
Abstract
Misregulation of the Wnt pathway has been shown to be responsible for a variety of human diseases, most notably cancers. Screens for inhibitors of this pathway have been performed almost exclusively using cultured mammalian cells or with purified proteins. We have previously developed a biochemical assay using Xenopus egg extracts to recapitulate key cytoplasmic events in the Wnt pathway. Using this biochemical system, we show that a recombinant form of the Wnt coreceptor, LRP6, regulates the stability of two key components of the Wnt pathway (β-catenin and Axin) in opposing fashion. We have now fused β-catenin and Axin to firefly and Renilla luciferase, respectively, and demonstrate that the fusion proteins behave similarly as their wild-type counterparts. Using this dual luciferase readout, we adapted the Xenopus extracts system for high-throughput screening. Results from these screens demonstrate signal distribution curves that reflect the complexity of the library screened. Of several compounds identified as cytoplasmic modulators of the Wnt pathway, one was further validated as a bona fide inhibitor of the Wnt pathway in cultured mammalian cells and Xenopus embryos. We show that other embryonic pathways may be amendable to screening for inhibitors/modulators in Xenopus egg extracts.
Introduction
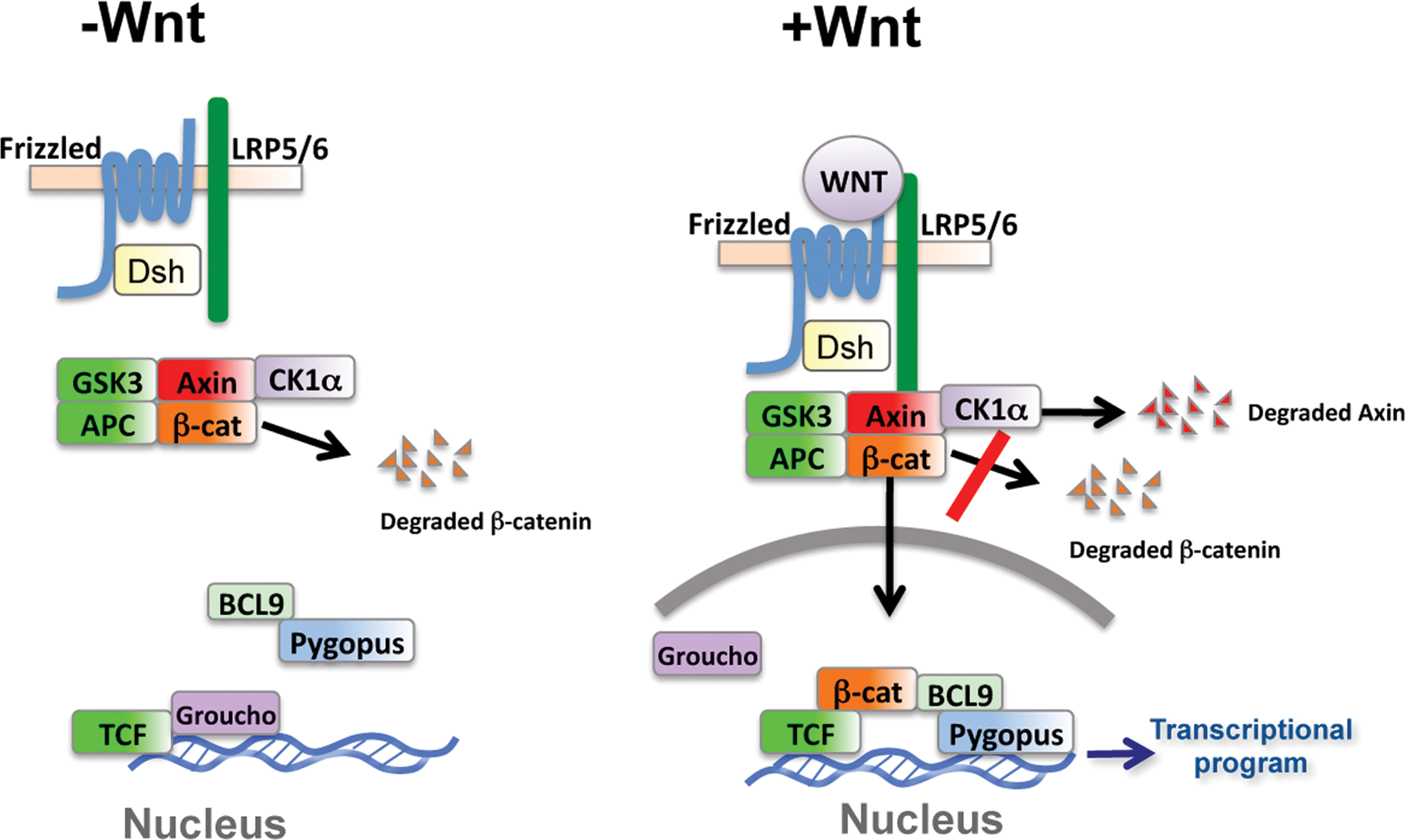
Model of the Wnt pathway. In the absence of Wnt ligand, β-catenin is degraded upon its recruitment to a complex containing the scaffold protein Axin. Within this complex, β-catenin is phosphorylated and targeted for degradation. In the presence of Wnt, degradation of β-catenin is inhibited and Axin degradation is stimulated. β-catenin accumulates, enters the nucleus, and initiates a Wnt pathway–specific transcriptional program. In many cases, activation of the Wnt pathway results in cellular growth and proliferation.
Degradation of β-catenin occurs in a complex primarily assembled by the scaffold protein Axin. In addition to β-catenin, Axin binds with high affinity the two kinases, glycogen synthase kinase-3 (GSK3) and casein kinase 1α (CK1α), and the tumor suppressor adenomatous polyposis coli (APC). 4 Within this complex, β-catenin is phosphorylated by CK1α at serine 45. This creates a priming site for subsequent and progressive phosphorylation by GSK3 at threonine 41, serine 37, and serine 33.5,6 Phosphorylated β-catenin is recognized by β-TRCP, a specificity subunit of the Skp1-Cullin-F-box (SCF) ubiquitin ligase, which catalyzes the polyubiquitination of β-catenin. Polyubiquitinated β-catenin is subsequently degraded by the proteosome. 7 The continual degradation of β-catenin ensures that cytosolic β-catenin levels are maintained below the threshold for Wnt target gene regulation. Axin has also been shown to be ubiquitinated and degraded via the proteosome.8–10 Because the concentration of Axin is a limiting factor in the establishment of the β-catenin degradation complex, alterations in Axin levels are likely to have a dramatic effect on Wnt signaling. 3 Axin stability has been shown to be regulated by phosphorylation and poly(ADP) ribosylation via the actions of GSK3 and Tankyrases, respectively.3,8–10 In the case of the latter, small-molecule inhibitors of Tankyrases have been shown to be potent inhibitors of the Wnt pathway. 10
Xenopus laevis has played a prominent role in our understanding of early vertebrate development. In the case of Wnt signaling, the activation of this pathway on the future dorsal side of the early Xenopus embryo is a critical event in the formation of the Spemann organizer, a tissue-organizing center found in vertebrates. 11 Ectopic activation of the Wnt signaling in the future ventral side of the embryo induces a second organizer that can coordinate the formation of a complete secondary body axis.12,13 Conversely, early inhibition of Wnt signaling results in animals that have reduced dorsal-anterior structures (ventralized).14,15 To date, almost all major members of the Wnt pathway have been validated using the Xenopus axis specification assay.
The misregulation of Wnt signaling plays a critical role in many human diseases. 1 Because of this pathway’s role in human development, its early misregulation leads to developmental defects. A variety of developmental genetic defects are known to occur as a result of misregulation of Wnt signaling, including defects in limb formation (tetra-amelia), bone density, eye vascularization, and tooth formation.16–21 In the adult, Wnt signaling plays an important role in stem cell renewal and tissue homeostastis. 22 Thus, perturbation of the Wnt pathway is associated with human degenerative diseases such as heart disease, schizophrenia, Alzheimer’s disease, and diabetes.23–25 Misregulation of the Wnt pathway is strongly associated with major human cancers. Loss of APC occurs in the majority (>80%) of sporadic colorectal cancer. 26 Oncogenic misregulation in Wnt signaling has been found in many other cancers including liver cancer, skin cancer, lung cancer, Wilms’ tumor, prostate cancer, and breast cancer (reviewed by Klaus and Birchmeier 27 ). Given the role of activating and loss-of-function mutations in the Wnt pathway in the genesis of a number of human diseases, there is great interest in identifying both positive and negative modulators of Wnt signaling. To date, there are currently no drugs in late clinical trials or in clinical use that specifically target the Wnt pathway.
Xenopus egg extracts contain cytoplasmic proteins, plasma membrane, organelles, amino acids, and nucleotides at or near physiological levels. 28 The Xenopus egg extract system represents a biochemical system that has been used to successfully recapitulate many complex biological reactions including microtubule and actin dynamics, DNA replication, nuclear envelope reformation, mitotic spindle assembly, DNA damage and repair, apoptosis, and oxygen homeostasis.28–36 Taking advantage of the ability of Xenopus extracts to reconstitute a wide variety of complex biology reactions, severals screens using Xenopus extracts have been previously performed to isolate modulators within various cellular compartments. These include screens for inhibitors of actin assembly, spindle assembly, and proteasome-mediated degradation.37–39 To our knowledge, this system has not been applied for high-throughput screening (HTS) of small molecules that target signal transduction pathways.
Xenopus extracts currently represent the only a simple biochemical system in which the dynamics of β-catenin and Axin degradation can be simultaneously monitored. We have previously shown that Xenopus egg extracts can recapitulate regulated degradation of β-catenin and Axin using a recombinant protein that encodes the intracellular domain of the Wnt coreceptor, LRP6.8,18,40 An assay for the identification of regulators of β-catenin turnover in Xenopus extracts has been previously described, although to our knowledge, no specific compound has been identified and/or characterized using this assay. 41 In the current report, we describe in detail major modifications of this system that take advantage of LRP6-regulated β-catenin and Axin degradation in Xenopus egg extracts. In addition, we demonstrate that Xenopus extracts can be used to identify modulators of the Wnt pathway. Recently, we used this screen to identify the Food and Drug Administration–approved drug pyrvinium as a potent Wnt inhibitor, although a detailed description of this screen was not provided. 42 Finally, we demonstrate for the first time that Xenopus extracts can potentially be used to identify small-molecule modulators of other embryonic pathways such as the Hedgehog and the Notch pathways, for which there has been intense interest in developing inhibitors as potential therapeutics. To our knowledge, a reconstituted biochemical system for the identification of small-molecule inhibitors of the Hedgehog and Notch pathways has not been previously described.
Materials and Methods
Chemical compounds and recombinant proteins
Reagents 3,6-dihydroxyflavone, 5,7-dihydroxyflavone, catechin hydrate, and cycloheximide were all purchased from Sigma-Aldrich (St. Louis, MO). LRP6ICD was prepared as previously described with modifications.8 Bacterial pellets were denatured with 6 M guanidine HCl, and histidine-tagged LRP6ICD was bound to nickel resin and eluted with imidazole. Eluate was dialyzed to remove guanidine HCl. All chemical structures were drawn using CS ChemDraw Ultra. DMSO was used as vehicle for all compounds in all experiments unless otherwise stated.
In vitro–translated proteins
Radiolabeled β-catenin and Axin were generated in rabbit reticulocyte lysates (Promega, Madison, WI) according to the manufacturer’s instructions. β-catenin-FLuc and Axin-RLuc for HTS were generated in TNT SP6 High-Yield Protein Expression System (Promega) according to the manufacturer’s instructions.
Preparation of Xenopus egg extracts
Xenopus egg extracts preparation and degradation assays were performed as previously described with modifications. 43 Frogs are primed with 100 U of pregnant mare serum gonadotropin (PMSG, Sigma) 3 to 7 days before induction of ovulation. Ovulation is induced by administration of 500 U human chorionic gonadotropin (HCG), frogs are then placed in large bins contain approximately 2 L of 1x Marc’s Modified Ringer (100 mM NaCl, 2 mM KCl, 1 mM MgCl2, 2 mM CaCl2, 0.1 mM EDTA, 5 mM HEPES [pH 7.8]) per frog. Frogs are allowed to lay eggs for 12 to 18 hr at 16 °C and then transferred to separate containers. Eggs are combined in a single beaker and washed three times in chilled (16 °C) 1x MMR to remove debris. Eggs are allowed to settle, and excess buffer is removed. Eggs are resuspended by gentle swirling in an ample volume of 2% cysteine in MMR. The jelly coat of the eggs should dissolve in approximately 5 min and is completed when settled eggs pack closely. Eggs are washed with a large volume of 1x MMR until the buffer is no longer cloudy. Eggs are washed 2 more times in Xenopus buffer (100 mM KCl, 0.1 mM CaCl2, 1 mM MgCl2, 10 mM potassium HEPES [pH 7.7], 50 mM sucrose). With a wide-mouth plastic Pasteur pipette, any eggs that appear white are removed, as these are apoptotic and could taint the extracts.
Eggs are transferred to chilled 30 mL centrifuge tubes. Once eggs have settled, excess buffer is removed. The eggs are first spun at 200 rpm for 2 min in a Sorval RC-6 at 4 °C. This packs the eggs so additional buffer can be removed. A crushing spin is performed for 5 min at 21 000g at 4 °C to separate the extracts into three major layers. The middle layer contains cytosol and is extracted by piercing through the top lipid layer with a p1000 tip and transferred to a fresh, chilled 30 mL tube. Cytochalasin B (10 µg/mL) is added to prevent actin polymerization. Extracts are spun at 21 000g, and cytosol is removed two more times. Final extracts should be a straw yellow color. Protease inhibitor (1000× stock; leupeptin, chymostatin, and pepstatin, all at 10 mg/mL in DMSO) and energy mix (150 mM creatine phosphate, 20 mM ATP, 2 mM EGTA, and 20 mM MgCl2 in water, pH 7.7) are added and mixed. Extracts are aliquoted, snap frozen, and stored in liquid nitrogen. Extracts can maintain β-catenin and Axin degradation activity for more than 1 year.
384-well assay conditions
Previously prepared extracts were thawed rapidly and placed on ice. In vitro–translated (IVT) β-catenin-FLuc (1 mL), Axin-RLuc (1.5 mL), and LRP6ICD (400 nM) were added to 100 mL of egg extracts and rotated end over end at 4 °C for 10 min. Samples were dispensed into chilled 384-well plates (5 µL/well) manually using a 24-channel repeat pipetter and kept in a 4 °C cold room just prior to addition of compounds. Library compounds were transferred by pin tool (custom apparatus, Harvard Medical School) or by Echo550 acoustic plate reformatter (Labcyte, Vanderbilt Institute of Chemical Biology) to plates at 25 °C to a final concentration ranging from ˜40 to 200 µm in 2% DMSO. Plates were sealed, vortexed gently, and incubated for 4 hr at 25 °C. After incubation, firefly and Renilla luciferase activities were measured using the Dual-Glo Luciferase Assay (Promega) per the manufacturer’s suggested protocol. Luminescent signal was acquired on an EnVision plate reader (PerkinElmer, Waltham, MA) or FDSS system (Hamamatsu, Bridgewater, NJ). For testing dilutions, egg extracts were diluted in Xenopus buffer.
Screen statistics
We normalized each plate using the method outlined in Malo et al. 44 The normalization procedure uses Tukey’s median polish to remove systematic row and column biases (e.g., edge effects). 45 The differences of the normalized log intensities between the Axin and β-catenin wells for each plate were obtained (equivalent to the ratio on the nonlog scale). Compounds were identified as hits if the log differences were in the 5th and 95th percentiles of the empirical distribution in both replicates.
For Kruskal-Wallis analysis, data were transformed into z-scores (z = \frac{x-\mu}{\sigma}) and separated into 3 groups (activators, inhibitors, and inactive compounds) by thresholding the z-scores, where z < –3, z > 3, and –3 < z < 3, respectively. Application of the Kruskal-Wallis analysis indicated that the population means among these three groups were significantly different (P = 1.07e-19).
Reporter assays
For cell-based luciferase assays, HEK 293 STF cells were seeded into 96-well plates at subconfluent levels and luciferase activities measured by Steady-Glo Luciferase Assay (Promega). Luciferase activities were normalized to a viable cell number using the CellTiter-Glo Assay (Promega). All graphs were made in Prism 4 (GraphPad Software, San Diego, CA) with nonlinear regression fit to a sigmoidal dose-response curve (variable slope).
Cell lines
HEK 293 STF cells were a gift from Jeremy Nathans. HEK 293 and IEC-6 were purchased from ATCC. An HEK 293 cell line stably expressing firefly luciferase under the control of the CMV promoter (CMV-Luc) was generated using standard methods. Cell lines were maintained in DMEM, 10% fetal bovine serum, and antibiotics. Cells were treated with compounds and/or Wnt for 24 hr unless otherwise stated.
Antibodies and plasmids
Immunoblotting was performed using the α-β-catenin (BD Transduction Labs, Franklin Lakes, NJ) and α-tubulin (Santa Cruz Biotechnology, Santa Cruz, CA) antibodies. β-catenin-FLuc and Axin-RLuc fusions were generated using standard PCR-based cloning strategies and subcloned into the pCS2 vector. The bacterial expression construct of LRP6ICD has been described.8
Immunoblot and immunofluorescence
For immunoblot, cells were lysed in nondenaturing buffer (50 mM Tris-Cl [pH 7.4], 300 mM NaCl, 5 mM EDTA, 1% [w/v] Triton X-100) and the soluble fraction used for immunoblotting. For immunofluorescence, cells were seeded on fibronectin-coated coverslips, fixed in 3.7% formaldehyde, stained, and mounted in VectaShield with DAPI (Vector Laboratories). Images were acquired using a CoolSNAP ES camera mounted on a Nikon Eclipse 80i fluorescence microscope with 60× or 100× objectives.
Xenopus laevis studies
Xenopus embryos were in vitro fertilized, dejellied, maintained, and injected as previously described. 46 Compounds were dissolved in 100% DMSO. For test compound bathing experiments, embryos were cultured in MMR, to which compounds were added at the indicated concentrations. At stage 10, embryos were washed three times with a large volume of MMR on a rocking platform to remove residual test compound. Capped β-catenin FLuc and Axin RLuc mRNA was generated using Message Machine (Ambion, Austin, TX) according to the manufacturer’s instructions.
Results
Characterization of β-catenin-FLuc and Axin-RLuc
Consistent with previous studies, we show that degradation of β-catenin and Axin are regulated by the intracellular domain of the Wnt coreceptor (LRP6) in Xenopus extracts (LRP6ICD;
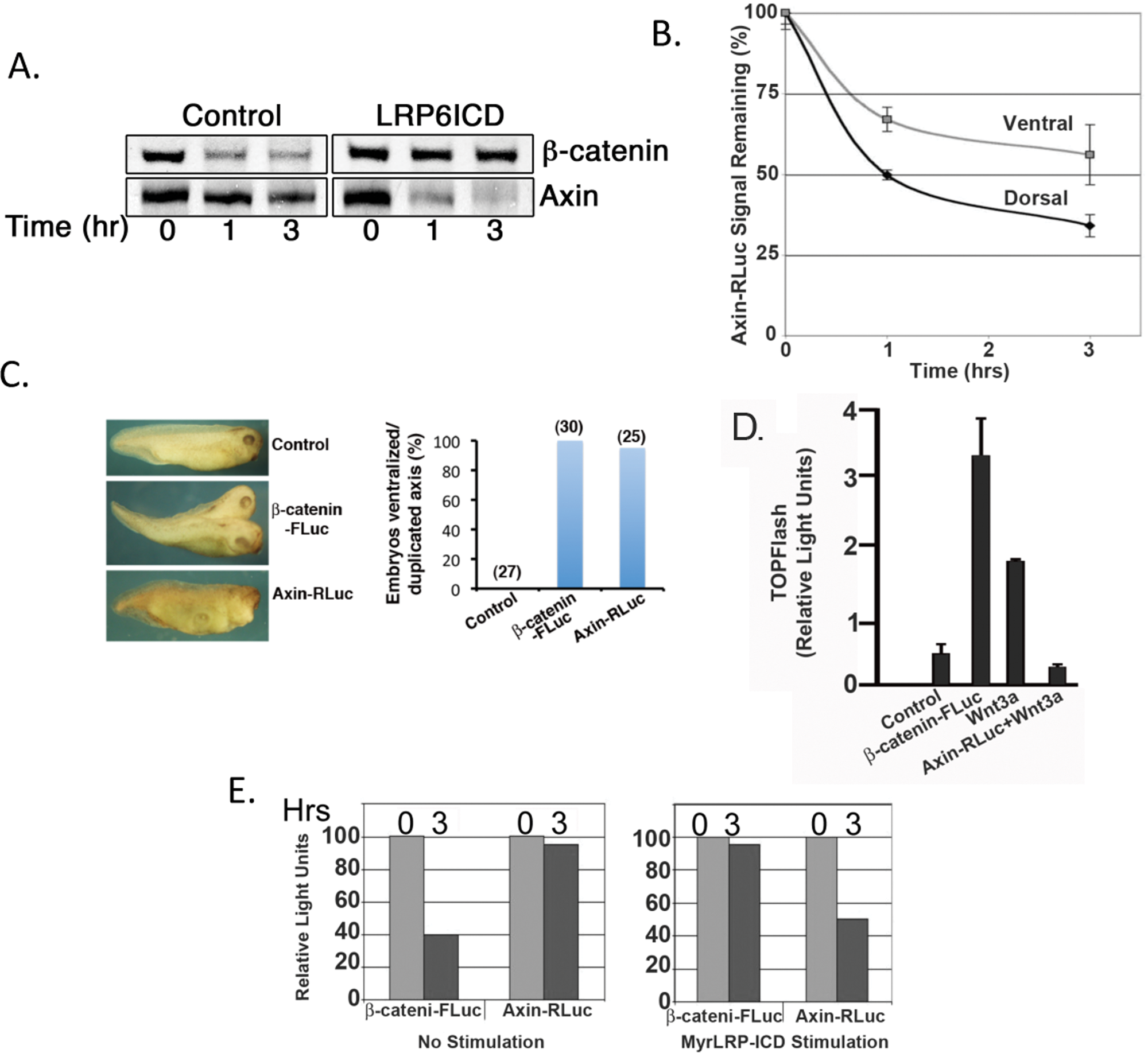
β-catenin-FLuc and Axin-RLuc behave similar to their wild-type proteins. (
Establishment of assay for β-catenin-FLuc– and Axin-RLuc–regulated stability by LRP6 and adaptation to a high-throughput format
An advantage of Xenopus egg extracts is that they can be readily scaled up for large volumes (liters). Such extracts were used as the basis for the purification of the anaphase promoting complex.
49
For our preliminary HTS, we were able to obtain sufficient amounts of Xenopus extracts (100 mL) from 100 female frogs injected with HCG 20 hr prior to egg collection (
For adapting to a high-throughput format for screening, we first demonstrate we can reliably and robustly detect changes in β-catenin-FLuc luciferase activity in response to a Wnt pathway activator (lithium) or inhibitor (Axin protein) in a 384-well plate format (
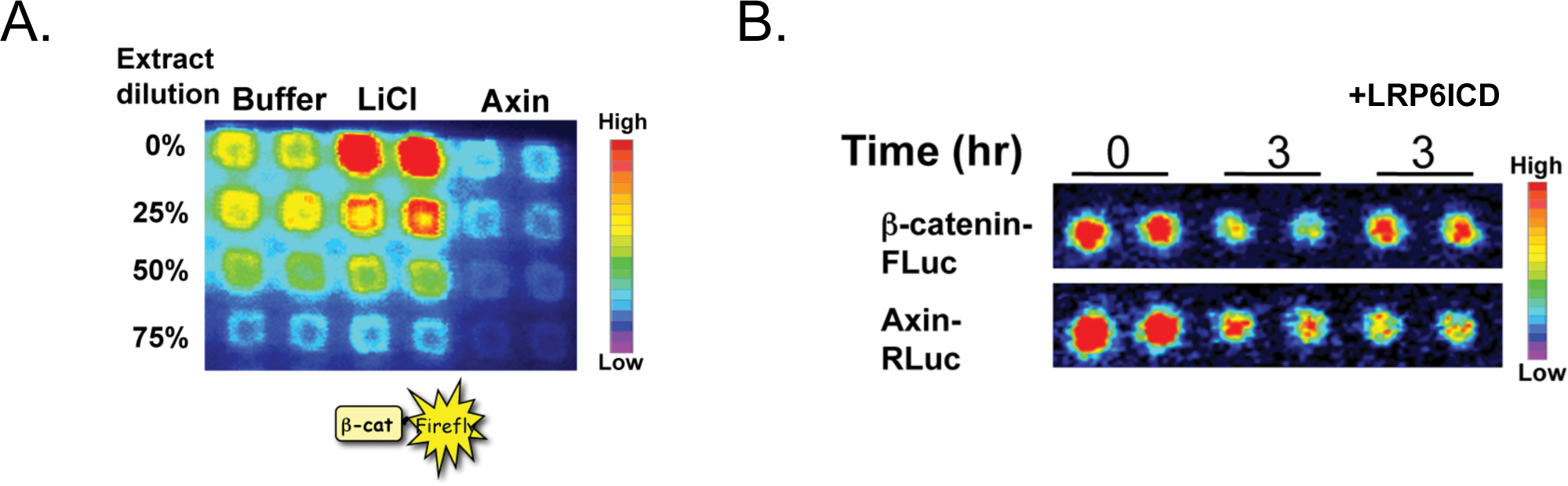
The regulated degradation of β-catenin-FLuc and Axin-RLuc in Xenopus extracts can be adapted for high-throughput screening. (
Results of biochemical screen for modulators of the Wnt pathway using diverse set of compound libraries
Preliminary screening and characterization of compounds was performed at the Vanderbilt Institute for Chemical Biology using commercially available ChemDiv and Chembridge chemical libraries. The distribution of signal strength from a representative plate after incubation with compounds is shown in
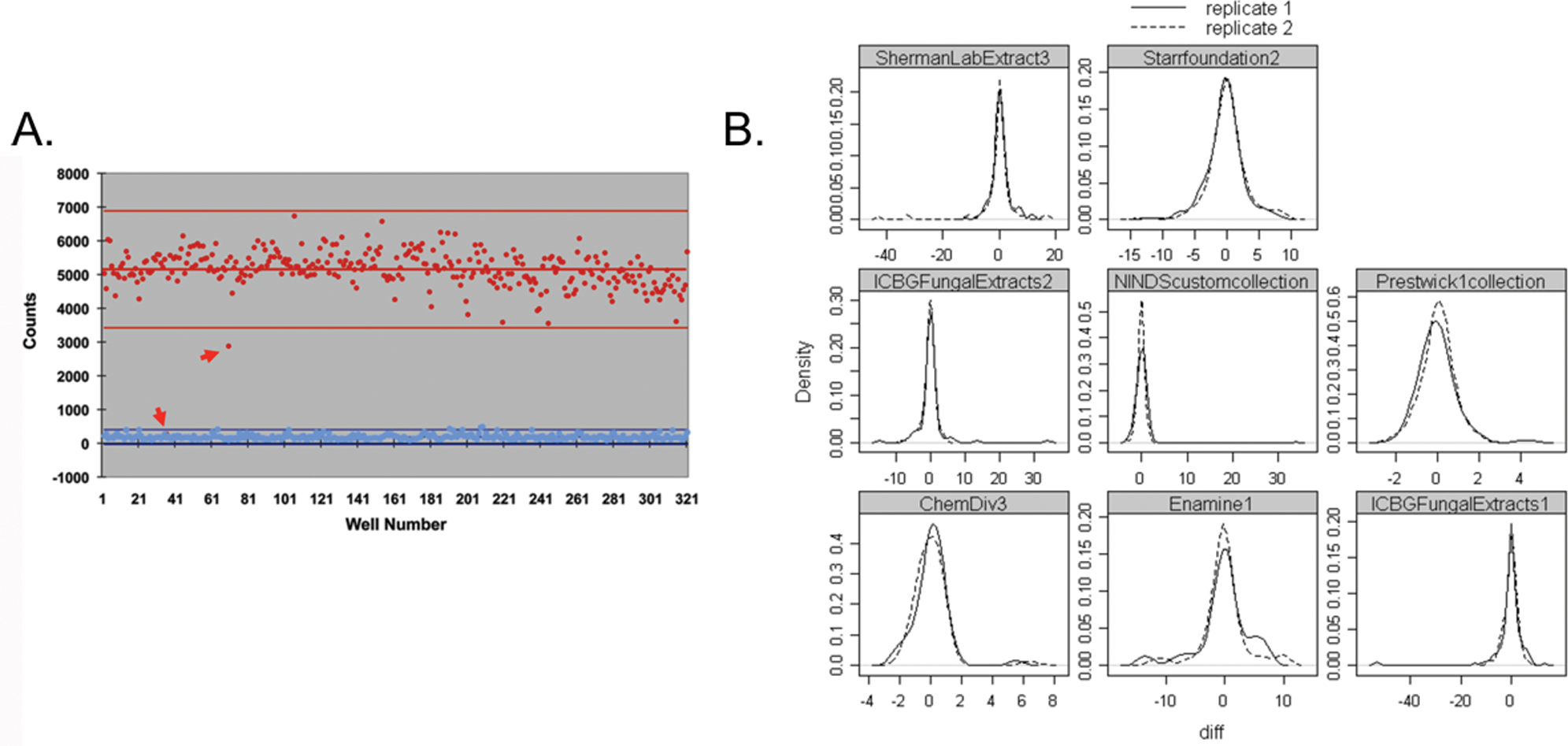
Behavior of screened compounds in Xenopus extracts that recapitulates Wnt signaling. (
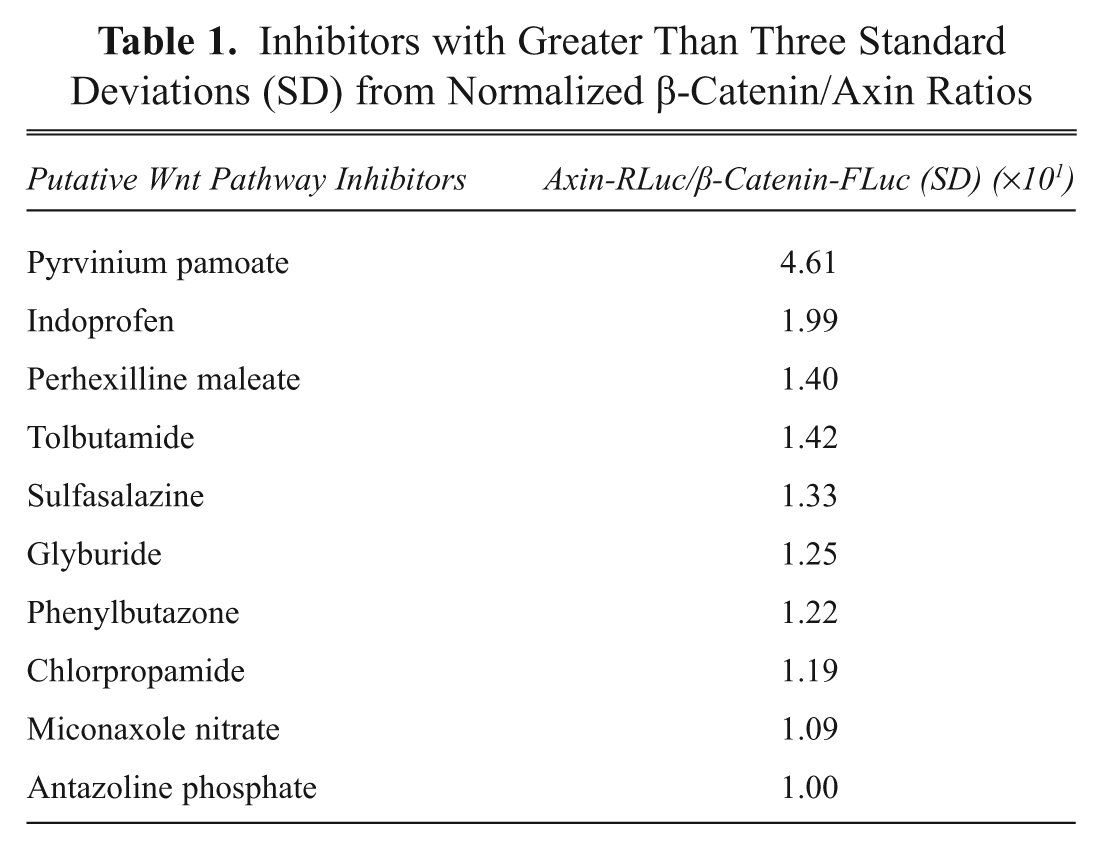
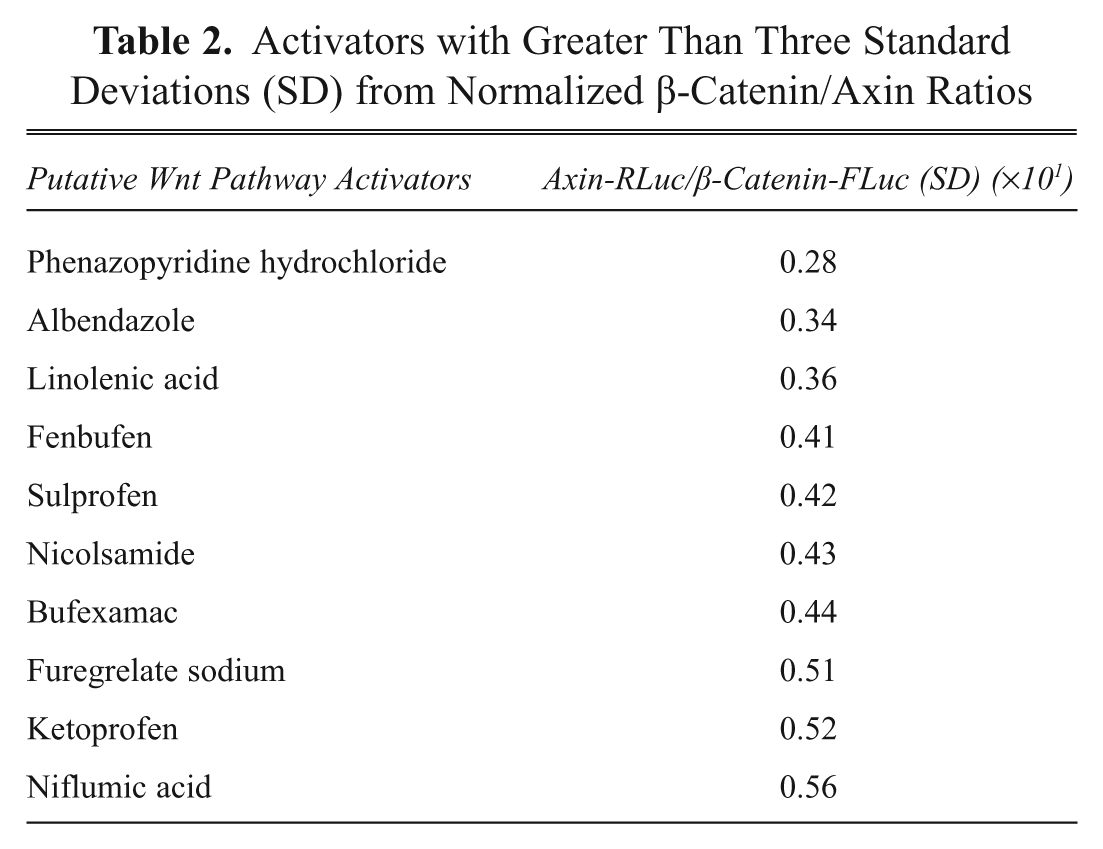
Approximately 6400 compounds were screened, and 125 hits (2% hit rate) for compounds that perturbed β-catenin-FLuc and/or Axin-RLuc were identified that gave signals three standard deviations above background. Further confidence in the assay resulted from the fact that we could identify structurally related compounds that altered β-catenin-FLuc signals and Axin-RLuc signals greater than three deviations from the mean (
Screening at Harvard was performed with three distinct libraries sets (total of approximately 8500 compounds): (1) Chembridge/ChemDiv, (2) NINDS bioactives, and (3) natural product (from Dr. Jon Clardy, Harvard Medical School). To optimize identification of hits with a high degree of confidence, we used an analysis method that is a modification of the method reported by Malo et al. and uses Tukey’s median polish to remove systematic row and column biases (e.g., edge effects;
If the compound was identified as a hit in both replicates, then it was added to the candidate list. Approximately 80 compounds were identified from screening the three classes of libraries with a high degree of confidence using our data analysis program. Of these, 60 compounds were from bioactive libraries (including Food and Drug Administration [FDA]–approved drugs), and 20 were hits from natural product extracts. Of the 60 compounds that were identified from the bioactive libraries, many are present in multiple libraries that we screened. Strikingly, we identified many such compounds in screens of different libraries, giving us another level of confidence that they are true hits. Priorities were assigned to compounds that stimulated Axin and inhibited β-catenin degradation. Compounds that solely stimulate or inhibit Axin turnover were prioritized because little is known as to the mechanism of Axin turnover, and these compounds would likely target proteins that are important for Axin degradation (e.g., tankyrase or E3 ligase for axin).11,51 Lower priority was assigned to compounds that inhibited both Axin and β-catenin degradation because they likely target common components of the degradation machinery (e.g., proteasome). Finally, compounds that inhibit both Axin and β-catenin-luciferase signals are likely (directly or indirectly) inhibitors of luciferase. A partial list of inhibitors and activators identified that are FDA-approved drugs is shown in
Inhibitors with Greater Than Three Standard Deviations (SD) from Normalized β-Catenin/Axin Ratios
Activators with Greater Than Three Standard Deviations (SD) from Normalized β-Catenin/Axin Ratios
Characterization of 3,6-dihydroxyflavone as an inhibitor of the Wnt pathway
To provide further evidence that identified compounds modulated Wnt signaling, we carried out extensive cell- and organism-based characterization of the activities of select compounds. A description of a particularly potent compound (pyrvinium) has already been described elsewhere.
41
In addition, we identified two compounds (3,6-dihydroxyflavone and 5,7-dihydroxyflavone) in our screen that were structurally related to flavonoids (
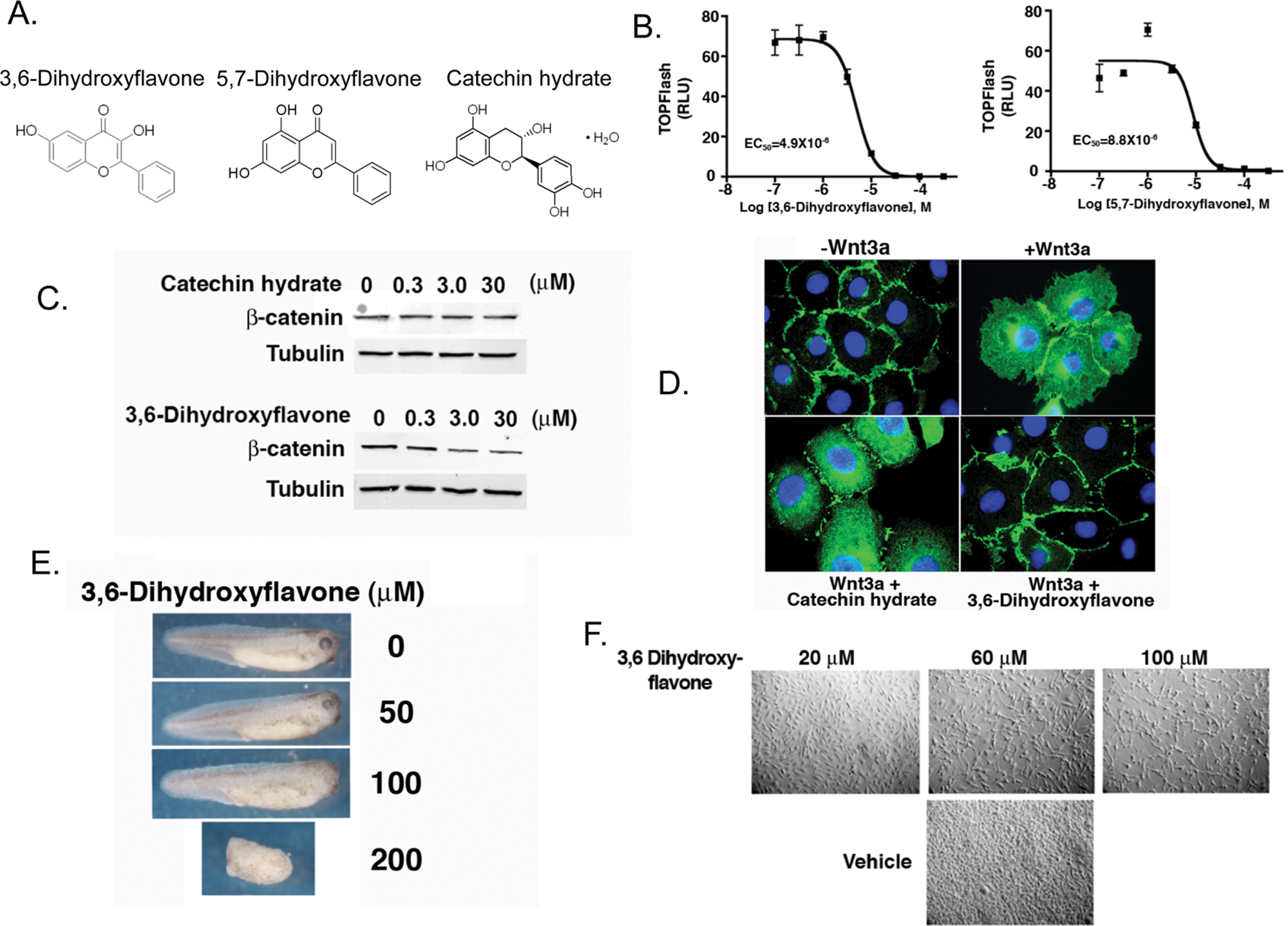
Validation of two hits identified from the Xenopus extracts screen as bona fide Wnt inhibitors. (
Discussion
We have developed a biochemical high-throughput assay using Xenopus egg extracts that recapitulates key biochemical steps in the Wnt pathway and screened a small set of chemically diverse compounds, which include FDA-approved compounds and natural product extracts. A major advance is that we are able to monitor two events of the Wnt pathway (β-catenin and Axin degradation) whose stabilities are inversely regulated in response to Wnt signaling. We have developed new data analysis software to identify potential hits with a high degree of confidence. For one of the hits (3,6-dihydroxyflavone), we validate that it inhibits both Wnt-mediated transcription and increase in β-catenin levels in cultured mammalian cells. We demonstrate that 3,6-dihydroxyflavone has in vivo activity consistent with inhibition of the Wnt pathway when incubated with Xenopus embryos. Finally, we show that 3,6-dihydroxyflavone inhibits the growth of a cancer line, MDA-MB-231, that has been shown to be dependent on Wnt signaling for growth and proliferation. Previous studies have demonstrated that certain flavonoids can inhibit Wnt signaling.53–55 For the most part, these studies indicate that flavonoids act in the nucleus, at the level of β-catenin and TCF/Lef1. It is likely that 3,6-dihydroxyflavone (and possibly 5,7-dihydroxyflavone) inhibit Wnt signaling via a distinct mechanism because our Xenopus extracts screen focused on compounds that alter the turnover of β-catenin and Axin. We observed decreases in steady-state cellular levels of β-catenin by immunoblotting and immunofluorescence upon treating cells with 3,6-dihydroxyflavone. Thus, in addition to identifying new flavonoids that inhibit Wnt signaling, we have uncovered a novel mechanism of action by which they act. Overall, these results provide strong evidence for the use of Xenopus egg extracts as a biochemical tool to identify small molecules that modulate Wnt signaling.
To date, there are no Wnt inhibitors in the clinic or in late clinical trials. Given the importance of the Wnt pathway in human development and disease, there has been intense interest in academic medical centers and in the pharmaceutical/ biotechnology industry to develop modulators of this pathway. Such compounds would have broad applications in the treatment of major human diseases such as schizophrenia, Alzheimer disease, heart disease, diabetes, and most major human cancers (colorectal, liver, and breast).
Our initial studies with NotchICD and Gli1 reported herein indicate that their turnover can be recapitulated in Xenopus egg extracts. As with the Wnt pathway, the Notch and Hedgehog pathways appear to lack obvious targets for drug development, and it has been particularly difficult to develop drugs against these pathways. 56 Our incomplete understanding of the circuitry of these pathways and how they are regulated is a major hindrance for drug development. Xenopus egg extracts have great potential for identifying compounds that inhibit/modulate Notch and Hedgehog pathways as well as other signaling pathways that are particularly difficult to target for drug development.
Footnotes
Acknowledgements
We thank members of the E. Lee and L. Lee lab for critical reading of the manuscript, Ashley Spann and Jessica Sweatt for assistance with tissue culture and experiments, and members of the Institute of Chemistry and Cell Biology–Longwood for assistance with screening. This work was supported by American Cancer Society Research Scholar grant RSG-05-126-01, National Cancer Institute grant GI SPORE P50 CA95103, National Institutes of Health grant 1 R01 GM081635-01 (E. Lee), American Heart Association Predoctoral Fellowship 0615279B, Molecular Endocrinology Training grant 5 T 32 DK007563 (C. A. Thorne), National Institute of General Medical Studies Medical-Scientist Training grant 5 T32 GM007347, Ruth L. Kirschstein National Research Service Award 4042800401 (A. J. Hanson), American Heart Association Predoctoral Fellowships 0615162B, and National Institutes of Health Cancer Biology Training grant T32 CA09592 (K. K. Jernigan). C. Wichiadit is supported in the Altschuler and Wu labs by Welch grant (I-1644) and Eureka grant (GM085442). E. Lee is a recipient of a Pew Scholarship in the Biomedical Sciences.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.