
Abstract
dTDP-L-rhamnose as a sugar donor provides L-rhamnosyl residue in the synthesis of disaccharide linker (D-N-acetylglucosamine-L-rhamnose), the key structure of the Mycobacterium tuberculosis cell wall. Four enzymes are involved in the formation of dTDP-L-rhamnose and D-glucose-1-phosphate thymidylyltransferase (RmlA) catalyzes the first step of D-glucose-1-phosphate and dTTP to dTDP-D-glucose and PPi. The previous studies on RmlA essentiality proved RmlA as a potential target for antituberculosis drugs. However, there has not been a suitable assay for RmlA to screen inhibitors currently. In this study, the authors reported a microtiter plate–based colorimetric assay for RmlA enzyme activity. Using this assay, the kinetic properties of M. tuberculosis RmlA including initial velocity, optimal temperature, optimal pH, the effect of Mg2+, and kinetic parameters were determined. The establishment of the accurate and rapid colorimetric assay and kinetic analysis of M. tuberculosis RmlA will facilitate high-throughput screening of RmlA inhibitors.
Introduction
Tuberculosis (TB) is still a global problem that we cannot ignore. One-third of humanity, more than 2 billion people, carry Mycobacterium tuberculosis that causes TB, and each year 2 million people die of this disease. Multidrug-resistant and extensively drug-resistant TB cases are increasing fast, especially in the low-income countries. 1 There is an urgent need for identifying more targets to develop new anti-TB drugs.
M. tuberculosis has a unique impermeable cell wall, which contains many potential drug targets. The cell wall core is composed of three covalently attached layers: peptidoglycan, arabinogalactan, and mycolic acids. A D-N-acetylglucosamine-L-rhamnose disaccharide as a linker connects arabinogalactan to peptidoglycan and thus is critical to the structural integrity of M. tuberculosis cell wall. 2 The L-rhamnose of the disaccharide is from a donor, dTDP-L-rhamnose. Four enzymes (RmlA-D) are involved in the dTDP-L-rhamnose synthetic pathway, and D-glucose-1-phosphate thymidylyltransferase (RmlA) catalyzes the first step of dTTP and D-glucose-1-phophate (D-Glc-1-P) to dTDP-D-glucose (dTDP-D-Glc) and PPi. 3 Insertional mutagenesis studies using transposon site hybridization methodology reported that M. tuberculosis rmlA was an essential gene. 4 In our previous studies, the rmlA gene was knocked out from the genome of Mycobacterium smegmatis mc2155 in the presence of M. tuberculosis rmlA on a temperature-sensitive plasmid, and the mutant cells did not grow at nonpermissive temperature, which also confirmed the essentiality of RmlA for mycobacterial growth. 5 In addition, neither L-rhamnose nor the genes encoding RmlA-D have been identified in humans so far. 6 These results all supported RmlA as a potential target for anti-TB drug development.
RmlA is also present in other Gram-negative and Gram-positive bacteria, in which L-rhamnose is involved in the biosynthesis of some extracellular components, such as O-antigen or glycoprotein.7–9 Traditionally, the D-glucose-1-phosphate thymidylyltransferase activity of RmlA was detected through high-performance liquid chromatography (HPLC)7,8 or capillary electrophoresis (CE) 10 analysis. However, HPLC and CE methods consumed time and needed more amounts of substrates and enzymes, and thus were not suitable for high-throughput screening of inhibitors. Some other methods such as an NADH oxidation-coupled assay 11 and pHBH assay, 12 which detected PPi and D-Glc-1-P, respectively, could also be used to detect RmlA activity. However, both assays had complicated procedures. In the pHBH assay, the reaction mixture needed to be heated at 100 °C for 5 min and subsequently transferred to another clear-bottomed 96-well plate to measure its absorbance, and in the NADH oxidation-coupled assay, four enzymes (pyrophosphate, aldolase, triosephosphate isomerase, and glycerophosphate dehydrogenase) needed to be coupled to detect PPi finally.
In this study, a simple and rapid colorimetric assay was developed for RmlA enzyme activity. The kinetic properties of M. tuberculosis RmlA including the optimal reaction temperature, pH, and concentration of Mg2+, as well as Km and Vmax values of two substrates (dTTP and D-Glc-1-P), were also measured using the assay.
Materials and Methods
Bacterial strains, plasmids, and growth conditions
pET16b (Novagen) was selected as an expression vector. Escherichia coli NovaBlue and Escherichia coli BL21(DE3) (Novagen) were used as hosts for pET16b-Mtb rmlA construction and RmlA protein expression, respectively. These strains were all grown in LB medium. Ampicillin (Amp) was used at the final concentration of 100 µg/ml.
RmlA protein expression and purification
The M. tuberculosis rmlA gene was obtained from previously constructed pMD-rmlA plasmid 5 and inserted into the NdeI and XhoI sites of pET16b vector, yielding an expression plasmid pET16b-Mtb rmlA.
pET16b-Mtb rmlA plasmid was transformed to E. coli BL21(DE3) competent cells. For RmlA protein expression, E. coli BL21(DE3) carrying pET16b-Mtb rmlA was grown in 200 mL of LB broth containing Amp at 37 °C for 3 h followed by induction with 0.25 mM IPTG at 30 °C for another 3 h. The induced cells were harvested by centrifugation, resuspended in 4 mL lysis buffer (20 mM Tris-HCl, pH 8.0, 500 mM NaCl, 20% glycerol, and 1 mM phenylmethyl sulfonylfluoride) and sonicated. After centrifugation at 16,000 × g for 30 min, the supernatant was applied to a 1.0-mL column volume of Ni-NTA Superflow (Qiagen, Venlo, the Netherlands). The column was washed with 20 mL washing buffer (lysis buffer with 40 mM imidazole), and the RmlA protein was eluted by 10 mL elution buffer (lysis buffer with 200 mM imidazole).
The purified RmlA protein was detected by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and identified by Western blot. The gel of SDS-PAGE was stained with Coomassie blue R-250. For Western blot analysis, the proteins on the gel were transferred to a nitrocellulose membrane and incubated with (anti)-polyhistidine clone HIS-1 antibody (Sigma, St. Louis, MO) followed by antimouse-IgG-AP conjugate (Sigma). The protein bands on the membrane were finally visualized in BCIP/NBT solution.
HPLC analysis
The D-glucose-1-phosphate thymidylyltransferase activity of the purified RmlA was detected through HPLC analysis. The enzyme reaction was performed in 50 µL of 50 mM Tris-HCl (pH 7.5) containing 10 mM D-Glc-1-P, 4 mM dTTP, 2 mM MgCl2, and 0.1 µg purified RmlA at 37 °C for 30 min, and 20 µL of the reaction was injected to Nova-Pak C18 column (3.9 × 150 mm; Waters, Manchester, UK). Mobile phase was 500 mM KH2PO4 with a flow rate of 0.8 mL/min at a column temperature of 25 °C. The substrate dTTP and the product dTDP-D-Glc were monitored at 254 nm.
Colorimetric assay
A pyrophosphatase-coupled colorimetric assay for RmlA enzyme activity was developed with the purified RmlA protein. The enzyme reaction was carried out in a 96-well microtiter plate containing 50 mM Tris (pH 7.5), 1 mM dithiothreitol, 10% glycerol, 5 mM MgCl2, 0.2 mM dTTP, 1mM D-Glc-1-P, 45 ng purified RmlA protein, and 0.04 unit of Saccharomyces cerevisiae pyrophosphatase (Sigma; CAS # 9024-82-2) up to a final volume of 50 µL. One unit of pyrophosphatase was defined as the amount of pyrophosphatase that caused the appearance of 1.0 µmol of Pi per minute at pH 7.2 at 25 °C. The enzyme reaction was initiated by adding the purified RmlA protein. After incubation at 37 °C for 3 min, the enzyme reaction was terminated by adding 50 µL malachite green reagent (0.03% (w/v) malachite green, 0.2% (w/v) ammonium molybdate and 0.05% (v/v) Triton X-100 in 0.7 N HCl) at 37 °C for 5 min. The absorbance at 630 nm (OD630 nm) was monitored by a microplate reader (Thermo Scientific Multiskan Ascent). The reaction containing all reactants except RmlA was carried out as a negative control to correct the interference by substrates and buffer components. A standard curve of PPi concentration, which was obtained by performing the reaction containing only 0.01 to 0.08 mM PPi and 0.04 unit pyrophosphatase, was used for calculating the amount of PPi produced in RmlA reaction. In addition, a titration test with different amounts of pyrophosphatase was performed to determine if pyrophosphatase was used in excess in the assay.
To develop and optimize the colorimetric assay, the Z′ factor, a value to estimate the quality of the assay itself, was calculated with the equation defined by Zhang et al. 13 The “positive controls” were reactions with RmlA enzyme, and the “negative controls” were described above.
Kinetic analysis
Using the colorimetric assay, the kinetic properties of M. tuberculosis RmlA were determined in this study. The initial velocity was determined by performing the reactions at a different concentration of RmlA and at a different incubation time.
In the range of initial velocity, optimal pH, optimal temperature, the effect of Mg2+, and Km and Vmax values of M. tuberculosis RmlA were determined. Each parameter was measured in triplicate, and the data were analyzed using Microsoft Excel. The optimal pH was determined by performing the enzyme reaction in different buffers at the final concentration of 50 mM: NaH2PO4-NaOH (pH 5–6), Tris-HCl (pH 7–9), and NH4Cl-NH3·H2O (pH 10). The optimal temperature was measured by incubating the reaction mixture at 16 °C, 30 °C, 37 °C, 40 °C, 50 °C, and 60 °C, respectively. The optimal concentration of Mg2+ for the enzyme activity was determined by performing the enzyme reaction containing 0, 2.5, 5, 10, 20, and 40 mM Mg2+, respectively.
The Km and Vmax values of dTTP were determined by performing the enzyme reaction with various concentrations of dTTP (0.01–0.08 mM) with D-Glc-1-P at a saturating concentration (1 mM), and the Km and Vmax values of D-Glc-1-P were determined with various concentrations of D-Glc-1-P (0.05–0.40 mM) with dTTP at a saturating concentration (0.20 mM).
Results and Discussion
RmlA protein expression and purification
The Mycobacterium tuberculosis rmlA gene was cloned into the NdeI and XhoI sites of the pET16b vector generating a pET16b-Mtb rmlA expression plasmid. RmlA protein with a fused His-tag at the N-terminus was expressed in E. coli BL21(DE3) cells and purified by Ni2+ affinity chromatography. SDS-PAGE and Western blot analysis ( Fig. 1A ) confirmed the purified RmlA with an expected molecular weight of 33.67 kD and a perfect homogeneity.
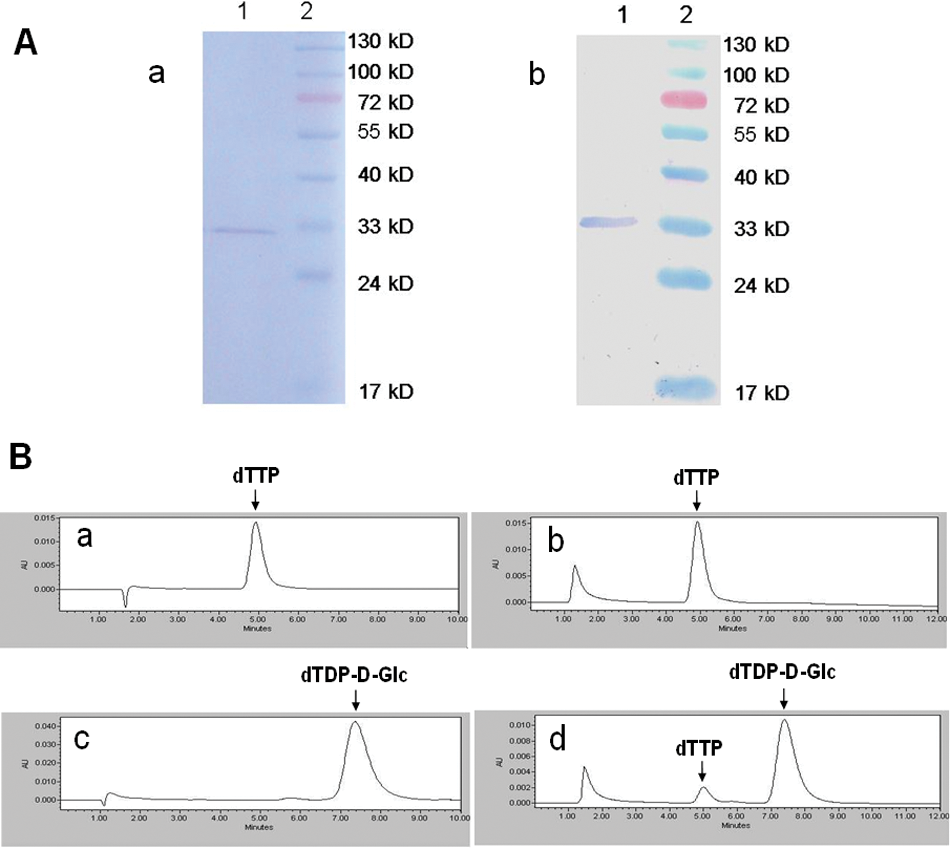
Purification and activity confirmation of M. tuberculosis RmlA. (
HPLC analysis
The enzyme activity of the purified RmlA was detected by HPLC analysis. After incubated at 37 °C for 30 min, the enzyme reaction mixture was injected into the column. A formation of dTDP-D-Glc and a decrease of dTTP were found in the enzyme reaction incubated at 37 °C for 30 min ( Fig. 1B ). This result confirmed that the purified RmlA had the D-glucose-1-phosphate thymidylyltransferase activity and could be used in the assay development and kinetic analysis below.
Colorimetric assay
Inspired by the enzyme assays for ADP-glucose pyrophosphorylase 14 and UDP-GlcNAc pyrophosphorylase, 15 here we developed a colorimetric assay for RmlA activity. In this assay, as shown in Figure 2A , RmlA catalyzed the formation of dTDP-D-Glc and PPi from dTTP and D-Glc-1-P, and pyrophosphatase hydrolyzed PPi into Pi. The Pi molecules were finally detected by the malachite green reagent. As a coupled enzyme, pyrophosphatase needed to be in excess. The titration test showed that the product Pi stopped increasing at 0.01 unit of pyrophosphatase, indicating that 0.01 unit was the minimum amount required for a complete reaction. To ensure this enzyme was in excess, fourfold of 0.01 unit of pyrophosphatase was finally used in the assay. In addition, as a key reagent, malachite green should be prepared freshly.
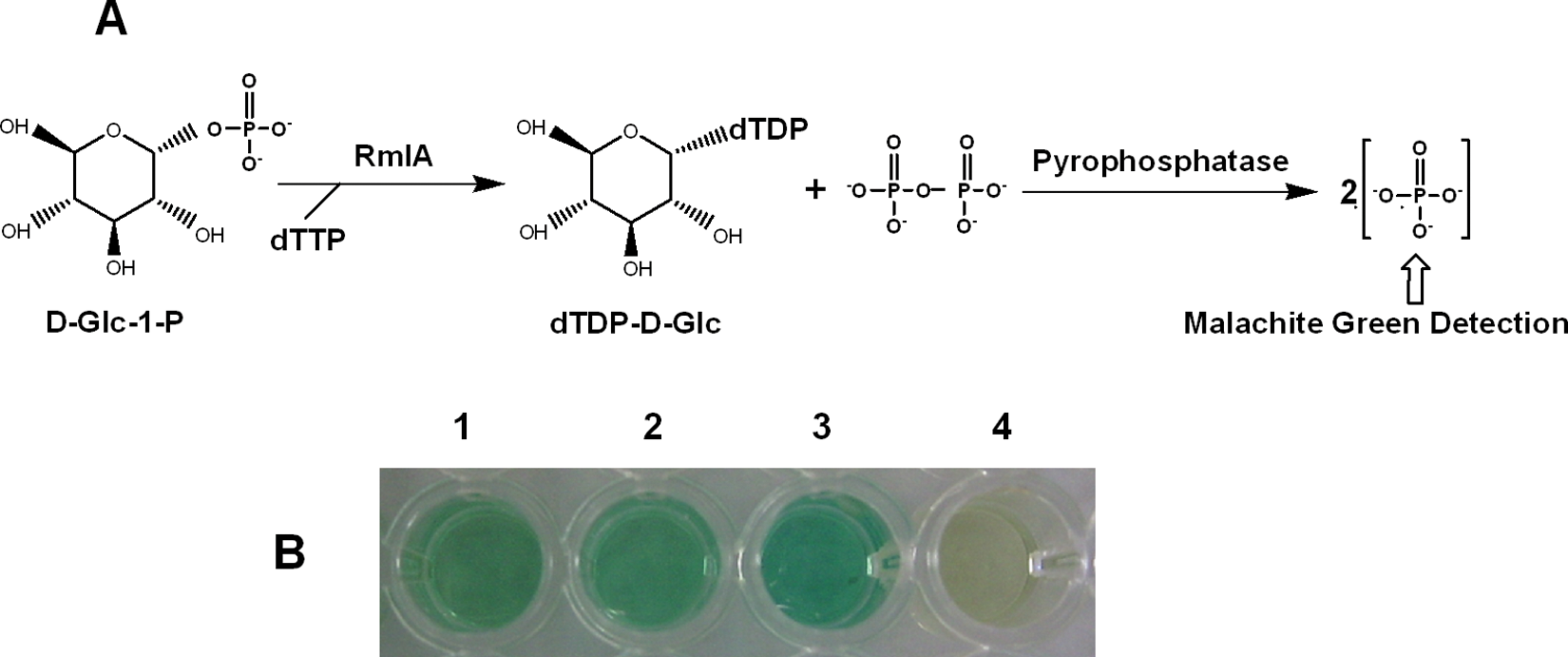
Colorimetric assay of RmlA. (
The results showed the RmlA reaction had an apparent color change from yellow-green to blue-green after malachite green reagent was added, which could be easily read at 630 nm ( Fig. 2B ). The ratio of signal to background was above 2.0 statistically. A Z′ value of 0.70 for this assay was obtained from 50 positive and 50 negative controls, which demonstrated the adequate sensitivity and reproducibility of the assay. Furthermore, the assay required only a small amount of substrates (0.05 µmol D-Glc-1-P and 0.01 µmol dTTP) and enzymes (45 ng purified RmlA protein and 0.04 unit pyrophosphatase) and could be finished in 96-well format at 37 °C within 10 min. Such a simple, rapid, and economical colorimetric assay is obviously more suitable for high-throughput screening of RmlA inhibitors than existing HPLC, CE, pHBH, or NADH oxidation-coupled methods.
Kinetic analysis
The kinetic properties of M. tuberculosis RmlA were measured using the developed colorimetric assay. The amount of PPi produced in the RmlA reaction was calculated from the standard curve of PPi concentration shown in the supplemental data. It was concluded from the time course and enzyme concentration curves that the initial velocity of RmlA reaction was limited within 3 min and 45 ng RmlA protein ( Fig. 3 ). In this range, the kinetic properties of M. tuberculosis RmlA were determined. The results showed M. tuberculosis RmlA had the relative maximal activity at pH 7.5 ( Fig. 4A ) and at the temperature of 37 °C ( Fig. 4B ). Mg2+ was necessary for RmlA activity, with an optimal concentration of 5 mM ( Fig. 4C ). This result of Mg2+ was consistent with the crystal studies on RmlA from E. coli 16 and Salmonella enterica LT2, 17 which indicated dTTP was the first substrate binding to the enzyme and Mg2+ was strictly required for lowering the activation energy and the binding of the second substrate D-Glc-1-P. The Km and Vmax values of M. tuberculosis RmlA against two substrates were calculated from the double-reciprocal plots shown in the supplemental data. The Km and Vmax against dTTP were 0.020 ± 0.004 mM and 0.003 ± 0.0003 mM/min, respectively, and against D-Glc-1-P were 0.069 ± 0.0050 mM and 0.002 ± 0.0001 mM/min, respectively. Comparing these results with the reported Km values of RmlA from other bacteria, such as S. enterica 7 (0.02 mM for dTTP and 0.11 mM for D-Glc-1-P) and Aneurinibacillus thermoaerophilus 9 (0.03 mM for dTTP and 0.20 mM for D-Glc-1-P), it was found that the Km values of RmlA against dTTP were identical, but against D-Glc-1-P were different.
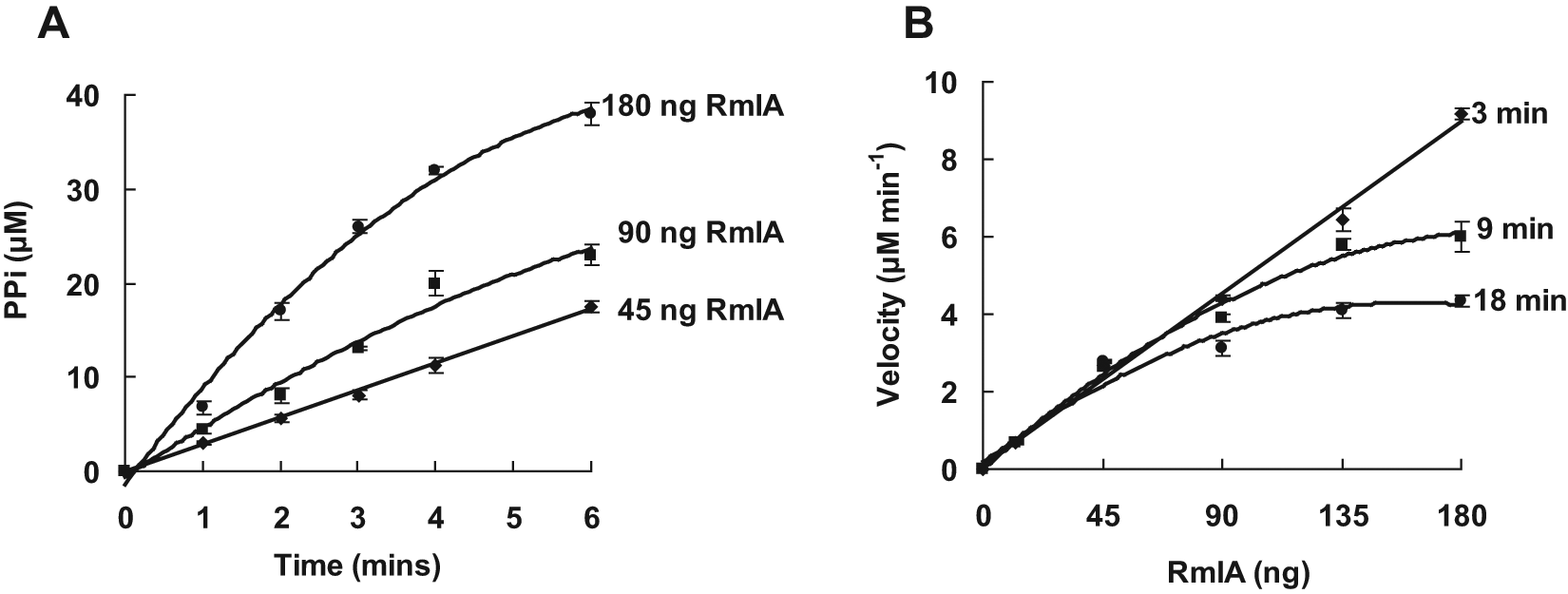
Time course (
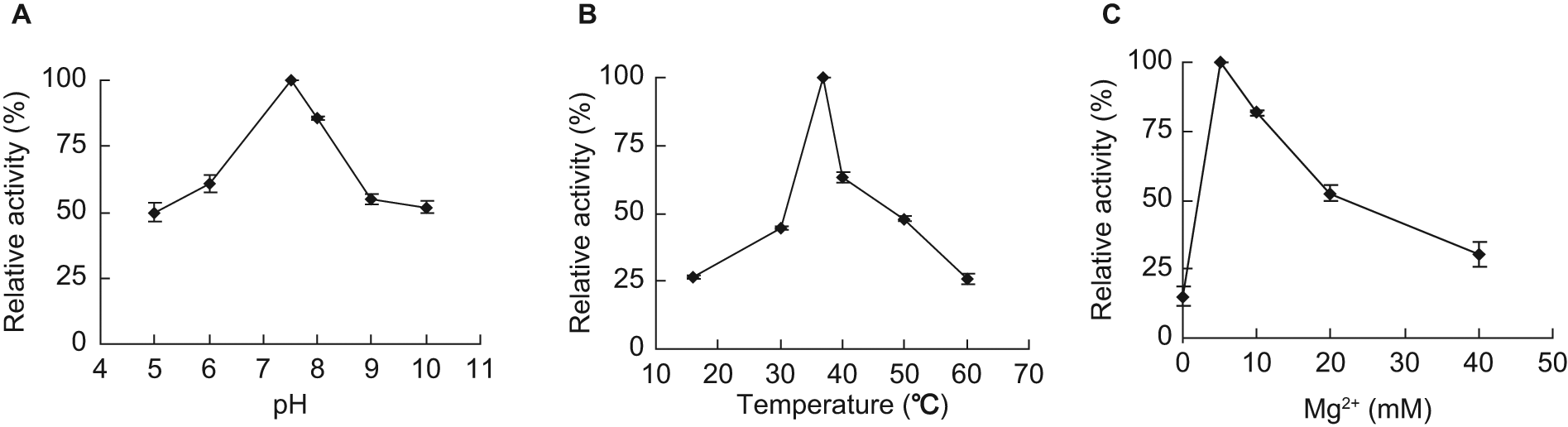
Determination of the optimal reaction conditions for M. tuberculosis RmlA.
In summary, we developed a colorimetric assay for RmlA in this study. Coupled with pyrophosphatase and detected by malachite green reagent, the assay allowed the fast and accurate determination of RmlA activity. With the assay, we measured the kinetic properties of M. tuberculosis RmlA including optimal pH, optimal temperature, the effect of Mg2+, and Km and Vmax values. These results will facilitate the high-throughput screening of RmlA inhibitors from large chemical libraries.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.