S.A./Universidad de Granada/Junta de Andalucía. T.F-A. was also supported by a predoctoral fellowship from the Ministerio de Educación y Ciencia (Spain).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.