
Abstract
This study aimed to generate a stable cell line harboring subgenomic dengue virus replicon and a green fluorescent gene (DENV/GFP) for a cell-based model to screen anti-DENV compounds. The gene-encoding envelope protein of DENV-2 was deleted and then replaced with fragments of the GFP gene and a foot-and-mouth-disease virus 2A–derived cleavage site. The human cytomegalovirus immediate early and antisense hepatitis delta virus ribozyme sequences were added at the 5′- and 3′-ends. An internal ribosome entry site and neomycin resistance genes were placed upstream and next to the NS1 gene. The recombinant plasmids were propagated in a mammalian cell line. A stable cell line with the brightest green fluorescent protein and the highest viral protein and RNA expression was selected from six clones. The clone was then examined for effectiveness in an antiviral drug screening assay with compounds isolated from the local plants using two known antiviral agents as controls. Two novel flavones, PMF and TMF, were discovered having DENV-inhibitory properties. The data were validated by a conventional plaque titration assay. The results indicate that this newly developed cell line is efficient for use as a cell-based model for primary screening of anti-DENV compounds.
Introduction
Dengue virus (DENV) is a causative virus of dengue hemorrhage fever and/or dengue shock syndrome. The disease causes a high mortality rate, and the incidence is increasing every year. DENV is transmitted to humans in tropical and subtropical regions by mosquitoes, specifically Aedes aegypti and Aedes albopicus species. Because of the unsuccessfulness of programs to control dengue vectors and lack of effective dengue vaccine, the disease remains a major health problem. 1 There are four dengue serotypes, DENV-1, -2, -3 and -4, that are known, and each is equally capable of causing the disease. Primary infection with one serotype induces an immune response to that specific infectious serotype, whereas a secondary infection with a different serotype enhances the viral pathogenesis leading to severe bleeding, shock, and death. 2 Currently, there is no specific drug for treatment of these syndromes. Only proper fluid replacement and care are the standard regimens in treatment until a person recovers. Therefore, research investigations on searching for effective DENV inhibitors have become important.
The DENV genome consists of 11 000 nucleotides, encoding structural (capsid [C], premembrane [prM], and envelop [E]) and nonstructural (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) proteins. The genome is flanked by 5′NCR (noncoding region) and 3′NCR sequences, which are known to participate in viral replication in viral-infected cells.3,4 Based on the known DENV life cycle, an appropriate inhibitor to DENV has to be an agent that targets the virus at entry, replication, assembly, or release. Several lines of evidence have demonstrated that inhibitors may interact with the virus or the cell surface receptors, 5 block viral replication using NS5 polymerase, 6 inhibit viral assembly and release by acting on certain viral proteases (e.g., NS2B and NS3), 7 and inhibit host proteins such as cellular glycosidase enzyme.8–10 Moreover, discovery of inhibitors has been proposed using structural-based and fragment-based designs.11,12 In these studies, the evaluation method toward anti–drug discovery is dependent on the standard plaque titration assay, which has been confirmed as the most reliable assay. However, this procedure is considered tedious and time-consuming. The newer application of reverse transcriptase polymerase chain reaction (RT-PCR) technology has enhanced the study, but indirect gene amplification requires the appropriate equipment and necessary materials and are the drawbacks. Alternatively, subgenomic replicons have been constructed in susceptible cell lines and used in assessment of viral replication in the cells. By deletion of the DENV-2 (New Guinea C strain) structural gene (C-pr/M-E) and transfection into LLC-MK2 cells, the first DENV replicon containing cells was successfully developed. 13 This original replicon system was capable in viral protein expression, but it was found to replicate inefficiently. To improve DENV replication and translation efficiency, several additional DENV replicon constructs have been subsequently developed, with some modifications. One of them had an insertion of a firefly luciferase reporter gene replacing the deleted structural gene (C-prM-E) of DENV-2 (16681 strain) and addition of a foot-and-mouth-disease virus 2A (FMDV-2A) cleavage site. 14 A similar replicon was then modified to have an additional hepatitis delta virus ribozyme (HDVr) sequence at the 3′-end. 15 These replicons showed good translation at 8 hours posttransfection and good replication efficiency at 48 and 96 hours posttransfection. A further construct was reported to have an addition of an internal ribosome entry site (IRES) gene after the NS5 of DENV-1 Brazillian strain (DV1-BR/90), to amplify the signal of the luciferase activity. 16 The most successful DENV subgenomic replicon development was the modification of these genes into DENV-2 (NGC) and subsequent expression in BHK-2. 17 More recently, inhibitor screening by biochemical analysis of the DENV replication using a fluorescence-based alkaline phosphatase-coupled DENV polymerase has been described. 18 Our study aimed to improve the replicon properties using a color reporter gene (green fluorescent protein [GFP]), with expression that is stronger and more readily observed under the fluorescent microscope. To enhance the screening process for potential dengue inhibitors in DENV-infected cells, the constructed replicon was transfected into a mammalian cell line (BHK-21 cells), and only cells with stable expression were selected for using in the screening assay.
Materials and Methods
Construction of a Subgenomic DENV/GFP Replicon
A plasmid containing full-length DENV-2 strain 16681 (pD2/IC-30P-A) was provided by Dr. Richard M. Kinney. 19 The viral E (envelope) encoding gene, nucleotides 1381–2209, was deleted, apart from nucleotides 937–1380 encoding the transmembrane region of E and nucleotides 2210–2421 encoding the signal sequences of nonstructural 1 (NS1) protein, 20 which were retained. The deleted E was replaced with a cDNA cassette of AcGFP (Aequorea coerulescens GFP) encoding gene (or GFP in this study) amplified from pAcGFP-N1 (Clontech, Mountain View, CA), followed by an FMDV-2A cleavage sequence, using a pair of forward and reverse primers: SphI-AcGFP-F (5′TATAATGCATGCAATGGTGAGCAAGGGCGCCG AGC3′) and FMDV2-BamHI- R(5′ATATATATAGGATCCGGGCCCGGGGTTGGACTCGACGTCT3′). The plasmid was further modified by adding a promoter sequence of the human cytomegalovirus immediate early (CMV-IE) gene upstream of the 5′ terminal of the DENV-2 gene. This sequence was also amplified from pAcGFP-N1, using the primer pair: CMV-F (5′GGCGCGCTAGTT ATTAATAGT3′) and CMV-R (5′ATATCGCCTAGCGGA TCTGA3′). Then, an antisense HDVr sequence was engineered at the 3′ end of the DENV-2 genome by blunt-end ligation. 15 An internal ribosomal entry site and neomycin resistance (IRES-Neo) gene were incorporated upstream to DENV nonstructural genes by sequence amplification from pFB-Neo plasmid (Stratagene, La Jolla, CA), using the pair of forward and reverse primers: BamHI-IRES-F (5′TAGGATCCGCCAGGTTTCCGGGCCCTCAC3′) and Neo-BamHI-R(5’AGCGGATCCGAGAACTCGTCAAG AAGGCGATA G3′). The recombinant plasmid was transformed into Escherichia coli XL-Gold cells (Clontech) and grown in LB medium containing 80 µg/mL ampicillin (Biobasic, Markham, Canada) at 37 °C, overnight. The plasmid DNA was extracted from transformed cells and purified, and the modified genome was sequenced to verify the modifications ( Fig. 1 ).
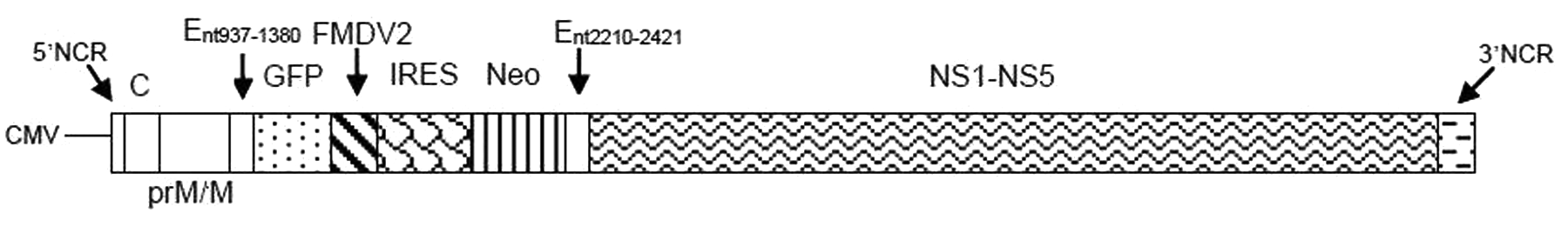
A depicted diagram of the DENV/GFP plasmid. CMV = cytomegalovirus promoter; 5′NCR = 5′ noncoding region; C = dengue capsid gene; prM/M = dengue premembrane/membrane gene; E = partial dengue envelope gene (nucleotides 937–1380 and 2210–2421); GFP = green fluorescent protein gene; FMDV2 = foot-and-mouth-disease virus 2A gene; IRES = internal ribosomal entry site sequences; Neo = neomycin resistance gene; NS1-NS5 = dengue nonstructural 1 to nonstructural 5 gene; HDVr = hepatitis delta virus ribozyme gene; 3′NCR = 3′ noncoding region.
Baby hamster kidney cells (BHK-21 cells) were maintained in Modified Eagle’s Medium (MEM), containing 10% fetal bovine serum (FBS), 0.75 g/L NaHCO3, and 1% penicillin-streptomycin. They were washed twice with DEPC-treated PBS solution, and 2 × 106cells were then resuspended in 0.3 mL of opti-MEM (Invitrogen, Carlsbad, CA) containing 10 µg of the constructed plasmid DNA. The cells were pulsed electrically at 0.5 kV/280 µF, using a BioRad Gene pulser (BioRad, Hercules, CA), and the electroporated cells were then plated into 24-well plates and grown in MEM containing 10% FBS for 24 h. Fresh medium, containing 700 µg/mL of G418 (Biobasic) as a selective marker for neomycin-resistant cells, was added on the following day. The medium was replaced every 3 to 4 d. After 2 to 4 wk, clones of neomycin-resistant DENV/GFP replicon–containing cells were collected and further cultured for several passages in the presence of G418.
Characterization of Stable Cell Lines Containing a DENV/GFP Replicon
GFP expression and flow cytometry analysis
The clonally derived DENV/GFP replicon–containing cells at passages 10, 20, and 30 were examined under the fluorescent microscope (Olympus, Center Valley, PA). The cells were harvested by trypsinization and then resuspended in fresh media at 4 °C. The positive GFP expression in each cell was further analyzed by flow cytometry. The mean fluorescent intensity (MFI) from each clone, read at 488 nm, was analyzed by a FACScanto flow cytometer (BD Biosciences, Franklin Lakes, NJ) at the Central Instrument Facilities of Mahidol University. Stable cells expressing DENV/GFP were expanded, collected, and stored in 10% DMSO and 50% FBS at −80 °C.
Quantitative real-time PCR for DENV-2 RNA replication
RNA was extracted from 1 × 106 DENV/GFP replicon containing BHK-21 cells after 48 h of culture using an RNeasy kit (Qiagen, Germantown, MD). Residual DNA was removed using an RNase-free DNase kit (Qiagen), and the total RNA was quantified by spectrophotometry. Negative sense cDNA synthesis was performed using RevertAid M-MuLV Reverse Transcriptase (Fermentas, Burlington, Ontario, Canada) with 1 µg (2% of the total extracted RNA). RT was performed using a specific primer sequence to DENV-2 3′NCR (5′TGCAGTCATATCGAGCGCTGCCATAG CTTGAGTAAACT ATGC3′), at 42 °C for 1 h, and the reaction was then terminated by heating at 70 °C for 10 min. The reaction product was purified using a PCR purification kit (Qiagen). Real-time PCR reaction was performed using SYBR Green I (BioRad), in a BioRad iQ5, with the primer pair (forward 5′GAAGTCAGGCCTCATAA A3′ and reverse 5′TCTCCTCTAACCGCTAGTCCACTA3′). Briefly, samples were assayed in 25 µL reaction mixture, containing 1 µL of total 20 µL of the cDNA, 2.5 µL 10× High Fidelity Reaction buffer, 0.2 µL 25 µM dNTPs, 1.25 µL 10 µM each primer, 0.75 µL SYBR Green I stock solution (1:1000 dilution in DEPC-treated water), and 0.2 µL High Fidelity PCR enzyme mix (Fermentus). The thermocycler profile for SYBR Green I–based PCR consisted of 3 min of Taq polymerase activation at 94 °C, followed by 45 cycles of PCR at 95 °C for 30 s (denaturing), 55 °C for 30 s (annealing), and 72 °C for 1 min (extension). Melting curve analysis was performed with software from BioRad to verify the correct product by its specific melting temperature (Tm). Melting curve analysis consisted of a denaturing step at 95 °C for 1 min, after which the temperature was lowered to 60 °C for 30 s, and then 35 cycles of incubation were performed in which the temperature was increased to 95 °C at a rate of 1 °C/30 s/cycles with continuous reading of fluorescence. Results were analyzed using BioRad iQ5 standard version 2.0. A standard curve was plotted in every experiment using 10-fold serial diluted standards with 1.16 to 1.16 × 1010 copies of DNA of infectious clone plasmid (pD2/IC-30P-A). Real time-PCR was performed as described above, except using specific primers for DENV-2 with the known size of the plasmid (13.6 kb). The copy numbers of the DENV-2/GFP replicon were calculated according to the standard curve in the real-time PCR for comparison. The total copy number of replicon per million cells equals the copy number per reaction ×1000.
Western blot analysis for DENV protein expression
The stable expressed DENV/GFP replicon cells were cultured in 6-well plates at a density of 1 × 105 cells/mL per well for 48 h. The cells were lysed in cold buffer (pH 7.5) containing 25 mM Tris-HCl, 150 mM NaCl, 1% NP-40 (nondedet-P-40), 1% sodium deoxycholate, and 0.1% SDS containing a protease inhibitor cocktail (#539134, Calbiochem). Cell lysate was centrifuged at 14 000 g for 20 min at 4 °C. The supernatant was determined for total protein concentration using a Bradford assay (BioRad). The proteins (30 µg) were separated by 12.5% SDS–polyacrylamide gel electrophoresis at 100 V in running buffer and then transferred to a 0.45 µm nitrocellulose membrane (Amersham, Little Chalfont, UK), using 30 V at 4 °C overnight in the transfer buffer. The membrane was blocked with nonfat dry milk (5% w/v) in Tris-buffered saline (TBS) containing 0.1% Tween-20 for 2 h at room temperature. It was then incubated with mouse monoclonal anti-mouse NS-1 antibody, kindly provided by Dr. Chunya Puttikhunt, Siriraj Institute of Molecular Biotechnology Research 21 (1:1000 dilution in TBS), overnight at 4 °C. After washing, the membranes were incubated with goat antimouse conjugated with HRP antibody (Santa Cruz Biotechnology, Santa Cruz, CA). The enzyme reaction was developed using a DAB substrate, and the color was observed within 10 min.
Assessment of the Established Stably Cell Line for DENV Inhibitors Assay
To assess for stability of the selected stable cell line, BHK-21 cells harboring DENV/GFP replicons from passages 10, 20, and 30 were grown in 10% MEM containing 700 µg/mL of G418 in 24-well plates at 37 °C for 24 h (70%–80% confluent). The cells were then incubated with varied concentrations (0.1–100 µM) of commercial antiviral compounds, namely, mycophenolic acid or ribavirin (Sigma), for 48 h. The cells were closely monitored for the expression of GFP under the fluorescence microscope. The MFI level in each GFP-positive cell was analyzed by flow cytometry as above. The effective inhibitory concentration that reduced 50% MFI level in GFP-positive cells (EC50) was calculated from a dose-response curve.
Using of the Established Stably Cell Line for Anti-DENV Compound Screening Assay
Plant compounds preparation
Two novel phenolic compounds expected to have antiviral properties were obtained from Dr. Patoomratana Tuchinda, Department of Chemistry, Faculty of Science, Mahidol University, and were extracted from leaves and twigs of Gardenia collinsae (Craib). The chemical structures of the compounds, determined from chloroform fractioning, showed both were derivatives of flavones: 5,3′-dihydroxy-6,7,4′,5′-tetramethoxyflavone (TMF) and 5′hydroxyl-6,7,3′,4′,5′-pentamethoxyflavone (PMF; Fig. 2 ). The compounds were dissolved in serum-free DMEM (Sigma) and kept as stock solutions (10 mg/mL) at 4 °C. Working solutions were freshly prepared from the stocks by dilution in free-serum medium containing DMSO (Sigma) and were filtered through 0.22-µm membranes to make final concentrations of DMSO less than 0.5% (v/v). Cytoxicity of the test compounds PMF and TMF to BHK-21 cells were determined using a standard MTT assay (Roche Applied Science, Penzberg, Germany), according to the manufacturer’s instructions. Briefly, BHK-21 cells were seeded into 96-well plates (1 × 104 cells per well) and incubated at 37 °C in a 5% CO2 incubator for 48 h. The culture medium was removed, and the cells were treated with 100 µL of MEM-containing test compounds at various concentrations. After 48 h, the cells were washed with PBS and 10% 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide was added. The color reactions were measured by an automatic VersaMax microplate reader (Sunnyvale, CA) at a test wavelength of 570 nm and a reference wavelength of 620 nm. The concentration of compound that decreased 50% cell viability (CC50 values) was determined and then compared with the untreated (DMSO treated) controls.
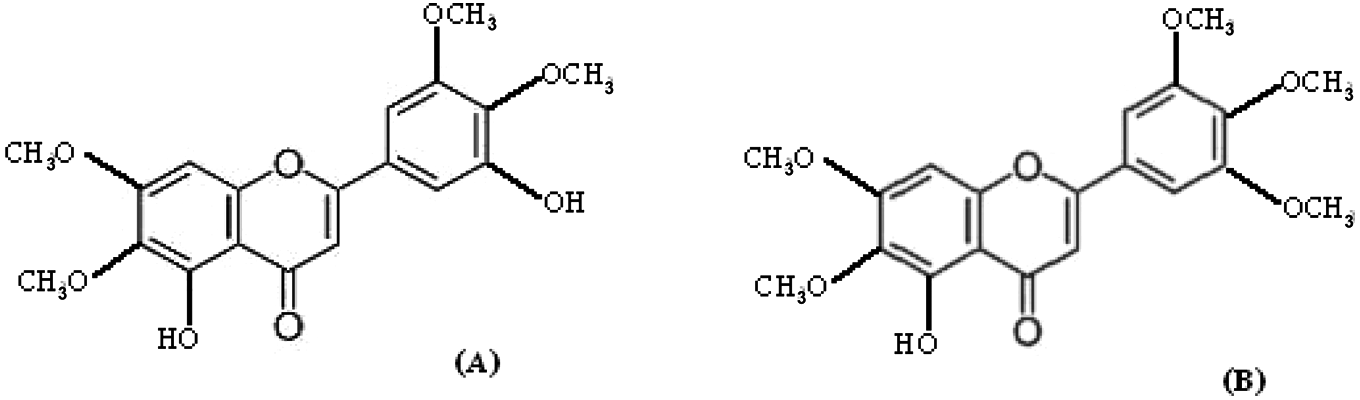
Chemical structures of (A) 5,3′-dihydroxy-6,7,4′,5′-tetramethoxyflavone (TMF) and (B) 5-hydroxy-6,7,3′,4′,5′-pentamethoxyflavone (PMF).
To determine the effective concentration (EC50) of the potential antiviral compounds, the established BHK-21 cells harboring DENV/GFP replicon were grown in 24-well plates at 37 °C for 24 h (70%–80% confluence) before incubation with various concentrations of the antiviral drugs and the test compounds, mycophenolic acid (0.025–50 µM), ribavirin (1–200 µM), PMF (1.25–20 µg/mL), or TMF (1.25–20 µg/mL), for 48 h. The MFI level in each GFP-positive cell was analyzed by flow cytometry, as above.
To screen the potential antiviral compounds using the established BHK-21 cells harboring DENV/GFP replicon, the stable cells expressing DENV/GFP were grown to confluence (100%) before incubation with the EC50 doses of the test compounds (mycophenolic acid ribavirin), PMF or TMF, in 24-well plates at 37 °C for 48 h. The MFI levels, and the quantitative viral RNA expression in the treated cells, were determined as above.
Data validation of viral inhibition by conventional plaque titration assay
To validate the results, antiviral activity was performed by conventional or standard plaque titration assay as described previously. 22 Briefly, normal BHK-21 cells (2 × 106cells) without a replicon were grown in 6-well plates to 70% to 80% confluence. Cells were inoculated with DENV-2 16681 virus suspension at multiplicity of infection (moi) of 0.01 (0.2 mL) and left for viral adsorption at 37 °C for 90 min. The medium was carefully aspirated, and the cells were treated with 2 mL of MEM, containing an effective concentration (EC50) of mycophenolic acid, ribavirin, or test compound at 37 °C in a 5% CO2 incubator. Treatment was performed in triplicate for each concentration. After 48 h incubation, the cells were first overlaid with a 2% stock of Seakem agarose mixed with an equal volume of 2× nutrient solution. The plates were incubated for 7 d at 37 °C in a 5% CO2 incubator, after which they were overlaid with a 2% stock of Seakem agarose mixed with an equal volume of 2× nutrient solution containing 1% neutral red. Plaque production was observed and then counted. Percentage plaque reduction was calculated from the number of plaque formation in the control minus the number of plaque formation by each compound, divided by the number of plaque in the control, and multiplied by 100 (N = 3).
Statistical Analysis
Except for Western blot analysis, each experiment was performed in triplicate. The results were analyzed using a one-way analysis of variance, followed by a Tukey post hoc test. All values were presented as mean ± the standard deviation, and differences were considered significant at p < 0.05.
Results
Clonal Selection of BHK-21 Cell Line Stably Expressed DENV/GFP Replicon
Sequence analysis of DNA from the plasmid construction determined the correct position of genes in the designed subgenomic replicon ( Fig. 1 ). During the transient transfection stage of the plasmid in BHK-21 cells, bright fluorescent signal from the GFP expression was observed at 6 h posttransfection, and the signal progressively increased throughout 72 h of observation (data not shown). After cells were grown in neomycin-selective medium for 2 to 4 wk, various clonally expressed cells were derived, of which six clones (#1, #9, #14, #17, #22, and #28) were positive.
To evaluate for stability and efficiency of the clonally derived cells, the expression of GFP and DENV RNA production in each clonally derived cell line was analyzed. With the exception of clones #9 and #14, the MFI levels in individual treated cells from clones #1, #17, #22, and #28 were constant from passages 10 to 30 ( Fig. 3A ). Real-time PCR analyses of DENV RNA replication at passages 10, 20, and 30 also showed decreased amounts of RNA syntheses in clones #9 and #14, correlating to their GFP expression ( Fig. 3B ). Among all clones, clone #17 produced the highest level of RNA replication. To evaluate the capability of the replicon in viral protein expression, Western blot analysis of the DENV NS1 protein was performed, and the result showed that cells derived from the six selected clones of DENV/GFP replicon at passage 10 expressed different levels of NS1 protein ( Fig. 4 ). Among them, clone #17 DENV/GFP replicon expressed the highest level of NS1 protein relative to the non-replicon–containing cells after infection with the same amount of RNA (moi 0.01) from the wild-type DENV-2 16681 infectious clone. The results indicated that clone #17 was the most efficient in RNA replication and viral protein expression.
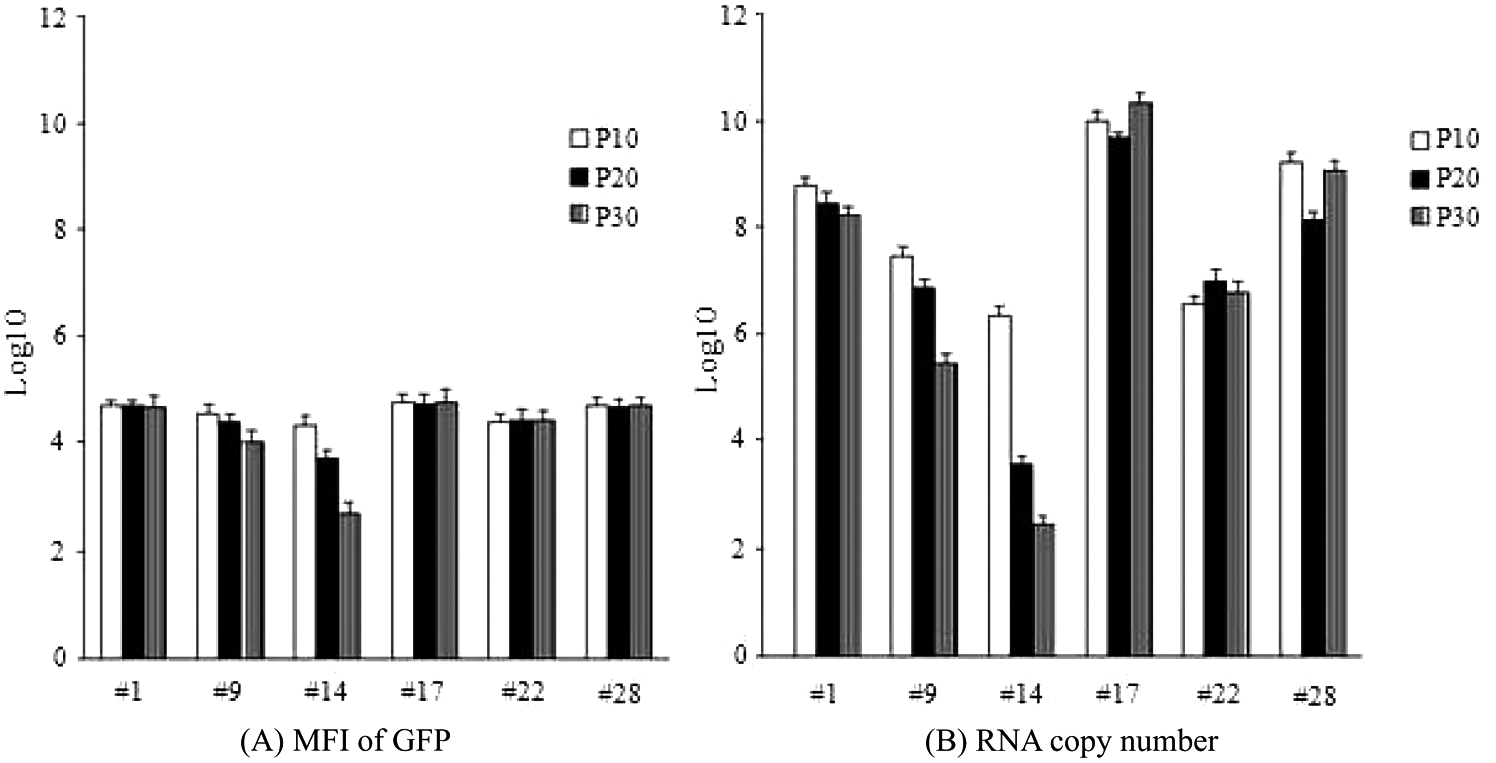
Histograms showing green fluorescent protein (GFP) expression and viral RNA replication of six clonally derived DENV/GFP replicon–containing cells at passage 10, 20, and 30 (P10, P20, and P30). (A) Mean fluorescence intensity (MFI) of GFP of each clone, with the mean recorded as log10 of the arbitrary unit. (B) Quantitative real-time PCR measurement of viral RNA replication, recorded as mean of log10 of RNA copy number. Bar lines represent the standard deviation (SD) of the means of these experiments.
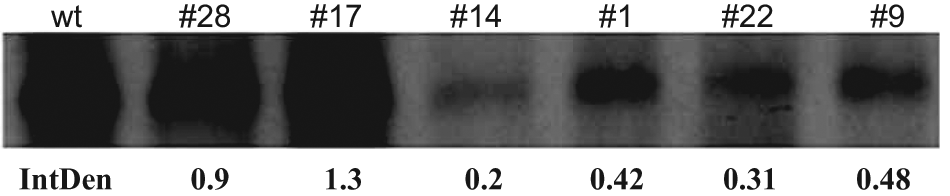
Western blot analysis of DENV-2 NS1 protein expression in six clonally derived DENV/GFP replicon–containing cells at passage 10. The translation efficiency was compared with the non–replicon-containing cells infected with the same amount of RNA (multiplicity of infection 0.01) obtained from the wild-type (wt) DEN-2 16681 infectious clone. Protein band density (IntDen) was determined by ImageJ.
A further study was performed to evaluate the consistency and efficiency of GFP expression in the selected stable cell line, BHK-21 cells harbored the DENV/GFP replicon #17. As shown in Table 1 , treatment of the cells from passages 10, 20, and 30, with known antiviral compounds mycophenolic acid and ribavirin, resulted in a nonsignificant difference of MFI level and their effective concentration. This indicated that #17 DENV/GFP replicon–containing cells was stable and appropriate to be used as a model for evaluation of dengue inhibitors. Thus, clone #17 DENV/GFP replicon–containing cells was chosen for further use as a model for screening of DENV inhibitors.
Evaluation for Consistency of GFP Expression in the Stably Established #17 DENV/GFP Replicon–Containing Cells after Treatment with Various Concentrations of Mycophenolic Acid and Ribavirin (0.1–100 µM) at Passage 10, 20, and 30 (P10, P20, and P30) a
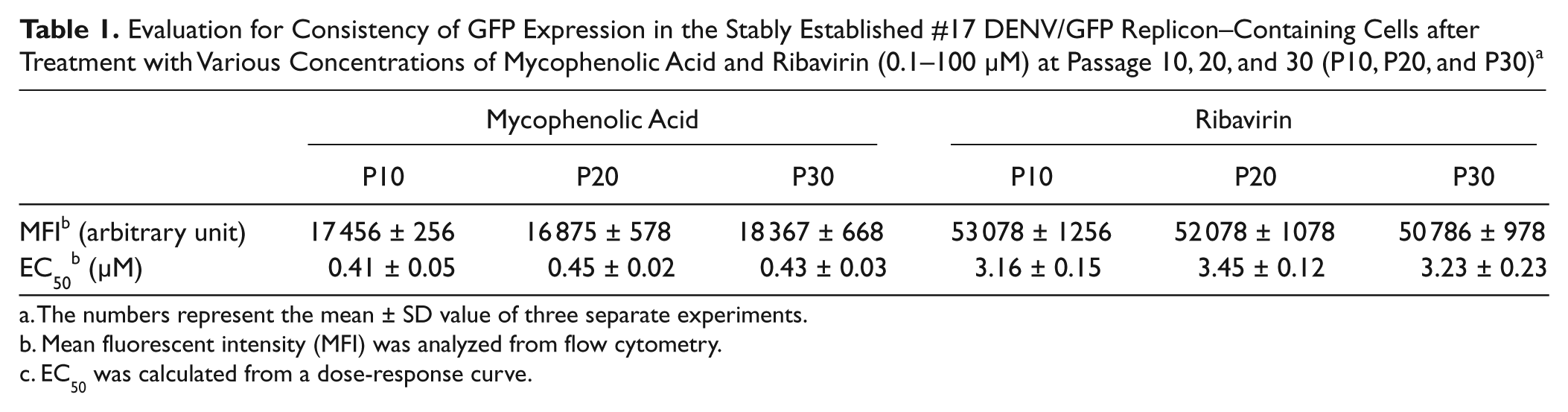
The numbers represent the mean ± SD value of three separate experiments.
Mean fluorescent intensity (MFI) was analyzed from flow cytometry.
EC50 was calculated from a dose-response curve.
Screening Assay for DENV Inhibitors Using the Established Stable Cell Line
To test the efficacy of the established stable cell line, we screened several pure compounds from plant extracts that we had obtained from the chemistry department. Because flavonoid derivatives have previously shown broad spectra of biological activities, we evaluated two novel flavones (i.e., PMF and TMF) for potential anti-DENV effects in the #17 stable cell line, using mycophenolic acid and ribavirin as positive controls. First, the stable cells expressing DENV/GFP were grown to subconfluence (70%–80%) before treatment, with varied concentrations of the antiviral drugs and the test compounds, in 24-well plates at 37 °C for 48 h. The EC50s of mycophenolic acid and ribavirin, determined from the dose-response curves ( Fig. 5A , B ), were in the same ranges as the EC50s obtained in Table 1 . The EC50s of PMF and TMF were at 4.47 µg/mL and 5.62 µg/mL respectively ( Fig. 5C , D ). Then, the EC50 doses of the test compounds were incubated with the newly established stable cells expressing DENV/GFP that had been grown to confluence (100%) in 24-well plates at 37 °C for 48 h. After treatment with PMF and TMF compounds, there was a decrease of more than 50% in MFI and more than 2 log RNA copies in the treated cells. The MFI level ( Fig. 6A ) was decreased significantly (p < 0.05), in a pattern that correlated with the significant decreased (p < 0.05) DEN-2 RNA expression ( Fig. 6B ). The cytotoxic concentrations (CC50) of PMF and TMF to normal BHK-21 cells were 4.22 µg/mL and 25.03 µg/mL, at 48 h posttreatment, respectively. The selective index ratio between CC50 and EC50 of PMF and TMF was 0.94 and 4.45, respectively ( Table 2 ).
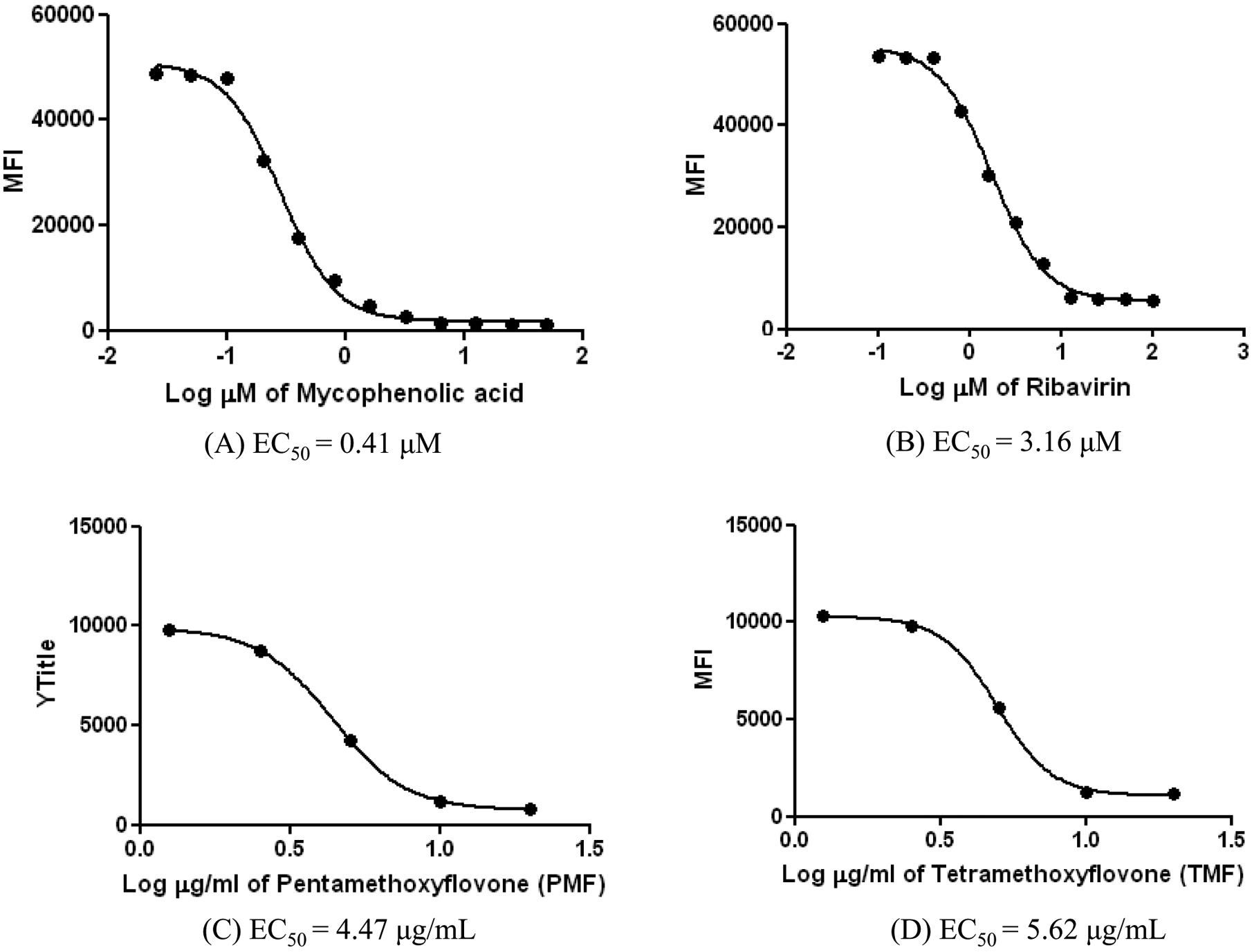
The dose-response curve analysis of the known antiviral drugs and the potential anti-DENV compounds as determined from mean fluorescent intensity (MFI) levels in the established stable cell line, DENV/GFP replicon #17–containing cells. The concentration of the compounds varied from 0.025 to 50 µM for mycophenolic acid, 1 to 200 µM for ribavirin, and 1.25 to 20 µM for PMF and TMF. MFI is shown in arbitrary units. EC50 = effective concentration.
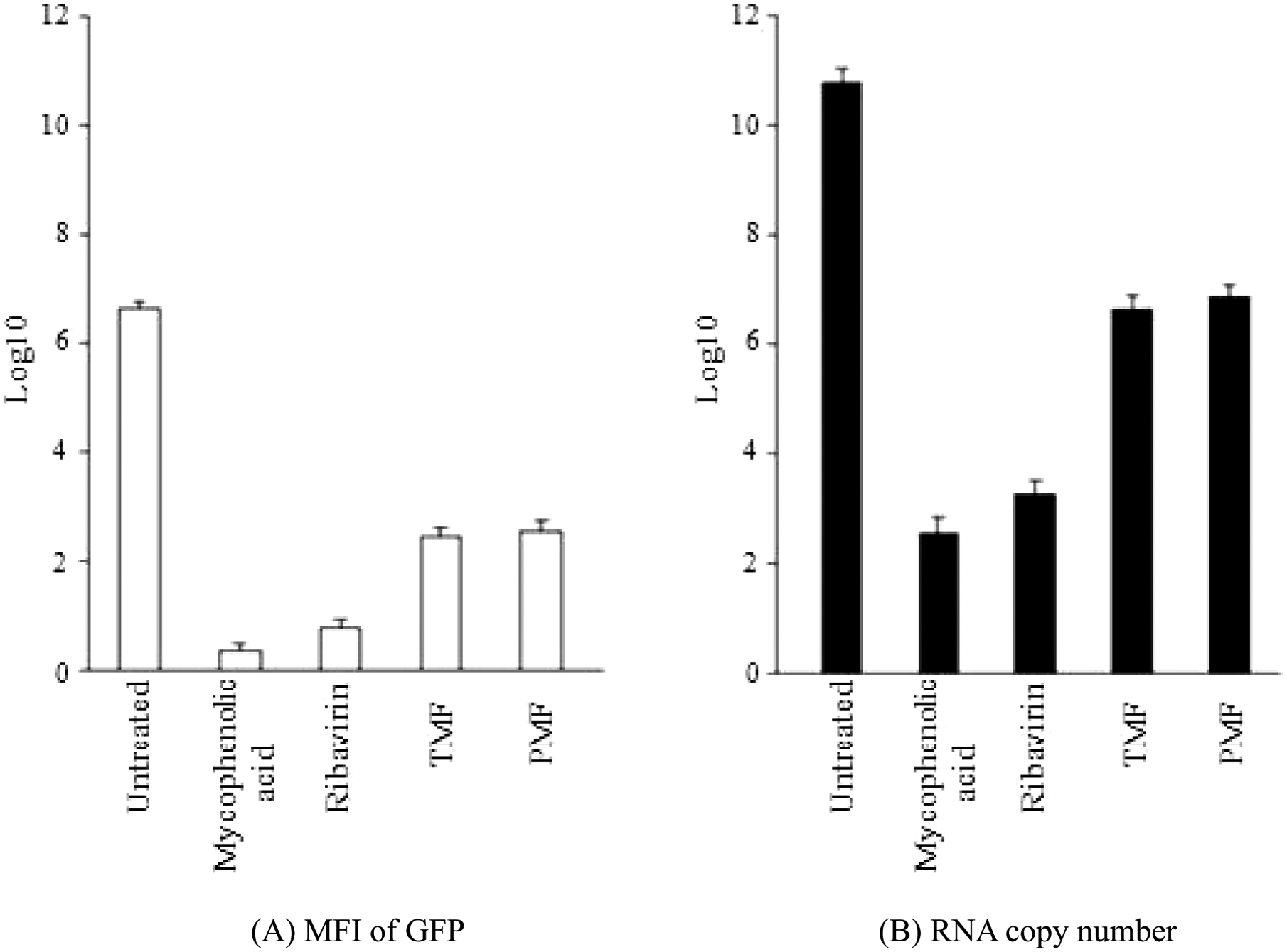
Analysis of green fluorescent protein (GFP) expression and DENV RNA in DENV/GFP replicon #17–containing cells after treatment with the EC50 concentrations of mycophenolic acid, ribavirin, PMF, and TMF. (A) Mean fluorescence intensity (MFI) from flow cytometry analysis, with the mean recorded as log10 of the arbitrary unit. (B) RNA copy number from the quantitative real-time PCR assay, recorded as the mean of log10 of the RNA copy number. Bar lines represent the standard deviation (SD) of the means of these experiments.
The Compounds’ Effectiveness Determined from a Selective Index Value and Conventional Plaque Titration Assay a
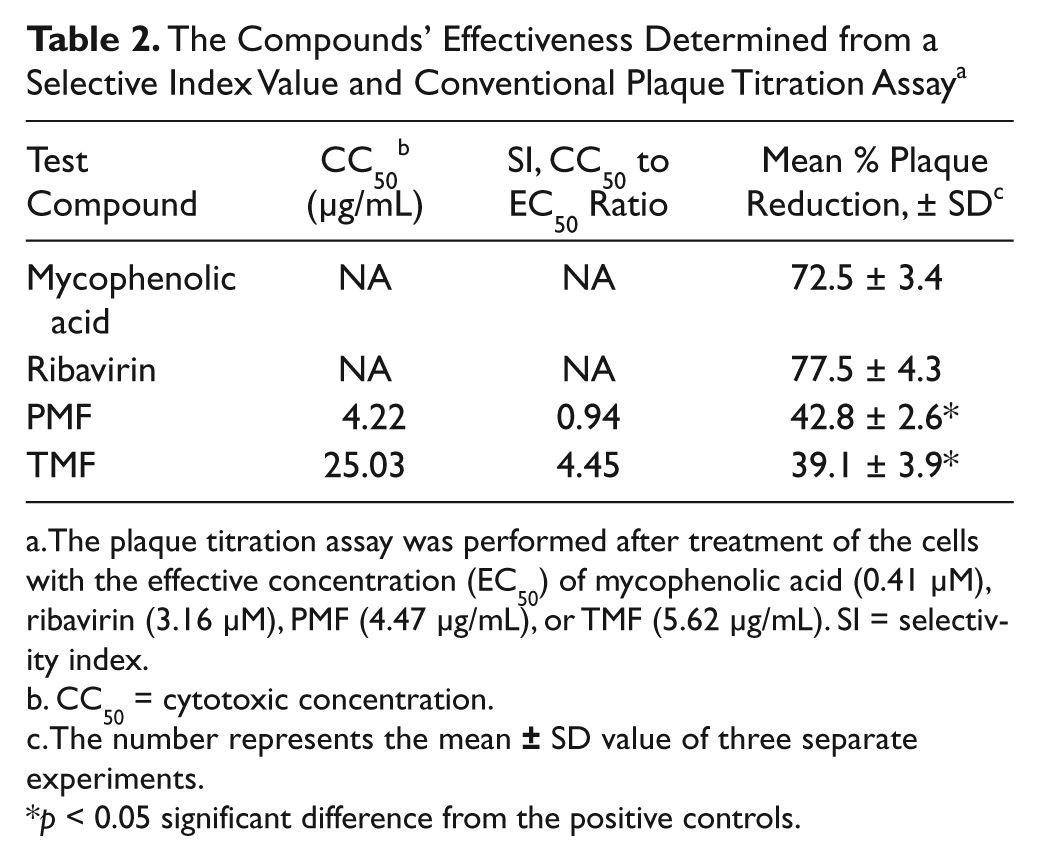
The plaque titration assay was performed after treatment of the cells with the effective concentration (EC50) of mycophenolic acid (0.41 µM), ribavirin (3.16 µM), PMF (4.47 µg/mL), or TMF (5.62 µg/mL). SI = selectivity index.
CC50 = cytotoxic concentration.
The number represents the mean
p < 0.05 significant difference from the positive controls.
Validation of DENV inhibition effects was performed in DENV-2–infected BHK-21 cells by treatment with the EC50s of the test compounds. The broad-spectrum antiviral compounds (i.e., mycophenolic acid and ribavirin) used as positive controls reduced virus production in the viral infected cells by approximately 75%, whereas both PMF and TMF reduced virus production by approximately 40% ( Table 2 ).
Discussion
In this study, stable BHK-21 cell lines that co-expressed DENV and GFPs were constructed successfully. The replicon used in the expression was constructed by placing a GFP gene at the partial deletion of DENV E sequences and also by adding some necessary genes to obtain appropriate stable cells for the rapid screening of dengue inhibitors. Deletion of the dengue E encoded gene has previously shown no serious effect on the replication capability of the replicon, and the GFP insertion revealed no perturbation effect to the viral translation process (our unpublished data). Because the envelope protein of DENV determines the viral serotype, removal of the E could be representative of most types of dengue replicon in cellular model assays for screening dengue inhibitors. Our study employed an AcGFP gene as the reporter marker instead of the original EGFP gene because the coding sequence of the AcGFP had previously been modified to a silent mutation causing enhanced GFP expression but maintained stability in mammalian cells. 23 The design was different from a previously reported DENV-2 NGC strain replicon constructed in LLC-MK2 cells, in which GFP replaced the whole structural gene, resulting in a low yield of replicon-expressing cells, and there was no stable cell line generated. 13 The construct in this study was considered a large subgenomic replicon containing only a minor part of the E gene of the wild-type DENV-2 16681 strain. Further replicon developments include addition of a CMV promoter sequence upstream of the 5′-end of DENV gene, insertion of a FMDV-2A cleavage site 14 immediately after the GFP, addition of an IRES gene 16 instead of a pac gene (resistance marker) 24 upstream from the NS protein, and addition of an HDVr sequence at the 3′-end. 15 These applications followed the others’ previous studies, resulting in a highly significant improvement of DENV replication and translation efficiency. Insertion of EMCV-IRES in the constructed plasmid has been proved to drive sufficient transcription and translation of DENV-2 at 24 to 48 h and 2 to 8 h, respectively. 25 We also found that CMV-IE insertion at the 5′NCR was involved in the initial plasmid transcription and translation, but the increased RNA at 24 to 72 h was mainly due to autoreplication of the constructed replicon (our parallel study, data not shown). The results in this study indicated that the replicon was self-replicating and maintained continuously in the cultured BHK-21 cells. The selected subgenomic replicon (DENV/GFP #17) was developed to express consistently in the cells by insertion of the IRES and neomycin resistance genes upstream to the nonstructural genes of DENV. They are known to drive adequate translation of proteins and can act as selection markers. 26 Compared with previous studies, this novel replicon was engineered to have highly efficient RNA replication and protein translation. Evaluation of the established replicon demonstrated high GFP expression levels in cells that could be detected 6 h posttransfection and with a signal that increased continuously throughout 72 h. The intracellular viral protein production also confirmed complete viral translation. These results showed an amplification of the signal of a GFP-based replicon superior to that of the stably expressed DENV-2 NGC replicon previously established in BHK-21 cells using a synthetic luciferase gene (from Promega). 17 Although the original luciferase gene had been optimized to promote reporter translation and replication efficiency, the translation was detected at 8 h and replication was detected at 48 h posttransfection. 17 A drawback of using a luciferase gene is that the assay requires an additional enzymatic process, whereas no enzymatic process is required when using the GFP gene. Another benefit of GFP is that the green fluorescent illumination can be observed directly under a fluorescence microscope, providing a simple and rapid screening of a large number of compounds in a very short time. In addition, after protein translation, the GFP is cleaved at the FMDV-2A site and floats into the nucleus without disruption to cell function. This system has provided us a reliable microscopic study of treated cells in an evaluation of cytotoxic morphological changes in real time.
The initial evaluation of the best-performing cell line, containing the selected constructed replicon, with commercially available viral inhibitors indicated that the cell line was stable, allowing it to be used as a suitable tool for primary screening of potential DENV inhibitors. In the screening assay, our novel replicon-containing cells were successfully applied to screen several compounds from plant extracts by comparing their activities to two known antiviral drugs. The GFP expression analysis was performed in only live cells. The EC50 (determined from MFI level) of each pure compound varied, suggesting a different degree of inhibition. The effectiveness of each of the two established broad-spectrum antiviral drugs (i.e., mycophenolic acid and ribavirin) was found to be in a similar range to the EC50s determined previously using a different DENV-2 replicon system. 17 The EC50 results in this study suggested that mycophenolic acid performed a stronger inhibitory action than ribavirin did. It is known that mycophenolic acid powerfully inhibits DENV by preventing replication of both positive- and negative-strand viral RNA,27,28 and ribavirin effectively suppresses viral enzyme RNA-dependent RNA polymerase (RdRp) of several RNA viruses. 29 The plaque titration assay of the two antiviral drugs also confirmed that this cell-based assay was very efficient in identifying new DENV inhibitors. Among the compounds that showed positive results, both PMF and TMF compounds had potent DENV inhibitor properties. As revealed in Table 2 , the anti-DENV effect of PMF and TMF was less than 50% efficacy relative to the mean efficacy of known antiviral drugs tested, which are mycophenoic acid and ribavirin (72.5 and 77.5, respectively). To rule out the problem of the lose-of-signal endpoint that creates a false-positive from drug-mediated cytotoxicity, our counter study by an in vitro system of cytotoxicity assay and morphological observation has confirmed that both compounds did not affect cell survival. Thus, PMF and TMF are our newly discovered compounds that might be the significant lead compounds to be further developed to anti-DENV drugs. The selectivity index of 0.94 for PMF and 4.45 for TMF may suggest the potential compound toxicity, but this needs to be further clarified in vivo. Although a certain degree of replication and translation inhibition effect of the inhibitors could be demonstrated, this assay could not clearly make a distinction between effects on replication or translation because both were interrelated in our system. More detailed studies on the mechanism of inhibition are required. These include inhibition of both positive- and negative-strand RNA replication, as reported with mycophenolic acid,27,28 or inhibition of viral enzyme RdRp, as reported with ribavirin.28,29
In conclusion, our established cell-based model should have benefits for the primary screening of natural compounds, such as plant extracts and synthetic inhibitors, to detect dengue inhibitors. The cell-based model was created specifically to DENV, and it is highly efficient in providing an evaluation within a few days. Mass screening is also possible, providing a low-cost and rapid screening assay. This method will facilitate new drug discovery against DENV.
Footnotes
Acknowledgements
We Thank Dr Peter J Hanna for his help in English correction of the manuscript.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
This study was funded by Mahidol University, the Office of the Higher Education Commission, and Mahidol University under the National Research Universities Initiative and partly by the Thailand BIOTEC/NSTDA. V.L. is supported by the Thailand Research Fund.