
Abstract
During the process of thrombopoiesis, invaginations of the plasma membrane occur in megakaryocytes. Since acetylsalicylic acid (aspirin), the most commonly used anti-inflammatory and antiplatelet drug, interacts with the lipid bilayers of the plasma membranes, this drug would affect the process of thrombopoiesis. In the present study, employing a standard patch-clamp whole-cell recording technique, we examined the effects of aspirin on delayed rectifier K+-channel (Kv1.3) currents and the membrane capacitance in megakaryocytes. Using confocal imaging of di-8-butyl-amino-naphthyl-ethylene-pyridinium-propyl-sulfonate (di-8-ANEPPS) staining, we also monitored the membrane invaginations in megakaryocytes. Aspirin suppressed both the peak and the pulse-end currents with a significant increase in the membrane capacitance. Massive di-8-ANEPPS staining after treatment with aspirin demonstrated the impaired membrane micro-architecture of megakaryocytes. This study demonstrated for the first time that aspirin induces microscopic surface changes in megakaryocytes. Such surface changes were thought to stimulate thrombopoiesis in megakaryocytes as detected by the increase in the membrane invaginations.
Keywords
Introduction
Acetylsalicylic acid (aspirin) is the most commonly used nonsteroidal anti-inflammatory drug (NSAID) that relieves fever, pain, and inflammatory symptoms. 1 Since it exerts antiplatelet effects, aspirin is also used to prevent cardiovascular diseases, such as myocardial infarction, ischemic stroke, and vascular thrombosis. 2 –4 As for the molecular basis, aspirin suppresses the production of thromboxane A2 and thus inhibits the aggregation of platelets. 5 In patients with platelet dysfunction, whether it is congenital or acquired, the numbers of circulating platelets are often kept normal or slightly elevated. 6,7 These findings indicate the presence of a mechanism that compensates for such impaired platelet function. Previously, Sullivan and McDonald demonstrated in experimental animals that aspirin-induced platelet dysfunction stimulated reactive thrombopoiesis. 8 In their study, due to the increase in the size of platelets and the amount of 35S incorporation into the platelets, the enhanced production of thrombopoietin was thought to be responsible for the mechanism. Recent advances in molecular biology have revealed that thrombopoiesis consists of 2 stages: initial invaginations of megakaryocyte plasma membranes (demarcation membrane system, DMS) and following the release of platelets from enlarged megakaryocytes. 9,10 Since NSAIDs are generally known to interact with the lipid bilayers of the plasma membranes, 11 and since they actually induced microscopic surface changes in thymocytes, 12 aspirin would directly disturb megakaryocyte plasma membranes and thus affect the process of thrombopoiesis. To examine this, we employed a standard patch-clamp whole-cell recording technique and confocal imaging of a membrane-impermeant fluorescent indicator 13 to determine the effects of aspirin on megakaryocyte membrane deformations. Here, we show for the first time that aspirin induces microscopic surface changes in megakaryocytes. We also show that such surface changes stimulate thrombopoiesis in megakaryocytes as detected by the increase in the membrane invaginations.
Materials and Methods
Cell Sources and Preparation
Male Wistar rats more than 25 weeks old, supplied by Japan SLC Inc (Shizuoka, Japan), were deeply anesthetized with isoflurane and then killed by cervical dislocation. The protocol for animal use was approved by the Animal Care and Use Committee of Tohoku University Graduate School of Medicine. Single megakaryocytes were isolated from rat bone marrow as described in our previous study. 14 Briefly, bone marrow obtained from femoral bones was flushed with standard external (bathing) solution containing (in mmol/L) NaCl, 145; KCl, 4.0; CaCl2, 1.0; MgCl2, 2.0; HEPES, 5.0; bovine serum albumin, 0.01% (pH 7.2 adjusted with NaOH) and dispersed by repetitive pipetting. After removing large pieces of tissue, cells were washed twice by gentle centrifugation for a minute and resuspended in the standard external solution. They were maintained at room temperature (22°C-24°C) for use within 8 hours.
Electrical Setup and Patch-Clamp Recordings
We conducted standard whole-cell patch-clamp recordings using an EPC-9 patch-clamp amplifier system (HEKA Electronics, Lambrecht, Germany) as described previously. 12,14,15 The patch pipette resistance was 1 to 3 MΩ when filled with internal (patch pipette) solution containing (in mmol/L) KCl, 145; MgCl2, 1.0; EGTA, 10; Hepes, 5.0 (pH 7.2 adjusted with KOH). After a giga-seal formation, we applied suction briefly to the pipette to rupture the patch membrane. The series resistance of the whole-cell recordings was maintained below 10 MΩ during the experiments. All experiments were carried out at room temperature.
Drug Delivery
Aspirin, purchased from Nacalai Tesque Inc (Kyoto, Japan), was dissolved in the external solution at a final concentration of 3 mmol/L. We delivered the reagent to the cells by a standing hydrostatic pressure of 3 cm H2O from a nearby pipette as described previously. 12,15 Then, whole-cell membrane currents were recorded before and after 30-second exposure to the reagents. To rule out the possibility that the observed effect just resulted from the procedure of the reagent application, we simply applied the external solution to the cells and confirmed the absence of any significant changes in the channel currents (Figure 1A).
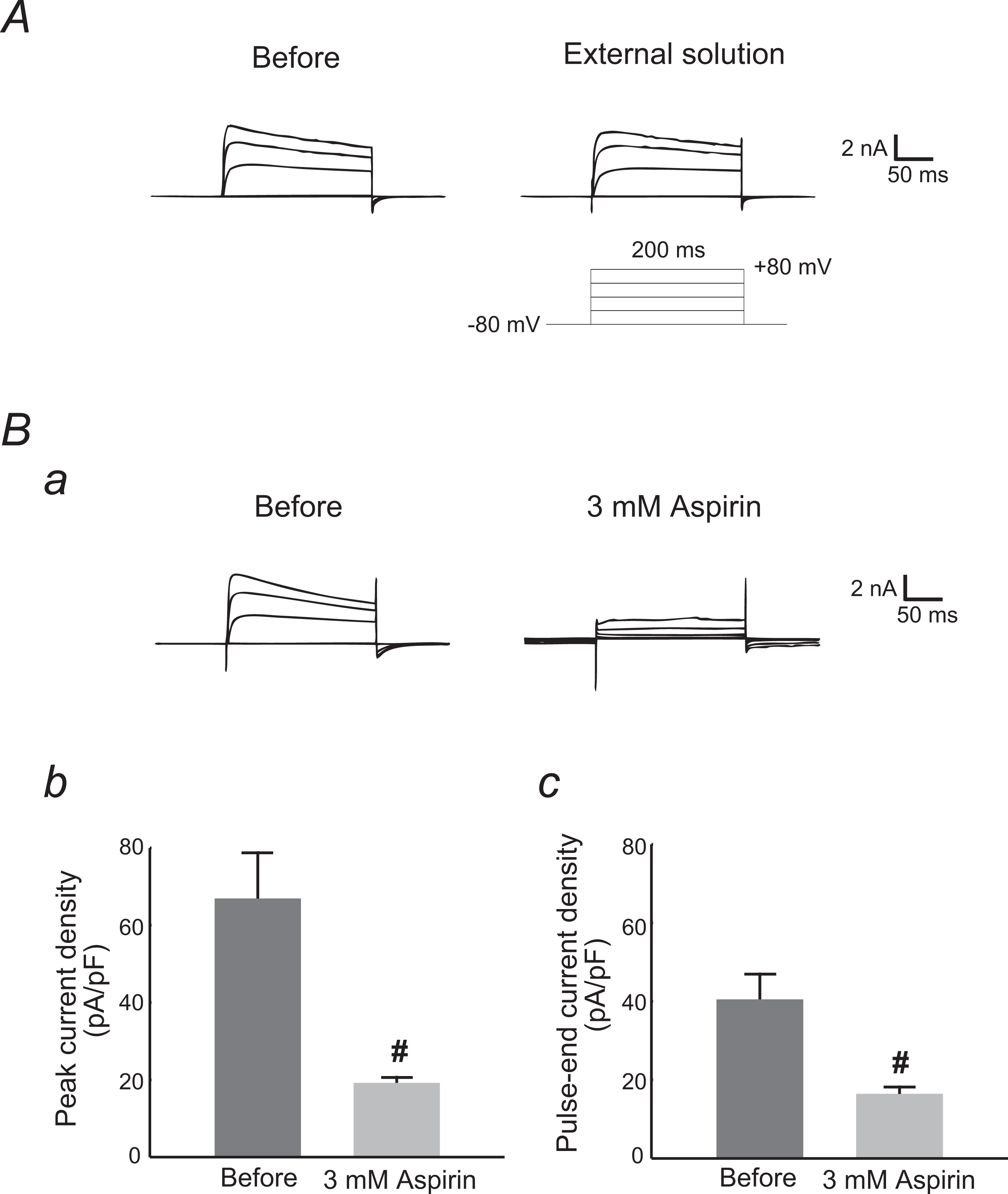
Effects of aspirin on Kv1.3 channel currents in rat megakaryocytes. Effects of the external solutions containing no drug (A) and 3 mmol/L aspirin (B). A, Ba: typical whole-cell current traces at different voltage steps recorded before and after the reagent application. The currents were elicited by voltage steps from the holding potential of −80 mV to −40, 0, 40, and 80 mV, as depicted in the voltage protocol in (A). Each pulse was applied for a 200-ms duration between 10-second intervals. Bb, peak current densities (peak currents normalized by the membrane capacitance) obtained from the records in a at the voltage step of 80 mV. Bc, pulse-end current densities (pulse-end currents normalized by the membrane capacitance) obtained from the records in a at the voltage step of 80 mV.
Membrane Capacitance Measurements
To measure the membrane capacitance of the megakaryocytes, we employed a sine plus DC protocol using the Lock-in amplifier of the EPC-9 Pulse program. An 800-Hz sinusoidal command voltage was superimposed on the holding potential of −80 mV. The membrane capacitance (Cm), as well as membrane conductance (Gm) and series conductance (Gs), was continuously recorded before and after 30 seconds of exposure to the reagents during the whole-cell recording configuration. Specific Cm was expressed per unit spherical cell surface area (μF/cm2).
Di-8-ANEPPS Staining
After adding aspirin at the final concentration of 3 mmol/L or after washing it out, single megakaryocytes were incubated for 30 minutes at room temperature in the external solution containing styryl membrane indicator, di-8-butyl-amino-naphthyl-ethylene-pyridinium-propyl-sulfonate (di-8-ANEPPS; Biotium, Hayward, California; final concentration 20 μmol/L). Fluorescent images were taken using a TE 2000-E Nikon Eclipse confocal microscope (Nikon, Tokyo, Japan).
Statistic Analyses
Data were analyzed using PulseFit software (HEKA Electronics, Lambrecht, Germany), IGOR Pro (WaveMetrics, Lake Oswego, Oregon), and Microsoft Excel (Microsoft Corporation, Redmond, Washington) and reported as means ± standard error of the mean (SEM). Statistical significance was assessed by 2-way analysis of variance followed by Dunnett’s or Student’s t test. A value of P < .05 was considered significant.
Results
Effects of Aspirin on Kv1.3-Channel Currents in Rat Megakaryocytes
Stepwise changes in the membrane potential, from the holding potential of −80 mV to the various depolarizing potential levels, evoked membrane currents in the megakaryocytes, showing a voltage-dependent activation and inactivation pattern characteristic of delayed rectifier K+-channels (Kv1.3; Figure 1A). 16 To investigate the effect of aspirin on the channel currents, we applied the external solution containing 3 mmol/L aspirin to the megakaryocytes and examined the changes in the whole-cell currents (Figure 1B). Since we delivered the drug by a puff application method, there was the possibility of partial and insufficient exposure of the drug to the cells. Therefore, we applied a higher concentration of aspirin than normally used for in vitro studies. 17,18 Similar to the effects of other NSAIDs on thymocytes, 12 aspirin suppressed the Kv1.3-channel currents in the megakaryocytes (Figure 1Ba). Since it significantly lowered the peak (from 66.9 ± 11.9 to 19.2 ± 1.46 pA/pF, n = 6, P < .05; Figure 2Bb) and the pulse-end currents (from 40.5 ± 6.47 to 16.4 ± 1.79 pA/pF, n = 6, P < .05; Figure 2Bc), the suppression pattern was similar to that of indomethacin, 12 showing a “C-type inactivation” pattern in the kinetic studies. 19 These results suggested that aspirin induced the conformational collapse of the selectivity filter (inactivation gate) within a pore-forming domain of the K+-channel.
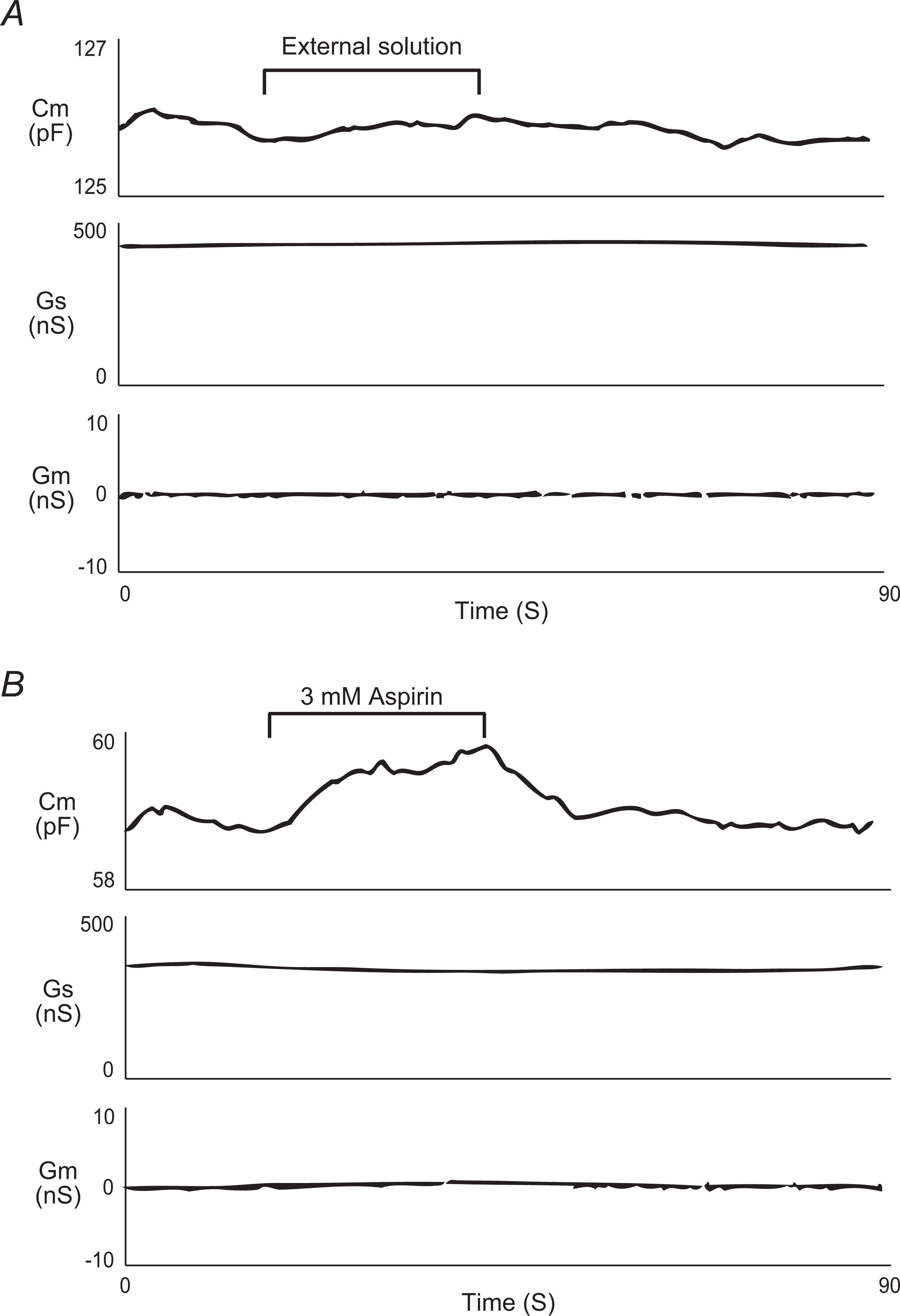
Aspirin-induced changes in meagakaryocyte membrane capacitance, series, and membrane conductance. After establishing the whole-cell configuration, external solutions containing no drug (A) and 3 mmol/L aspirin (B) were delivered for 30 seconds to single megakaryocytes. Membrane capacitance, series, and membrane conductance were monitored for at least 2 minutes. N = 5 for each trace. Cm indicates membrane capacitance; Gs, series conductance; Gm, membrane conductance.
Effect of Aspirin on Whole-Cell Membrane Capacitance in Rat Megakaryocytes
Since NSAIDs are known to interact with the lipid bilayers of plasma membranes, 11 and since aspirin is likely to induce the conformational collapse of the channels in megakaryocytes, it was conceivable that this drug would generate microscopic changes in the megakaryocyte membrane surface. In thymocytes, the microscopic changes in the cell surface area were best monitored by measurement of the whole-cell membrane capacitance. 12 In the present study, therefore, we employed this electrophysiological approach to detect the microscopic changes in the megakaryocyte membrane surface (Figure 2). Numerical changes in the specific Cm expressed per unit spherical cell surface area (μF/cm2) are summarized in Table 1. By simply applying the external solution alone to megakaryocytes (Figure 2A), we confirmed that our procedure of reagent application, a puff application by constant hydrostatic pressure with a nearby pipette, was without significant effects on the Cm and other parameters, such as Gm and Gs (Figure 2A, Table 1). However, the pipette inclusion of 3 mmol/L aspirin induced a significant increase in ▵Cm immediately after the application (Figure 2B, Table 1) with minimal changes in Gm and Gs. These results indicated that aspirin actually induced microscopic changes in the megakaryocyte membrane surface.
Summary of Changes in Membrane Capacitance After Application of Aspirin.a

Abbreviation: Cm, specific membrane capacitance.
a Values are means ± SEM.
b P < .05 vs ΔCm after application of the external solution.
Effects of Aspirin on Membrane Micro-Architecture in Rat Megakaryocytes
During the process of thrombopoiesis, invagination of the plasma membrane occurs in megakaryocytes providing the extracellular space required for the formation of proplatelets. 9,10,20 Since the value of specific Cm in megakaryocytes reflects the number of invaginated plasma membranes, 13,14 the increase in Cm would also predict the membrane invaginations induced by aspirin. In bright field images, megakaryocytes incubated in the aspirin-containing solution appeared to have more wrinkles on their cell surface, as demonstrated by the increase in black shadows most prominently in the center of the cells (Figure 3A, top right vs top left), which tended to decrease after washout of the drug (Figure 3A, bottom right). To examine whether the wrinkles represented the impaired membrane micro-architecture, we finally stained the megakaryocytes with di-8-ANEPPS, a membrane-impermeant fluorescent dye better retained in the outer leaflet of the plasma membrane. 13 In megakaryocytes that were not treated with aspirin, di-8-ANEPPS stained the peripheral plasma membranes of the cells (Figure 3B, top left). However, after incubating megakaryocytes in the aspirin-containing solution, the dye penetrated deep into the cells and stained throughout the extranuclear cellular volume (Figure 3B, top right). Then, after washout of the drug, the dye only stained the peripheral membranes again (Figure 3B, bottom right), indicating that the effect of aspirin was reversible. These results indicated that aspirin allowed the inflow of the dye into the folds of the membrane, suggesting that the membrane micro-architecture of megakaryocytes was impaired by aspirin.
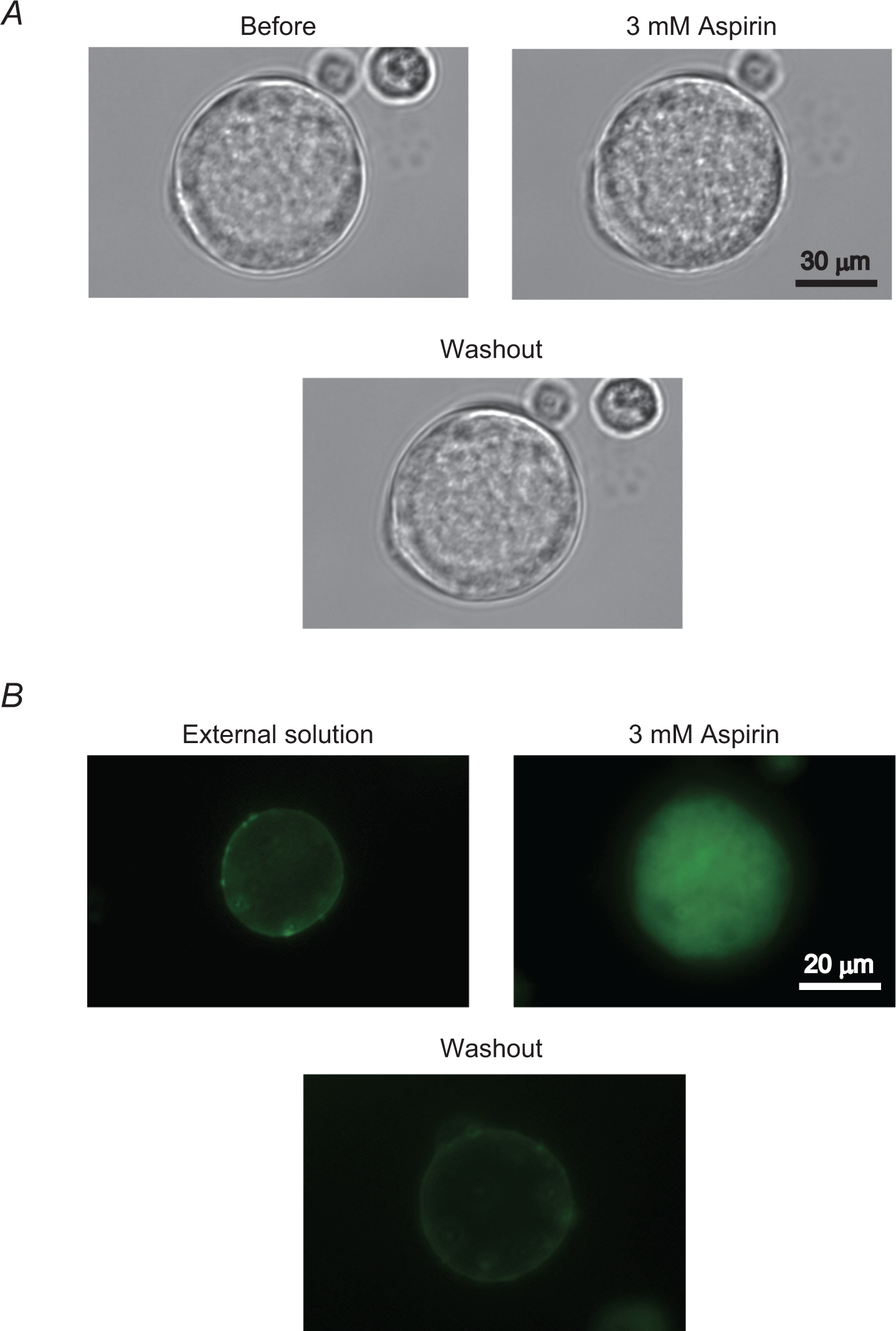
Images of aspirin-induced membrane surface deformations in megakaryocytes. A, Differential-interference contrast (DIC) microscopic images of megakaryocytes before (top left) and after incubation in the external solution containing 3 mmol/L aspirin (top right), and after washout of the drug (bottom right). B, Confocal fluorescence images of di-8-butyl-amino-naphthyl-ethylene-pyridinium-propyl-sulfonate (di-8-ANEPPS) labeled megakaryocytes after incubation in the external solutions containing no drug (top left) and 3 mmol/L aspirin (top right), and after washout of the drug (bottom right).
Discussion
In our previous studies, we examined the effects of salicylate, a deacetylated metabolite of aspirin, on Kv1.3-channel currents in both thymocytes and megakaryocytes. 12,21 In those cells, salicylate significantly suppressed the channel currents in a wide range of voltages. In the present study, similar to the effects of salicylate, aspirin markedly suppressed the channel currents in megakaryocytes. However, differing from the effect of salicylate, 12 aspirin significantly increased the membrane capacitance, inducing microscopic changes in the cellular surface. In addition, this drug actually impaired the membrane micro-architecture of megakaryocytes. Prior to the onset of proplatelet formation, cytoskeletal rearrangements occur in megakaryocytes, including the disassembly of centrosomes and the translocation of microtubules to the cell cortex. 10,22,23 Since the aspirin-induced changes in the membrane micro-architecture would include such cytoskeletal rearrangements, aspirin was then thought to trigger the plasma membrane invagination in megakaryocytes and thus stimulate thrombopoiesis.
Since salicylate is an amphiphilic and negatively charged compound, it partitions into the outer leaflet of the plasma membrane to generate the outward membrane bending. 24 Therefore, such induced changes in the membrane curvature were thought to be responsible for the current suppression by salicylate. 21 On the other hand, since aspirin is more lipophilic than salicylate, 25 it tends to accumulate more easily between the lipid bilayers of the plasma membranes after penetration. 26 Then, in addition to disturbing lipid–lipid interactions, the accumulated aspirin may directly perturb the composite domains of the channels from inside the membranes. This would include the constriction or the conformational collapse of the selectivity filters within the pore-forming domains. 19 Thus, aspirin was thought to induce the “C-type inactivation” pattern of the channel currents and to generate microscopic surface changes in the megakaryocyte plasma membranes.
Previously, we have actually measured the cytosolic Ca2+ concentration in mouse megakaryocytes and revealed the physiological mechanisms of its regulation. 27 As an extension of this study, McCloskey et al recently demonstrated in mouse megakaryocytes and platelets that Kv1.3 channels expressed in these cells regulate agonist-evoked Ca2+ increases and influence the circulating platelet counts. 16 In the present study, although we did not directly examine Ca2+ influx in response to K+ efflux, the previous findings strongly indicate that the cytosolic Ca2+ concentration in megakaryocytes is determined by the changes in the outward K+ currents. Since aspirin markedly suppressed the K+-channel currents in the present study, the cytosolic Ca2+ concentration in megakarycoytes was thought to be decreased. However, despite such absence of free Ca2+ in the cytoplasm, plasma membrane invaginations in the megakaryocytes were increased by aspirin. From these findings, aspirin-induced thrombopoiesis was not considered likely to be mediated through Ca2+ signaling. It was rather likely to be mediated through a direct effect of aspirin on the plasma membranes as demonstrated by the increase in the membrane capacitance. According to a recent report, the accumulation of phosphotidylinositol 4,5-bisphosphate (PI-4,5-P2) in megakaryocyte plasma membranes will facilitate the membrane invaginations and the following assembly of actin fibers required for thrombopoiesis. 20 Since aspirin inhibits the activity of phospholipase C that catalyzes PI-4,5-P2, 28 it would increase the accumulation of PI-4,5-P2 in the plasma membranes and thus may facilitate the process of thrombopoiesis.
Besides its anti-inflammatory effect, recent advances in pharmacological studies have revealed the therapeutic efficacies of aspirin for cancer progression, 29 atherosclerosis, 30 and diabetic nephropathy. 31 At low doses, aspirin is known to exert antiplatelet effect by irreversibly damaging the platelet function. 2 –5 However, at high doses, the effect is paradoxically attenuated by a phenomenon called “aspirin dilemma.” 32 Additionally, the present study demonstrated a novel aspect of aspirin as a direct stimulator of thrombopoiesis. In patients with platelet dysfunction 6,7 or drug-induced thrombocytopenia that is frequently encountered in the clinical settings, 33 the thrombopoietic activity of megakaryocytes is usually kept intact. In this regard, besides the use of thrombopoietin receptor agonist which is indicated for the treatment of refractory immune thrombocytopenia, 34 the use of high-dose aspirin may also be beneficial for the resolution of bleeding symptoms in those patients.
In summary, this study demonstrated for the first time that aspirin induces microscopic surface changes in megakaryocytes. Such surface changes were thought to stimulate thrombopoiesis in megakaryocytes as detected by the increase in the membrane invaginations.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.