
Abstract
The most widely used assay format for characterizing plasma membrane transporter activity measures accumulation of radiolabeled substrates in tissues or cells expressing the transporters. This assay format had limitations and disadvantages; therefore, there was an unmet need for development of a homogeneous, nonradioactive assay for membrane transporter proteins. In this report, the authors describe the development of a label-free homogeneous assay for the sodium-dependent phosphate transporter NaPi-IIb using the Epic system. The addition of phosphate stimulated a dynamic mass redistribution (DMR) profile unique to cells expressing NaPi-IIb but not on parental cells. This DMR profile was phosphate specific because sulfate or buffer alone did not elicit the same response. Furthermore, the DMR response observed was phosphate and sodium dependent, with Km values in the micromolar and millimolar range, respectively. A known NaPi-IIb noncompetitive inhibitor was shown to completely inhibit the phosphate-stimulated DMR response, suggesting that this observed DMR response is an NaPi-IIb–mediated cellular event. The results demonstrate that a novel label-free assay was developed for studying transporter-mediated cellular activity, and this DMR assay platform could be applicable to other membrane transporter proteins.
Introduction
Inorganic phosphate (Pi) is a critical component of bone mineralization, membrane phospholipids, and nucleotides, which provide energy as well as the backbone of DNA and RNA. Because Pi cannot be synthesized in the body, its presence is largely dependent on ingestion of Pi from the diet.
1
The kidney is a major regulator of Pi homeostasis and can increase or decrease its Pi reabsorption capacity to accommodate Pi retention. In chronic renal failure, Pi retention may exacerbate secondary hyperparathyroidism and uremic bone disease and progress to end-stage renal disease.
2
Another mechanism to control the Pi level in the blood is the regulation of intestinal Pi absorption, which occurs by both sodium-dependent and sodium-independent processes mainly in the proximal small intestine.
3
The sodium-dependent mechanism is mediated by a type IIb sodium-phosphate cotransporter, NaPi-IIb, which is expressed at the brush-border membrane of enterocytes.
4
This NaPi-IIb transporter is electrogenic and exhibits an Na+:
Despite advances in recent years in cellular assay technologies, few assays are available for the study of transporter activity. The most widely used assay format today measures the accumulation of radiolabeled substrates in tissues or cells that express the target transporters. In measuring NaPi-IIb transporter activity, [33P]orthophosphate is commonly used as the labeled substrate. The transport of [33P]orthophosphate from the extracellular buffer to inside NaPi-IIb–expressing cells is determined by incubating the cells with buffer containing the radiolabeled substrate for a defined period of time either at 37 °C or room temperature. 7 However, the use of radiolabeled substrates in a high-throughput assay has its limitations: a wash step may be needed, the use of radiolabeled substrate of high specific activity in a homogeneous assay format can result in high background, and the handling and storage of radioactive waste can be time-consuming and costly. More importantly, the amount of unlabeled substrate that can be added is limited because of dilution of the radiolabeled substrate transport; therefore, physiological concentrations of substrate cannot be implemented in this assay format. Thus, there is an unmet need for developing a homogeneous, nonradioactive assay in a high-throughput format to study transporter proteins.
In this report, we describe the development of a novel label-free NaPi-IIb transporter assay using the Epic system. This system uses resonant waveguide grating biosensors to monitor molecular movements inside cells, termed dynamic mass redistribution (DMR), upon the activation/modulation of cell surface targets. 8
Materials and Methods
Materials
All cell culture reagents were purchased from Life Technologies Corp. (Carlsbad, CA). Epic 384-well microplates were obtained from Corning (Corning, NY). [33P]orthophosphoric acid and Cytostar-T 384-well microplates were purchased from PerkinElmer (Waltham, MA). Adenosine triphosphate (ATP) was purchased from Sigma (St. Louis, MO). Chemical compounds used in this study were obtained from the Amgen compound collection.
Cell Culture and Western Blot Analysis of the Expression of NaPi-IIb Protein
The Chinese hamster ovary (CHO) cell line stably expressing the human NaPi-IIb transporter gene (SLC34A2) was developed at Amgen and maintained in a growth medium containing DMEM supplemented with 10% fetal bovine serum, 1 mg/mL G418, 1 mM glutamine, 0.1 mM nonessential amino acids, 1x HT supplement, 100 U/mL penicillin, and 100 µg/mL streptomycin. The parental CHO cell line, transfected with the same expression vector but without the NaPi-IIb gene, was maintained in the same growth medium.
For cell membrane preparation, NaPi-IIb–CHO cells or parental CHO cells were washed with phosphate-buffered saline (PBS) and harvested individually by scraping in PBS. The cell suspension was centrifuged at 1900 g for 10 min, and the resulting cell pellet was frozen at −80 °C until membrane preparation. The cell pellet was thawed on ice and suspended in a homogenization buffer (50 mM Tris·Cl pH 7.4, 1 mM EDTA, and complete EDTA-free protease inhibitor; Roche Applied Science; Indianapolis, IN). The cell suspension was homogenized with a Potter-Elvehjem homogenizer and centrifuged at 1000 g for 12 min at 4 °C in a J2-MC centrifuge using an F8-4x1000y rotor. The supernatant was removed and centrifuged at 100,000 g for 1 h at 4 °C in a Beckman Optima XL-100k centrifuge using a F40L-8x100 rotor to produce a crude membrane particulate fraction. The cell membrane pellets were suspended in an appropriate volume of cell lysis buffer (50 mM Tris·HCl pH 7.5, 150 mM NaCl, 1% NP40, 0.25% sodium deoxycholate, 0.7 µg/mL pepstatin, 1 mM NaVO4, 1 mM EDTA, 1 mM NaF, and complete EDTA-free protease inhibitor), and protein concentrations were determined using the Coomassie protein assay reagent (Pierce, Rockford, IL). The lysates were resolved by SDS-PAGE and transferred onto nitrocellulose membrane (Life Technologies). The blot was blocked for 1 h in Tris-buffered saline–Tween (TBST; 20 mM Tris·HCl pH 7.6, 137 mM NaCl, 0.1% Tween-20) containing 5% milk (blocking buffer) before incubation with a murine anti-human NaPi-IIb antibody (diluted 1:500) at 4 °C overnight. The antibody was raised against a peptide corresponding to the N-terminus of human NaPi-IIb sequence at Amgen. After three washes with TBST, the blot was incubated with peroxidase- conjugated secondary antibody (diluted 1:1000) at room temperature for 1 h. The antibody binding was detected using enhanced chemiluminescence. Figure 1 shows that NaPi-IIb protein was expressed and detected in NaPi-IIb–CHO cell membranes in a protein concentration–dependent manner but not in the parental CHO cell membranes.
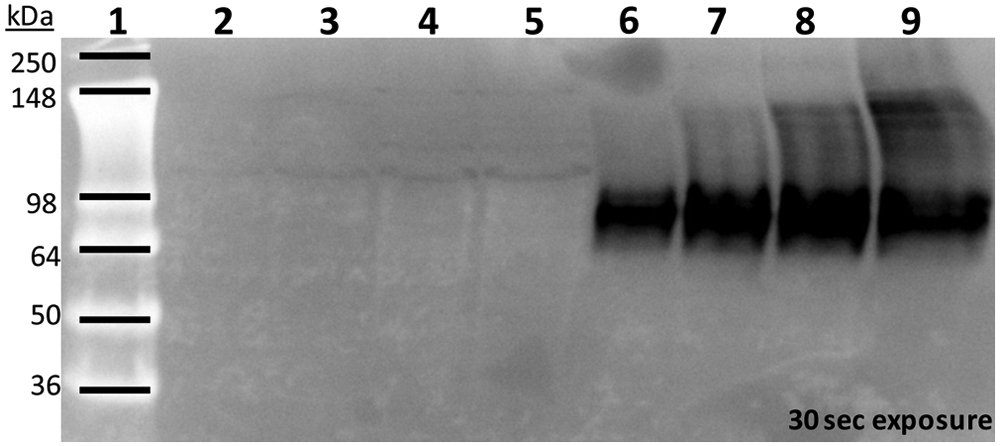
Western blot analysis of human NaPi-IIb protein in the NaPi-IIb–CHO cell membrane. NaPi-IIb protein was detected in the total membrane protein prepared from NaPi-IIb–CHO cells but not from parental CHO cells. Lane 1 is SeeBlue Plus2 prestained standard (Life Technologies). Lanes 2 to 5 are parental CHO cell membrane protein samples at increasing amounts: 25 µg (lane 2), 50 µg (lane 3), 100 µg (lane 4), and 150 µg (lane 5). Lanes 6 to 9 are NaPi-IIb–CHO cell membrane protein samples at increasing amounts: 25 µg (lane 6), 50 µg (lane 7), 100 µg (lane 8), and 150 µg (lane 9).
Measurement of Epic DMR Response
NaPi-IIb–CHO cells or parental CHO cells were seeded on 384-well fibronectin-coated Epic microplates at 9000 cells per well in 40 µL of growth medium and cultured at 37 °C in an atmosphere of 5% CO2. The next day, cell culture media were removed, and the cells were washed once with an assay buffer containing 10 mM HEPES pH 7.2 (Life Technologies), 137 mM NaCl, 5.4 mM KCl, 2.8 mM CaCl2, 1.2 mM MgCl2, 10 mM D-glucose, and 5 mM NaHCO3. Test compounds were diluted in the assay buffer and added to the cells to obtain a final volume of 40 µL in each well. The microplate was equilibrated with test compounds in the Corning Epic system (Corning, NY) at 26 °C for 1 h prior to a 5-min baseline measurement. After recording the baseline activity, 20 µL of KH2PO4/K2HPO4 (K-Pi) prepared in the assay buffer at the concentrations indicated in the text was added, and the DMR response, which measures a shift in the resonance wavelength (pm), was monitored for an additional 65 min.
In the sodium-dependent studies, NaCl was diluted in assay buffers containing 0 to 137 mM of choline chloride so that the sum of NaCl and choline chloride concentrations was 137 mM. The cells were equilibrated in the various NaCl concentrations for 1 h in the Epic reader before baseline measurement. The cells were then stimulated with 100 µM K-Pi prepared in the corresponding NaCl-choline chloride dilution buffers, and the DMR response was recorded for an additional 65 min. NaHCO3 was not included in the assay buffer for the sodium-dependent studies so that the various sodium concentrations could be accounted for accurately.
For concentration-response studies and IC50 determinations, the DMR response was calculated using the 35-min time point at which the buffer effect remained at the baseline prior to the gradual increase.
Percentage activity (%) was calculated as follows:

The positive control wells were preincubated with DMSO at a concentration equivalent to that present in the compound wells prior to the addition of 100 µM K-Pi. The negative control wells were preincubated with 20 µM of compound 1, an NaPi-IIb transporter noncompetitive inhibitor, which completely inhibited NaPi-IIb activity, as was reported previously. 7 Compound IC50 values were determined by nonlinear regression analysis of the concentration- response curves using Prism (GraphPad Software, San Diego, CA).
[33P]-Uptake Assay in NaPi-IIb–CHO Cells
The measurement of [33P]-uptake in NaPi-IIb cells was similar to that described previously, except it was performed in a homogeneous scintillation proximity assay (SPA) format using Cytostar-T plates. 7 NaPi-IIb–CHO cells were seeded on Cytostar-T 384-well microplates at 10,000 cells per well in 30 µL of growth medium and cultured at 37 °C in an atmosphere of 5% CO2. The next day, cell culture media were removed, and the cells were washed once with an assay buffer containing 15 mM Tris·Cl pH 7.4, 137 mM NaCl, 5.4 mM KCl, 2.8 mM CaCl2, and 1.2 mM MgSO4. Test compounds were diluted in the assay buffer and added to the cells 30 min prior to the addition of [33P]-orthophosphoric acid (75 ρM). After 2 h of incubation at room temperature, radioactivity accumulation in the cells was determined using the PerkinElmer Topcount reader (Waltham, MA). Percentage activity and IC50 calculation were similar to that described for the DMR measurement.
Results and Discussion
Effects of Phosphate on NaPi-IIb–CHO Cells
The addition of 100 µM K-Pi elicited a positive DMR response in NaPi-IIb–CHO cells ( Fig. 2A ), and the response was reproducible in all experiments. A gradual and continuous increase in signal was observed approximately 6 min after K-Pi addition, and the rate became steady after 30 min. The response observed at the end of the measurement corresponded to a wavelength shift of 80 pm. When the assay buffer was added to the cells in the absence of K-Pi, no initial response was generated. However, a small DMR response drift was detected after 40 min, which reached 15 pm at the end of the measurement. The addition of 100 µM potassium sulfate (K2SO4) did not elicit a response for up to 40 min, showing a kinetic profile similar to the assay buffer alone. Thus, NaPi-IIb expressed on the plasma membrane of this engineered cell line is a Pi-specific transporter responsible for transporting Pi, and not sulfate, into cells.
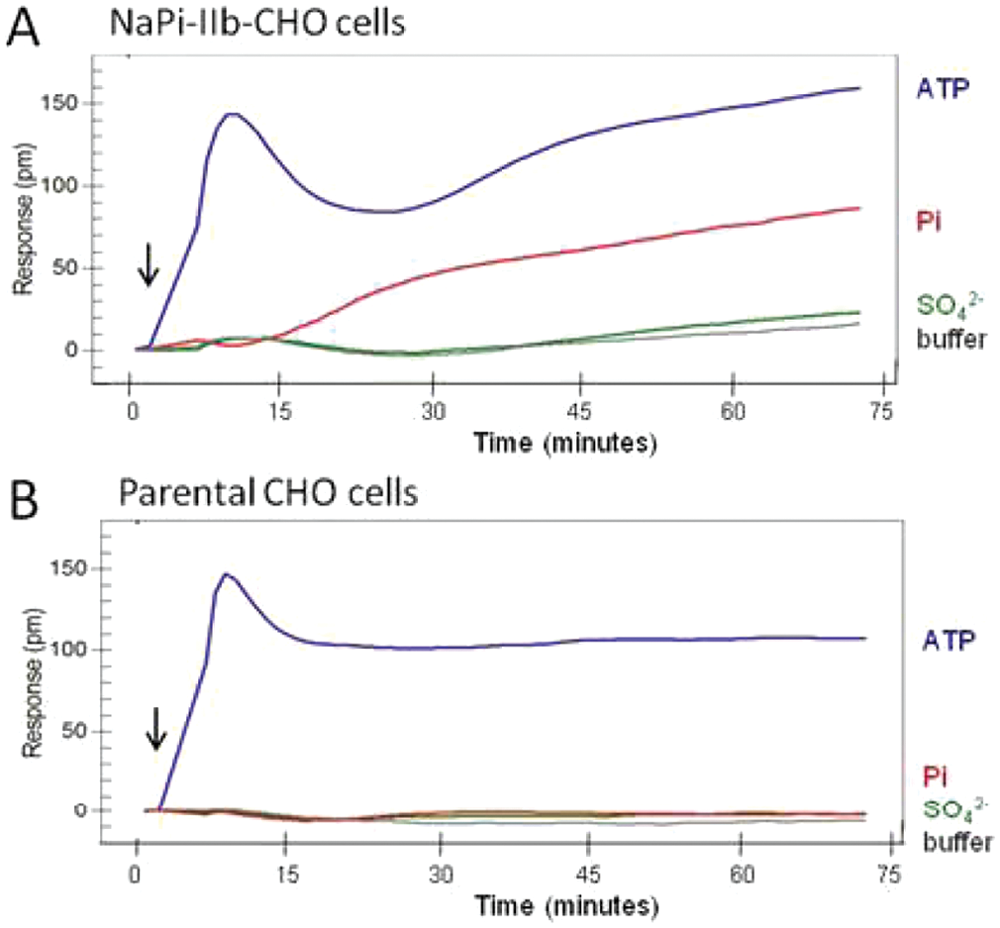
The dynamic mass redistribution (DMR) profiles in NaPi-IIb–CHO cells and parental CHO cells. NaPi-IIb cells (
Neither K-Pi nor K2SO4 stimulated a DMR response in the parental CHO cells (
The ATP-induced DMR responses in the NaPi-IIb–CHO cells and the parental CHO cells were identical at the initial peak, but a significant difference developed after 30 min. A second increase in DMR response was observed in the NaPi-IIb–CHO cells but not in the parental CHO cells ( Fig. 2A ). Interestingly, this second phase increased at a rate parallel to the K-Pi–induced response, suggesting that this may be an NaPi-IIb transporter–mediated mechanism. Whether ATP, at this concentration, acts as a substrate to the NaPi-IIb transporter remains to be elucidated.
Phosphate and Sodium Dependence
Addition of K-Pi, at various concentrations (0.1, 0.3, 0.6, 1.0, 1.8, 3.0, 10, 30, 100 µM), to NaPi-IIb–CHO cells in the presence of 137 mM NaCl elicited a DMR response in a concentration-dependent manner ( Fig. 3A ). The maximum signal, equivalent to a wavelength shift of 50 pm, was observed and reached a plateau at 10 µM and higher concentrations ( Fig. 3B ). The calculated Km value for Pi is 1.4 ± 0.3 µM (n = 4). This value is lower than those observed in murine NaPi-IIb (Km = 50 µM), which was determined using Xenopus oocytes injected with murine NaPi-IIb cRNA. 10 Similarly, the sodium dependence of the NaPi-IIb transporter was determined. NaPi-IIb–CHO cells were equilibrated in an assay buffer containing various concentrations of NaCl (5, 7, 13, 18, 25, 35, 60, 100, 137 mM) for 1 h prior to stimulation by 100 µM K-Pi, which was diluted in the corresponding assay buffers. Choline chloride was added to substitute NaCl so that the same osmolarity was maintained in the assay buffers containing various NaCl concentrations. Addition of K-Pi elicited a DMR response in a sodium-dependent manner ( Fig. 3C ). The response was initially detected at 18 mM NaCl and reached a plateau at 100 mM NaCl. The calculated Km value for sodium is 26.8 ± 2.8 mM (n = 3), which is similar to that reported on murine NaPi-IIb (Km = 33 mM). 10
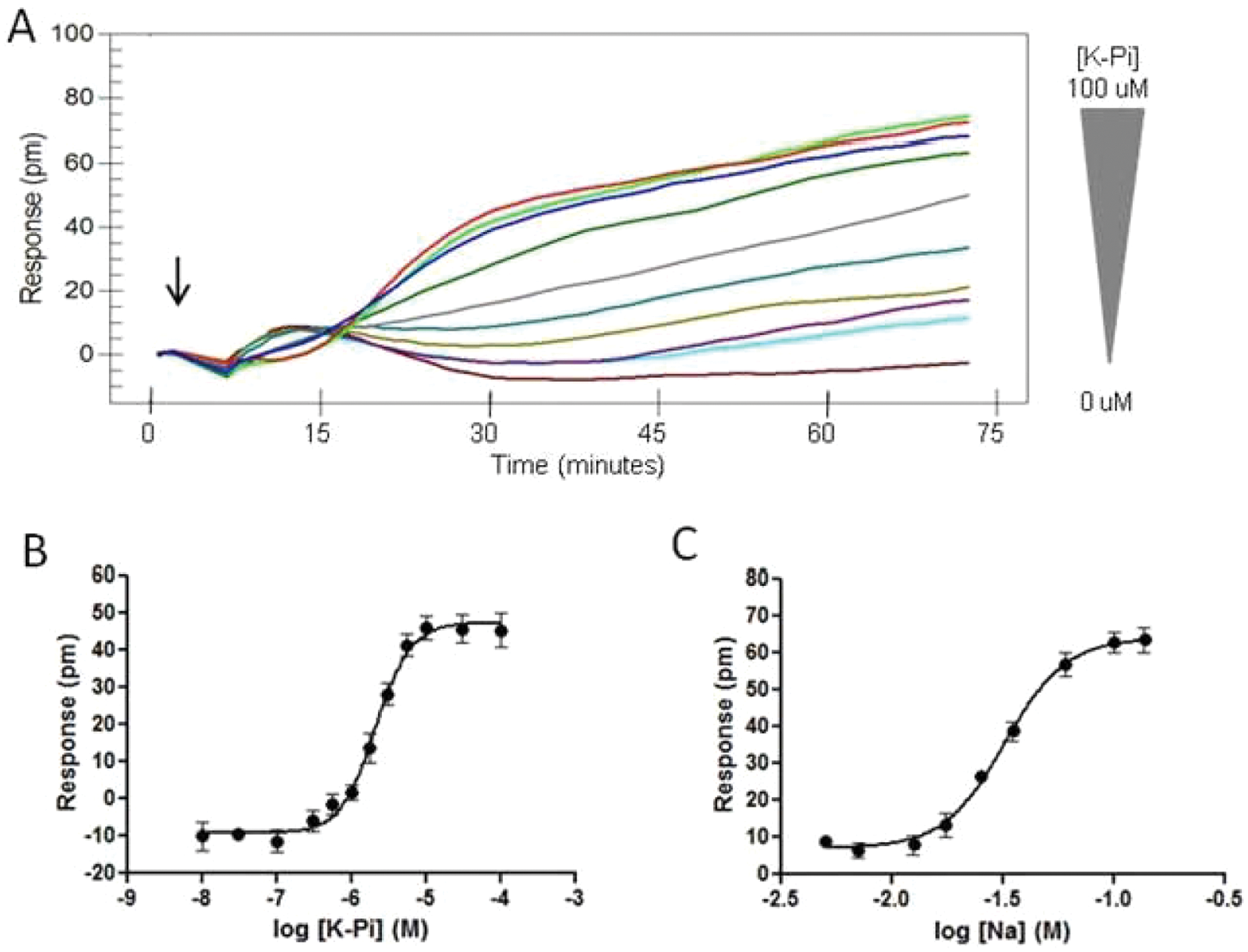
Influence of K-Pi concentration and sodium concentration on the dynamic mass redistribution (DMR) kinetic profile in NaPi-IIb–CHO cells. (
Effects of NaPi-IIb Inhibitors
A known NaPi-IIb inhibitor, JTP-59557 (compound 1), was evaluated for its effect on the K-Pi–induced DMR response on NaPi-IIb–CHO cells. JTP-59557 is a noncompetitive inhibitor of the NaPi-IIb transporter and has a reported IC50 of 0.12 µM. 7 Compound 1 at various concentrations (from 7.5 to 0.0005 µM in a twofold dilution) was preincubated with NaPi-IIb–CHO cells for 1 h during equilibration of the cells in the Epic reader. Compound 1 inhibited the K-Pi–induced DMR response in a concentration-dependent manner with an IC50 of 0.44 µM, which is similar to the IC50 determined by the [33P]-uptake assay in NaPi-IIb–CHO cells ( Table 1 ). Compound 1 alone (in the absence of K-Pi) did not elicit a DMR response (data not shown), therefore confirming that compound 1 is a noncompetitive inhibitor for Pi transport as reported. The fact that compound 1 completely inhibited the K-Pi–induced DMR response further confirms that the observed DMR response is an NaPi-IIb transporter-mediated cellular event. The DMR assay has an average Z factor value of 0.59, showing that it is an excellent assay. 11
Comparison of Dynamic Mass Redistribution (DMR) Measurement to [33P]-Uptake Assay in NaPi-IIb–CHO Cells
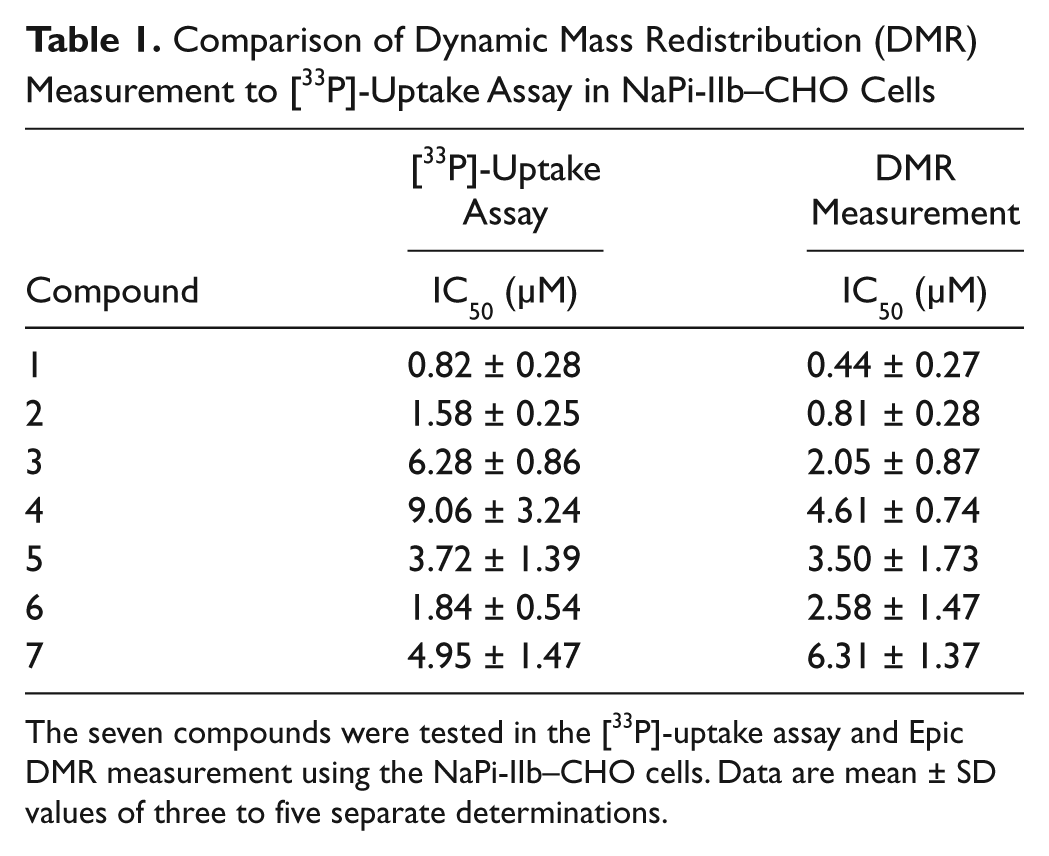
The seven compounds were tested in the [33P]-uptake assay and Epic DMR measurement using the NaPi-IIb–CHO cells. Data are mean ± SD values of three to five separate determinations.
Table 1 compares the IC50 values generated by the DMR measurement to the [33P]-uptake assay for an additional six NaPi-IIb inhibitors identified from a [33P]-uptake screen using the same cell line. For all six compounds, the IC50 values determined from the two assays are in good agreement and within a three-fold difference, suggesting this DMR measurement can be an alternative assay platform for studying activators and inhibitors of NaPi-IIb transporters. DMR responses generated by the activated NaPi-IIb transporters are smaller in magnitude than that observed from ATP. It is not known whether the smaller response is specific to this transporter protein class or a result of a low expression level of NaPi-IIb in this cell line. Further studies using other transporter proteins or developing a cell line expressing a higher level of NaPi-IIb transporters could elucidate the application potential of this label-free DMR technology platform in transporter research.
Conventional methods measuring transporter activity include radiolabeled substrate uptake assays, which require multiple wash steps, and electrophysiology studies using Xenopus oocytes as an expression system.12–15 Recently, an SPA format similar to the one used in this study has been described to measure transporter activity. 16 This SPA format offers one major advantage over the conventional uptake assay format, namely, a homogeneous 384-well assay without a wash step. However, the SPA format retains some drawbacks, including the use of radioisotopes, storage and disposal of radioactive waste, and limitation to low concentrations of radiolabeled substrates due to background issues in the absence of a wash step.
Electrophysiological assays measuring transporter activity make use of the electrogenicity of these targets. These assays, usually performed on oocytes, record the amount of current necessary to clamp the cell membrane at a given applied voltage in the presence or absence of expressed transporter, substrate, or inhibitor. Electrophysiology offers the advantage of a functional, nonradioactive readout with exquisite time resolution. However, its limitations include low throughput and requirements for highly skilled technical personnel as well as specialized equipment. Furthermore, because this assay format is dependent on the electrogenicity of the transporter and measures the net charge being transported, it is not suitable for all transporters. NaPi-IIb is electrogenic and cotransports Na-Pi with the translocation of one net positive charge. 6 However, K-Pi did not elicit detectable currents in the NaPi-IIb–CHO cells using the manual patch-clamp method (data not shown). This lack of detectable current suggests that a high expression level of the transporters, or using an overexpression system such as oocytes, is necessary for studying transporter proteins using electrophysiology methods.
In conclusion, we successfully developed a novel homogeneous label-free assay for the NaPi-IIb transporter using the Epic label-free system and a simple assay protocol. The DMR measurement enables determination of Km values for sodium and phosphate in the millimolar and micromolar range, respectively. This assay does not require substrate labeling and can be performed at any time when the target cell line is in culture. We believe this DMR assay platform could be applicable to other transporter proteins.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.