
Abstract
Neural functioning and plasticity can be studied on different levels of organization and complexity ranging from the molecular and synaptic level to neural circuitry of whole brain networks. Across neuroscience different methods are being applied to better understand the role of various neurotransmitter systems in the evolution of perception and cognition. GABA is the main inhibitory neurotransmitter in the adult mammalian brain and, depending on the brain region, up to 25% of the total number of cortical neurons are GABAergic interneurons. At the one end of the spectrum, GABAergic neurons have been accurately described with regard to cell morphological, molecular, and electrophysiological properties; at the other end researchers try to link GABA concentrations in specific brain regions to human behavior using magnetic resonance spectroscopy. One of the main challenges of modern neuroscience currently is to integrate knowledge from highly specialized subfields at distinct biological scales into a coherent picture that bridges the gap between molecules and behavior. In the current review, recent findings from different fields of GABA research are summarized delineating a potential strategy to develop a more holistic picture of the function and role of GABA.
Introduction: Molecules, Networks, and Behavior
Neuroscience aims to elucidate the neural underpinnings of perception, cognition, and adaptive behavior. Neural functioning and plasticity can be studied on different levels of organization and complexity ranging from the molecular and synaptic level to neural circuitry of whole brain networks. Across neuroscience, different methods are being applied to better understand the role of different brain regions, networks, and neurotransmitter systems in the evolution of higher brain functions.
γ-Amino-butyric acid (GABA) is the main inhibitory neurotransmitter in the adult mammalian brain, formed by the decarboxylation of glutamate, an enzyme-driven process that primarily takes place in GABAergic interneurons. This synthesis is associated with two different isoforms of the glutamic acid decarboxylase (GAD), GAD65 und GAD67. Both isoforms can be found in most GABAergic neurons. Whereas GAD67 is distributed throughout the cell cytoplasm, GAD65 is highly concentrated in nerve endings, as such GAD65 is critically involved in GABA synthesis targeting vesicular release (Soghomonian and Martin 1998). GABA either binds to GABAA receptors, which are chloride permeable pentameric channels, eliciting a chloride influx and subsequent hyperpolarization in mature cells (in healthy condition) (Kaila and others 2014), or to GABAB receptors, which are G-protein coupled receptors interacting with potassium and calcium channels. This interaction either elicits a postsynaptic potassium efflux inducing a slow inhibitory potential or inhibits presynaptic calcium channels leading to a suppression of neurotransmitter release (Ulrich and Bettler 2007).
GABAergic interneurons are diverse. Present-day classification systems predominantly use morphological, electrophysiological, and neurochemical properties to determine specific subtypes. About 20 cortical and 20 hippocampal subtypes have been identified so far. Forty percent of cortical interneurons have fast-spiking (FS) electrophysiological properties; that is, basket and chandelier cells and express the cytoplasmic calcium-binding protein (CaBP) parvalbumin (PV); about 30% of the cortical interneurons express the neuropeptide somatostatin, for example, Martinotti cells, and about 30% express the serotonin receptor 3A, for example, vasointestinal peptide (VIP) expressing cells, calretinin (CR) expressing, fast adapting cells and reelin expressing, late-spiking cells. Calbindin (CB), another CaBP, is expressed in a variety of GABAergic cells, but is considered an inaccurate classification marker. Importantly, current classification systems are viewed as “work in progress” by most researches, and further modifications are warranted. Of note, the majority of studies on GABAergic cells exclusively focuses on interneurons; however, there are long-range GABAergic neurons that, for example, reciprocally connect the entorhinal cortex (EC) and the hippocampus (Caputi and others 2013).
During early development, GABAergic and glutamatergic progenitor cells follow different paths to reach their destination. GABAergic interneurons are produced in the ventral telencephalon and migrate tangentially into the neocortex, whereas glutamatergic excitatory neurons are generated in the ventricular zone of the dorsal telencephalon and migrate radially into the cortical plate (Sultan and others 2013). Depending on the brain region, GABAergic interneurons constitute 10% to 25% of the total number of cortical neurons. During maturation the expression and functioning of various receptors, for example, NMDA and AMPA receptors, as well as ion channels change several times.
As GABAergic transmission matures, interneurons start to act as circuit pacemakers. Especially FS interneurons seem to play a critical role in synchronizing network activity, enabling oscillations in different frequency bands, for example, the generation of gamma rhythms (Le Magueresse and Monyer 2013). These rhythms and transient changes thereof are of great interest to the understanding of various cognitive processes. As such sharp wave-associated ripple oscillations in the hippocampus are thought to emerge from pacing of pyramidal cells by PV+, FS basket cells; the development of theta activity, the major hippocampal rhythm during exploration, also seems to be strongly modulated by GABAergic interneurons. In humans, gamma activity as well as alpha activity have been linked to cortical information processing (Lozano-Soldevilla and others 2014); furthermore, dysfunctional gamma oscillations, for example, in the prefrontal cortex, seem to reflect working memory deficits, for instance, in patients with schizophrenia (Senkowski and Gallinat 2015). At the systems level, a number of electroencephalography (EEG) and magnetoencephalography (MEG) studies, as well as computational modeling, suggest that the GABAergic system plays a pivotal role in synchronizing larger neural cell populations. However, the relationship between oscillations and behavior remains to be fully elucidated.
Overall, there is a well-established and steadily increasing knowledge about the development, the onset of functioning, the subtypes, the electrophysiological properties and the target cells of GABAergic neurons (Le Magueresse and Monyer 2013). In the long run, however, findings from basic neuroscience need to be interpreted in the light of behavior (in health and disease). In the current review article, we summarize recent developments in GABA neuroscience, relating evidence from different levels of neural organization and complexity to behavior in both animals and humans.
Inhibition in the Hippocampal-Entorhinal Formation
The hippocampal-entorhinal formation is essential for the formation of spatial and episodic memories, assembling what happened, when and where (Knierim and others 2014). Specifically, the role of the hippocampus for spatial learning and memory is well established. Moreover, selective genetic manipulations in the forebrain or only in the hippocampus, revealed the significance of inhibitory GABAergic neurons, in particular PV+, FS interneurons, for oscillatory network activities, spatial coding, and hippocampus-dependent working memory (Caputi and others 2012; Fuchs and others 2007). The entorhinal cortex (EC) is a major input structure to the hippocampus. The superficial layer II and layer III of the EC are the origin of the perforant path terminating in the dentate gyrus and the temporo-ammonic pathway terminating in the CA1 region of the hippocampus. However, if and how the EC participates in information processing and episodic memory formation is still a matter of debate. In rodents, the EC can be divided into two subdivisions, the medial EC (MEC) and the lateral EC (LEC). Of note, the two subdivisions of the EC are postulated to be part of two parallel input streams: the spatial stream provides spatial or “where” information through the MEC, and the nonspatial stream provides feature or “what” information through the LEC, to the hippocampus (Knierim and others 2014) (Fig. 1A).
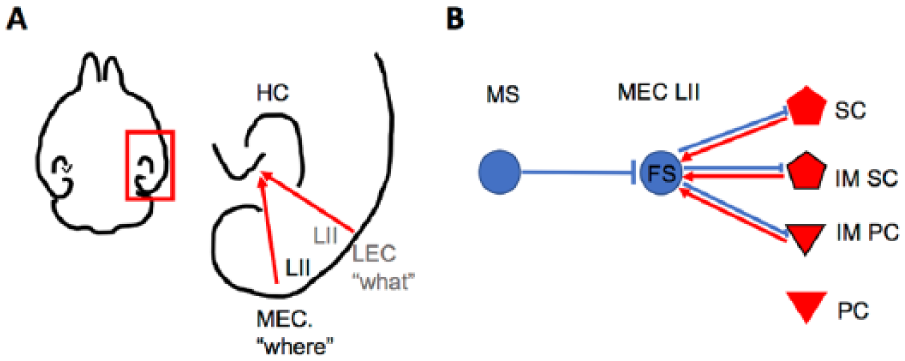
Local and long-range connectivity of inhibitory PV+, FS neurons in layer II of the MEC. (A) Scheme depicting the hippocampal-entorhinal formation in a rodent horizontal brain section (right). Boxed area shown at a higher magnification: input from LII of the MEC provides spatial “where” information, input from LII of the LEC provides feature or “what” information to the hippocampus (left). (B) Inhibitory (blue) FS interneuron in MEC LII receives inhibition via long-range GABAergic projection from the MS, and communicate locally with excitatory (red) SC, IMSC, and IMPC, but not PC in LII of the MEC. HC, hippocampus; MEC, medial entorhinal cortex; LEC, intermediate entorhinal cortex; L, layer; FS, fast-spiking neuron; SC, stellate cell; IMSC, intermediate stellate cell; PC, pyramidal cell; IMPC, intermediate pyramidal cell; MS, medial septum.
The wealth of information regarding inhibitory interneurons in the hippocampus is in stark contrast to the relatively scarce knowledge about interneurons and their function in the MEC, and even more so in the LEC. The MEC receives information from spatial input areas and during tetrode recordings in freely behaving rodents, neurons in the superficial layers of the MEC exhibit spatial-selective firing patterns. In vivo tetrode recordings in freely behaving animals are used to measure extracellularly field potentials from neuronal populations and spatially modulated firing of single neurons. The recorded action potentials from individual neurons, that is, spikes, differ in waveform and firing rate, which can be used to distinguish between spikes originating from FS inhibitory neurons and putative excitatory neurons (Buetfering and others 2014).
The best-studied examples are grid cells, which display multiple firing fields forming a hexagonal pattern in a two-dimensional environment (Hafting and others 2005). Recent experimental data highlight the role of inhibition for the formation of the particular firing pattern of grid cells. This notion is based on in vitro patch-clamp recordings of paired neurons demonstrating that putative grid cells, the so-called excitatory stellate cells in layer II of the MEC, do not communicate with each other, which is supported by in vivo recordings in freely behaving mice. However, these experiments showed that stellate cells are indirectly connected via FS interneurons (Couey and others 2013; Fuchs and others 2016). Notably, immunohistological experiments revealed that about half of all GABAergic neurons in layer II of the MEC are PV+, FS interneurons (Buetfering and others 2014). Interestingly, FS interneurons seem to be more broadly tuned as compared with spatial-selective grid cells (Buetfering and others 2014). Moreover, excitatory neurons in layer II of the MEC express either the CaBP CB or the extracellular matrix protein reelin. When taken electrophysiological and anatomical features into account, these two excitatory cell populations can be further subdivided into four distinct cell types that exhibit cell-type-specific local excitatory and inhibitory connectivity (Fuchs and others 2016) (Fig. 1B).
The LEC receives information from nonspatial input areas, that is, the perirhinal cortex and the olfactory bulb. Neurons in the LEC exhibit only sparse spatial selectivity although they respond to objects, the position of objects or odors (Knierim and others 2014; Leitner and others 2016), thereby encoding the “what” component of episodic memories. Little is known about the functional role or the connectivity of inhibitory neurons in the LEC. Of note, a recent study showed that also in the LEC GABAergic neurons are more broadly tuned to odors as compared with distinct populations of excitatory neurons in layer II of the LEC (Leitner and others 2016).
Finally, GABAergic neurons are critically involved in oscillatory network activities that may underlie numerous cognitive functions (Fuchs and others 2007). Supporting evidence for a crucial role of inhibitory neurons in shaping network activities are recent findings demonstrating that functionally related brain regions are often connected via reciprocal long-range inhibitory projections. Since the medial septum is considered the major pacemaker for rhythmic network activities in the hippocampal-entorhinal formation (Buzsáki 2002), the reciprocal inhibitory connectivity between the medial septum and the hippocampus (Freund and Antal 1988) and between the medial septum and the MEC (Fuchs and others 2016) are of special interest for hippocampal-entorhinal network activities. Moreover, long-range GABAergic neurons connect the MEC and the hippocampus (Melzer and others 2012), and the LEC and the hippocampus (Basu and others 2016). This long-range inhibitory connectivity between the medial septum, the hippocampus and the entorhinal subregions could orchestrate oscillatory activity and thereby support episodic memory formation.
Modulation of Cortical GABAergic Inhibition by Noninvasive Brain Stimulation
Functional assessment of GABAergic neurotransmission at high temporal and spatial resolution is required to learn more about the role of cortical inhibition in higher brain functions. In this context, noninvasive brain stimulation, specifically transcranial magnetic stimulation (TMS), has been successfully employed to assess the role of GABAergic inhibition in both animals and humans. Based on the physical principle of electromagnetic induction TMS allows for the activation of cortical neurons through the intact skin and skull. Repetitive TMS (rTMS), that is, the application of several hundred TMS pulses, is known to modulate cortical excitability even beyond the stimulation period (depending on the frequency and the pattern of stimulus trains). While several hundred pulses at 1 Hz reduce cortical excitability, frequencies of 5 Hz and above enhance it. A variety of protocols has been established in the field, including patterned stimulation protocols, for example, continuous and intermittent theta burst stimulation (cTBS and iTBS, respectively; Huang and others 2005).
However, the cellular and molecular mechanisms of rTMS-induced changes of cortical excitability, and consequently the neurobiology of rTMS-based therapies are not well understood (Cirillo and others 2017; Müller-Dahlhaus and Vlachos 2013; Pell and others 2011). The transient nature of rTMS-induced changes in motor evoked potentials (MEPs; lasting for several minutes to hours), and the fact that rTMS is neither context- nor input-specific makes it unlikely that associative plasticity of selective neural pathways and circuits, that is, long-term potentiation (LTP) and long-term depression (LTD) of excitatory postsynapses, underlies lasting TMS effects. Rather, changes in the cortical balance of excitation and inhibition may enable subsequent plastic changes of the network via “gating” and “metaplasticity” (Müller-Dahlhaus and Ziemann 2015; Ziemann and Siebner 2008).
Animal studies have shown that rTMS is able to affect distinct cortical inhibitory systems depending on the stimulation pattern employed (Fig. 2A), and that these effects interfere with sensory processing and learning (Mix and others 2010; Reis and others 2008; Thimm and Funke 2015). It turns out that iTBS most efficiently induces a strong reduction in the expression of PV+, FS interneurons, while cTBS and low-frequency (1 Hz) rTMS preferentially reduce the expression of CB, another CaBP found in non-FS interneurons (Markram and others 2004). CR+ interneurons which appear to preferentially target other interneuron populations (eg, the PV+ and CB+), were not affected by any stimulation protocol tested so far (Benali and others 2011) (Fig. 2B).
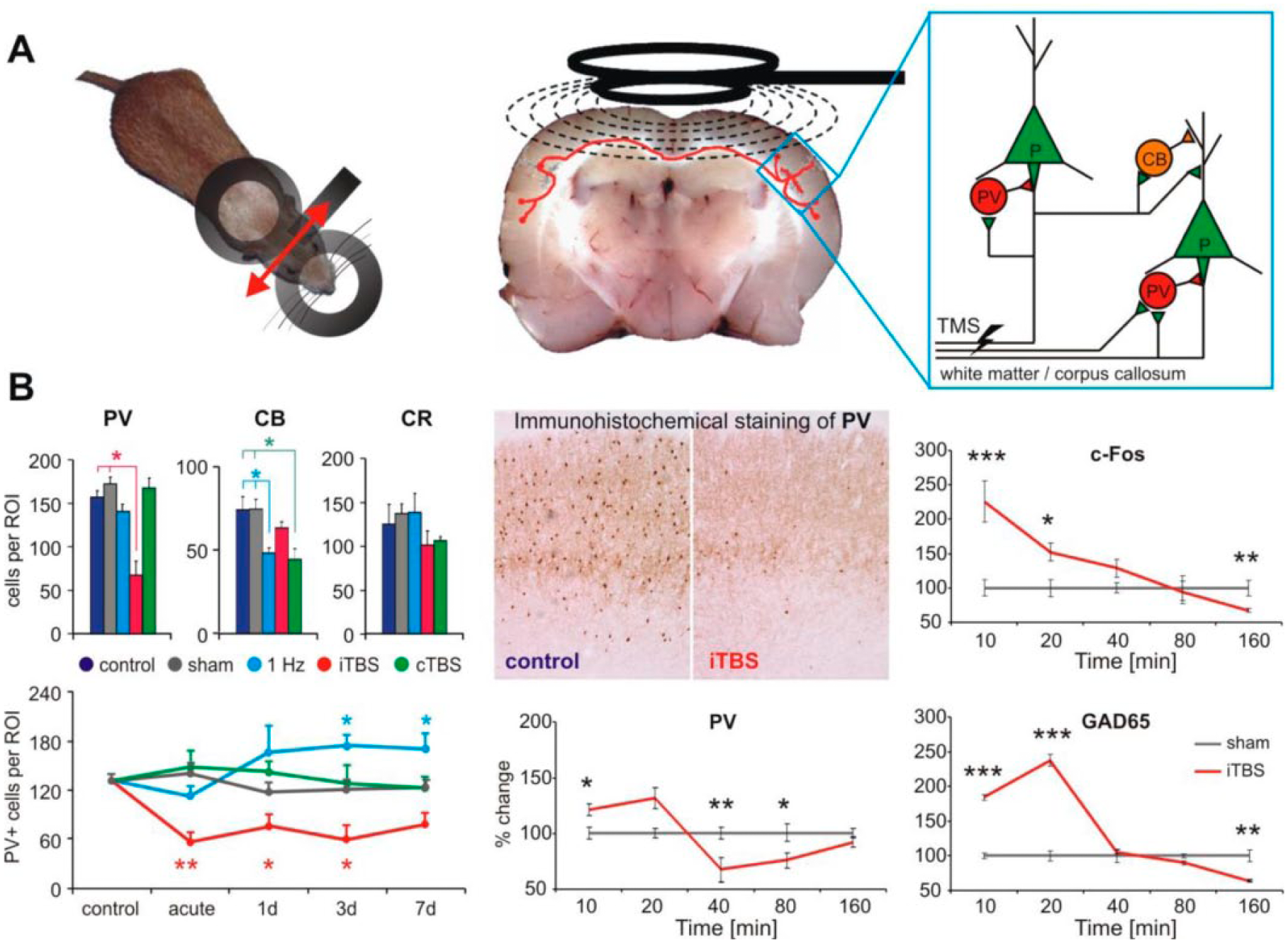
Modulation of the expression of activity markers of inhibitory cortical systems in a rat repetitive transcranial magnetic stimulation (rTMS) model. (A) rTMS was applied via a figure-8 shaped coil (2 × 70 mm) directed in a way to induce a mediolateral oriented electric field within the rat brain (left, red arrow) suitable to stimulate callosal axons (middle), thereby activating layer 2/3 pyramidal cells and interneurons (right). (B) Changes in the cortical (here motor cortex data shown) expression of the calcium-binding proteins parvalbumin (PV), calbindin (CB), and calretinin (CR) induced by either 1 Hz rTMS (1200 pulses in one train), intermittent theta burst stimulation (iTBS; bursts of 3 pulses at 50 Hz, repeated at 5 Hz with 2-second ON and 8-second OFF periodicity, 5 block of 600 pulses each) or continuous TBS (cTBS; single 40-second train of bursts in a block) compared with sham stimulation (left). iTBS strongly reduced PV expression, which could last several days without further interference (lower left). Expression of c-Fos and GAD65 were strongly increased immediately after one iTBS block while PV decreased after about 40 minutes (lower middle and right). *P < .05, **P < .01, ***P < .001. Part B of the figure has been redrawn using data of Benali and others (2011) and Hoppenrath and Funke (2013).
Rodent work has shown that only one block of iTBS (600 pulses within 192 seconds) induces a decline in PV, GAD67 and CB expression starting about 30 minutes after the stimulation and slowly recovering within 160 minutes (Hoppenrath and Funke 2013). However, a stronger reduction in PV expression obtained with five blocks of iTBS (at 15-minute intervals; Volz and others 2013) can also last for days while the decrease in CB caused by five cTBS blocks recovers more rapidly (Benali and others 2011). Early after iTBS (10-20 minutes) the immediate early gene product c-Fos and presynaptic GAD65 show enhanced expression, indicating that iTBS increases neuronal activity including that of GABAergic neurons (Fig. 3B, right panels). Apparently, distinct stimulation protocols can modulate the activity of different classes of GABAergic interneurons, with iTBS affecting the activity of FS interneurons that control pyramidal cell output activity via perisomatic inhibition, and 1 Hz and cTBS affecting the activity of non-FS (CB+) interneurons controlling the dendritic input to pyramidal neurons. Notably, these distinct interneuron populations and changes in the respective CaBPs (in part due to reduced interneuron cell number) are thought to be involved in the pathophysiology of psychiatric and neurological diseases such as schizophrenia and epilepsy (Lewis and others 2005).
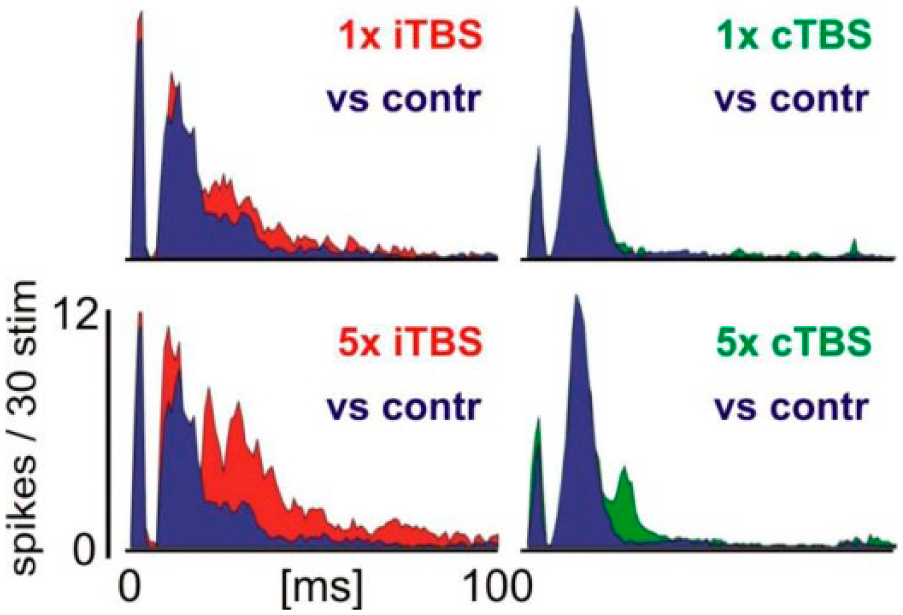
Increased cortical sensory responsiveness after intermittent theta burst stimulation (iTBS). With each of the 5 blocks applied (600p/block) iTBS increased sensory responses to whisker deflections (800 deg/s, 1.72 deg) in rat barrel cortex while continuous TBS (cTBS) had no significant effect. Especially late sensory response components suggested to be under control of recurrent inhibition are affected. Shown are peristimulus time histograms of single-/multi-unit recordings with responses obtained during sham-stimulation experiments in blue, verum iTBS in red, and verum cTBS in green. Responses to 30 identical stimulus repetitions were averaged. Modified from Thimm and Funke (2015).
The consequences of rTMS-induced modulation of GABAergic neurotransmission and its effect on perception and cognition remain to be fully elucidated. Recent basic science work employing optogenetic approaches has provided new insights into the functional role of inhibition for complex brain function (Letzkus and others 2011). These studies confirm previous work that discloses a relevance of disinhibition, that is, a transient reduction in cortical inhibition, in learning and memory formation. Hence, it is conceivable that rTMS modulates the ability of distinct cortical regions to express input- and context-specific plasticity by transiently changing local cortical GABAergic neurotransmission. Indeed, electrophysiological recordings of evoked sensory activity from the rat barrel cortex reveal a disinhibitory effect of iTBS, which was most prominent for the late intracortically mediated response components known to be relevant for the perception of stimuli (Thimm and Funke 2015) (Fig. 3).
Follow-up histology revealed a decrease in GAD67 and PV (but not CB) and increased c-Fos expression. No such prominent effect was found with cTBS, which decreased the CB and c-Fos expression but not that of PV. Another study revealed that iTBS, but not cTBS, improved the rat’s ability to learn a pure tactile orientation task (Mix and others 2010). Rats treated with iTBS shortly before each task block showed about a 50% increase in learning speed compared to sham treated animals (Fig. 4).
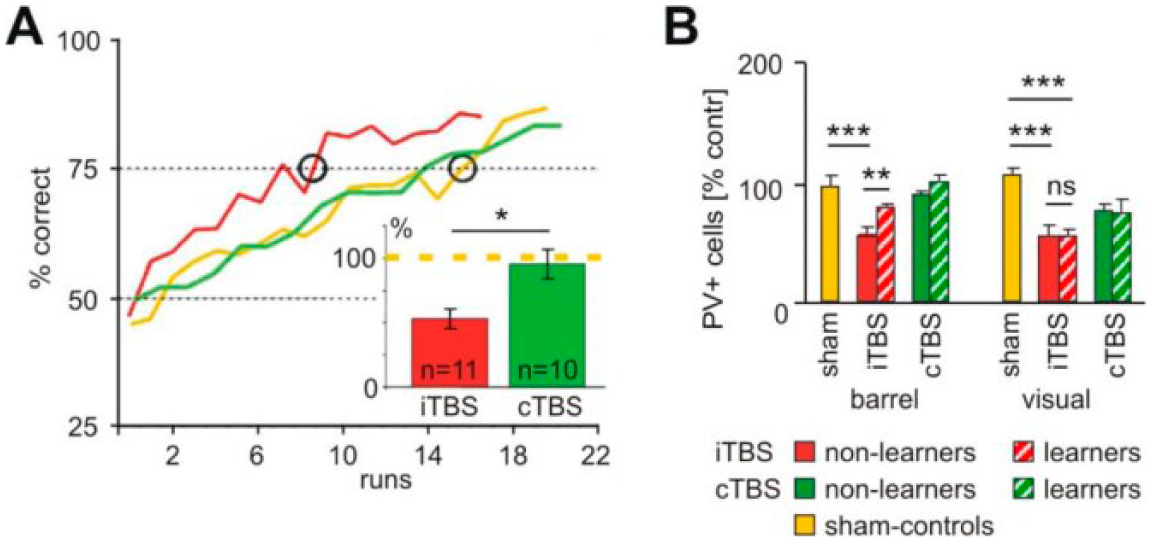
Improved learning performance after intermittent theta burst stimulation (iTBS) associated with modulation of parvalbumin (PV) expression. (A) Rats learned a tactile discrimination task in a radial maze significantly faster when treated with iTBS (red curve) compared with animals receiving continuous TBS (cTBS) (green) or sham stimulation (yellow). (B) The numbers of PV+ cells were strongly reduced in barrel and visual cortex (and other areas) if rats had been treated with iTBS but did not perform the task. However, PV expression increased after learning in task related areas (barrel, motor, frontal cortex) but not in visual cortex while cTBS had no significant effect. The figure has been redrawn on the basis of data published in Mix and others (2010).
Interestingly, PV showed a bidirectional change in expression depending on the combination of iTBS and learning. Animals treated with iTBS but not performing the task (put back to standard cage) showed the typical, about 50% reduction in PV+ cells in frontal, parietal, and occipital cortical areas. However, those rats completing the task showed significantly less reduction in PV expression in those cortical areas expected to be involved in the task (barrel, motor, and frontal cortex) but not in the visual cortex being not essential for the task.
These findings support the notion that modulation of inhibition is a key mechanism to control neuronal plasticity and the ability to learn (Letzkus and others 2015), that changed expression of PV is a sign of plasticity of FS interneurons in the course of learning (Donato and others 2013) and that non-invasive brain stimulation can be used to affect these processes.
Finally, in a recent study using 10 Hz repetitive magnetic stimulation of slice culture preparations direct experimental evidence was provided that repetitive magnetic stimulation reduces inhibitory neurotransmission in a Ca2+ dependent manner, which requires the activation of NMDARs and L-type voltage-gated calcium channel. Interestingly, dendritic but not somatic inhibition was affected 2 to 4 hours after stimulation. This observation adds an additional postsynaptic level of complexity, demonstrating that not only specific inhibitory interneurons are affected but also selected inhibitory postsynapses are modulated in response to repetitive magnetic stimulation (Lenz and others 2016; Lenz and Vlachos 2016).
Together, these studies demonstrate that rTMS is a powerful tool for modulation of cortical excitability through changes in inhibitory networks. It is interesting to speculate that the therapeutic effects of rTMS may in part underlie network disinhibition, which could improve or restore the ability of stimulated networks to express endogenous context-specific plasticity.
Noninvasive Brain Stimulation: A Tool to Assess Cortical GABAergic Neurotransmission in Humans
In line with stimulation protocols used in animal research, cortical GABAergic inhibition can be measured in humans using paired-pulse TMS. For example, short-interval SICI is determined by applying two consecutive TMS pulses at short interstimulus intervals (ISIs) of 1 to 5 ms, with the first conditioning pulse being subthreshold (i.e., not evoking an MEP), but suppressing the MEP induced by the second suprathreshold TMS pulse (Kujirai and others 1993). The ratio of the amplitudes of the conditioned and the unconditioned MEP gives a quantification of the inhibition induced by the conditioning pulse, that is short-interval intracortical inhibition (SICI). In contrast, long-interval intracortical inhibition (LICI) involves the application of two suprathreshold TMS pulses at longer ISIs of 50 to 200 ms; here the first conditioning pulse likewise suppresses the MEP evoked by the second test pulse (Valls-Sole and others 1992). Pharmacological studies indicated that SICI reflects synaptic GABAA receptor dependent inhibition, while LICI reflects activation of metabotropic GABAB receptors (McDonnell and others 2006; Ziemann and others 1996; Ziemann and others 2015). Hence, TMS can be used to determine GABAergic inhibition in the intact human motor cortex noninvasively, and relate level and changes in GABAergic neurotransmission to complex brain functions. This approach has been extensively used in clinical neuroscience in recent years and has provided important insights into the pathophysiological role of GABAergic inhibition in neurological and psychiatric diseases (Berardelli and others 2008; Bunse and others 2014).
However, TMS as a stand-alone technique has only limited capacity to gain insights into higher brain functions. This potential can be significantly enhanced by combining TMS with simultaneous measurements of cortical activity in other electrophysiological or imaging modalities (Ziemann 2011). Registration of electroencephalography during TMS (TMS-EEG) is an interesting tool, which allows to measure TMS effects also in nonmotor brain areas with high temporal and reasonable spatial resolution. Importantly, the development of TMS-compatible EEG amplifiers renders it possible to register TMS-evoked neural excitation at EEG electrodes overlying the stimulation site shortly (<10 ms) after the TMS pulse, and to record its propagation throughout large-scale brain networks for hundreds of milliseconds (Ilmoniemi and others 1997). As the EEG signal is mainly generated by excitatory and inhibitory postsynaptic currents TMS-EEG offers a unique window to noninvasively measure the excitation-inhibition balance in the intact human brain.
Pharmacological studies have shown that an early negative deflection of the TMS-evoked EEG response at around 45 ms after a TMS pulse (N45) reflects GABAA receptor–mediated synaptic inhibition, as it is enhanced by the GABAA receptor agonists alprazolam, diazepam and zolpidem (Fig. 5A and B), whereas a later negative potential at around 100 ms after the TMS pulse (N100) most likely reflects GABAB receptor mediated synaptic inhibition, as it is enhanced by the GABAB receptor agonist baclofen (Premoli and others 2014) (Fig. 5B).
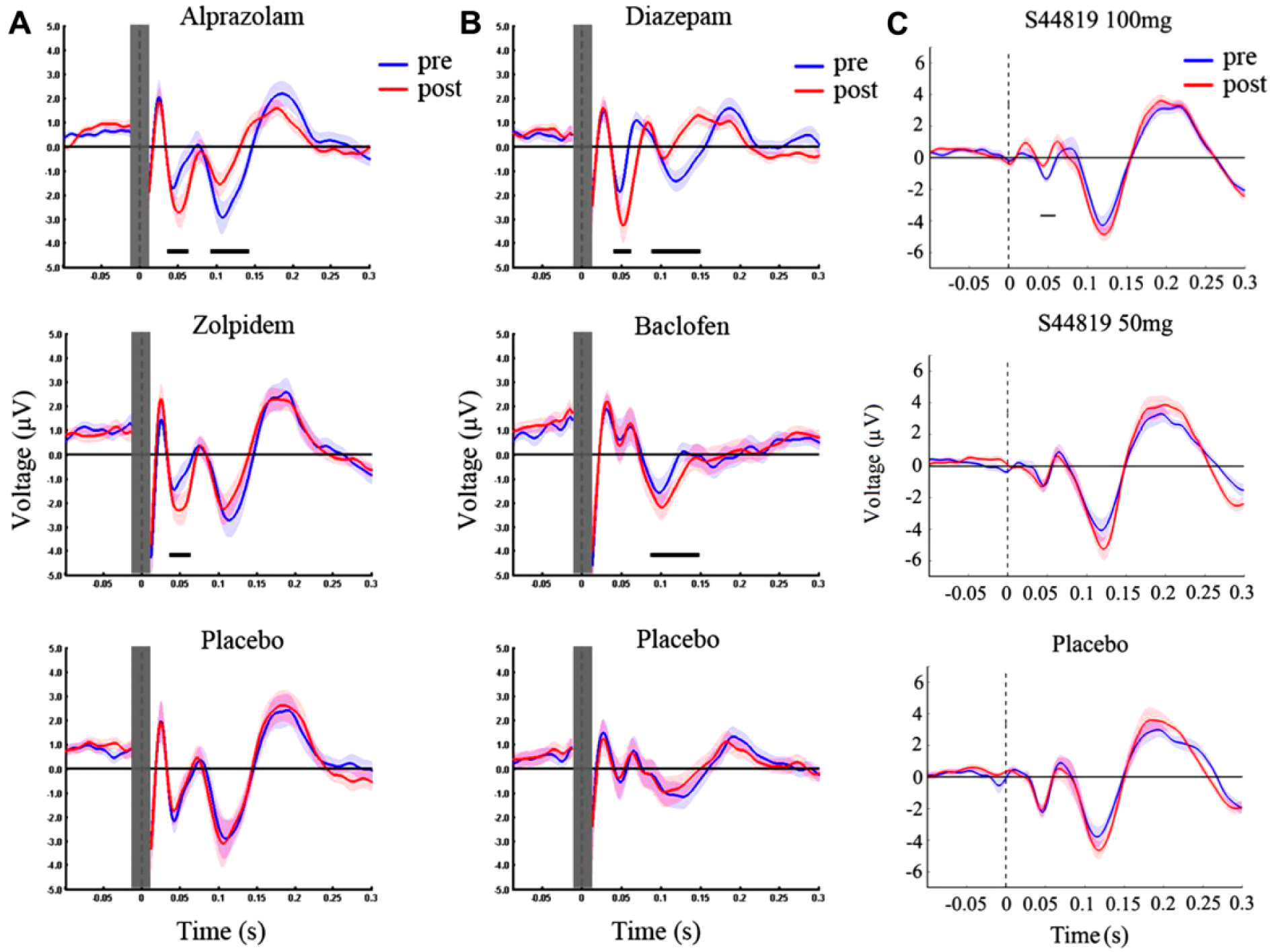
Drug-induced modulation of transcranial magnetic stimulation (TMS)–evoked electroencephalographic (EEG) potentials (TEPs). (A) TEPs elicited by TMS over the primary motor cortex were recorded before (pre, blue) and after (post, red) intake of a single oral dose of alprazolam (1 mg), zolpidem (10 mg), or placebo; (B) before (pre, blue) and after (post, red) intake of a single oral dose of diazepam (20 mg), baclofen (50 mg), or placebo. While alprazolam increased the N45 and reduced the N100 amplitude, zolpidem increased the N45 only. Diazepam increased the N45 and reduced the N100 similar to alprazolam, whereas baclofen increased the N100. Modified from Premoli and others (2014), with permission. (C) A single oral dose of 100 mg, but not 50 mg of S44819, a selective GABAAα5R antagonist, reduced the N45 amplitude (comparison between before [pre, blue] and after [post, red] drug intake). Modified from Darmani and others (2016), with permission. Horizontal bars underneath TEPs denote significant periods of drug-induced changes. Shades represent ±1 SEM.
The potential of these noninvasive markers of cortical GABAergic inhibition for clinical practice is exemplified by one recent study, which showed that the measurement of the N100 in the dorsolateral prefrontal cortex can offer prognostic value for therapeutic interventions in patients with treatment-resistant major depression (Sun and others 2016). In addition, recent work suggests that TMS-evoked EEG potentials may also reflect activity of extrasynaptic GABAAα5 receptors (Darmani and others 2016) (Fig. 5C), which mediate tonic inhibition and may play a crucial role in functional recovery poststroke (Clarkson and others 2010).
Together, these findings disclose that TMS is a promising tool to assess the functional role of GABAergic neurotransmission in human behavior and may help define measures of GABA with diagnostic and prognostic value in patients with neurological and psychiatric diseases.
Measuring GABA In Vivo With Magnetic Resonance Spectroscopy
GABA can be measured in vivo using magnetic resonance spectroscopy (MRS). This MRS measure of GABA is thought to reflect bulk GABA from a large region of interest, and is thought to predominantly reflect cellular, rather than synaptic GABA levels (Rae 2014; Stagg 2014). Functionally, GABA levels are thought to reflect the extent to which a region of interest can exert inhibition (Rae 2014). On this macro level, studies have shown that GABA is associated with brain function as measured with multimodal neuroimaging techniques (Duncan and others 2014). Given the relationship between GABA and brain function, a large number of studies have set out to investigate whether GABA levels, as measured with MRS, are associated with cognitive function in both health and disease.
The in vivo concentration of GABA (1-2 mM) places it at the lower end of the detectable range for MRS. The limited chemical shift separation of 1H-MRS gives an in vivo spectrum where individual signals highly overlapped, and therefore three main approaches are used to reduce signal overlap and detect GABA (Puts and Edden 2012): ultra-high-field MRS (at fields of 7 T and above); two-dimensional (usually J-resolved) MRS; and edited MRS. Of these, edited MRS is the most widely used approach, perhaps largely due to ease of implementation (Mullins and others 2014). Editing the MRS spectrum amounts to reducing the number of signals in the MR spectrum to reduce signal overlap and improve resolution of the GABA signals (Rothman and others 1993).
It is possible to design an edited experiment specifically to reveal GABA signals from underneath larger overlapping signals from more concentrated metabolites. J-difference editing involves acquiring two subexperiments that differ in their treatment of the GABA signals, but which treat creatine signals at 3 ppm the same. Thus, the difference spectrum, calculated by subtracting the results of the two subexperiments, has the creatine signals removed and the GABA signal revealed. In order to edit GABA, frequency-selective editing pulses are applied to GABA signals at 1.9 ppm to manipulate evolution of the GABA signal at 3 ppm, which is detected in the difference spectrum (Fig. 6A).
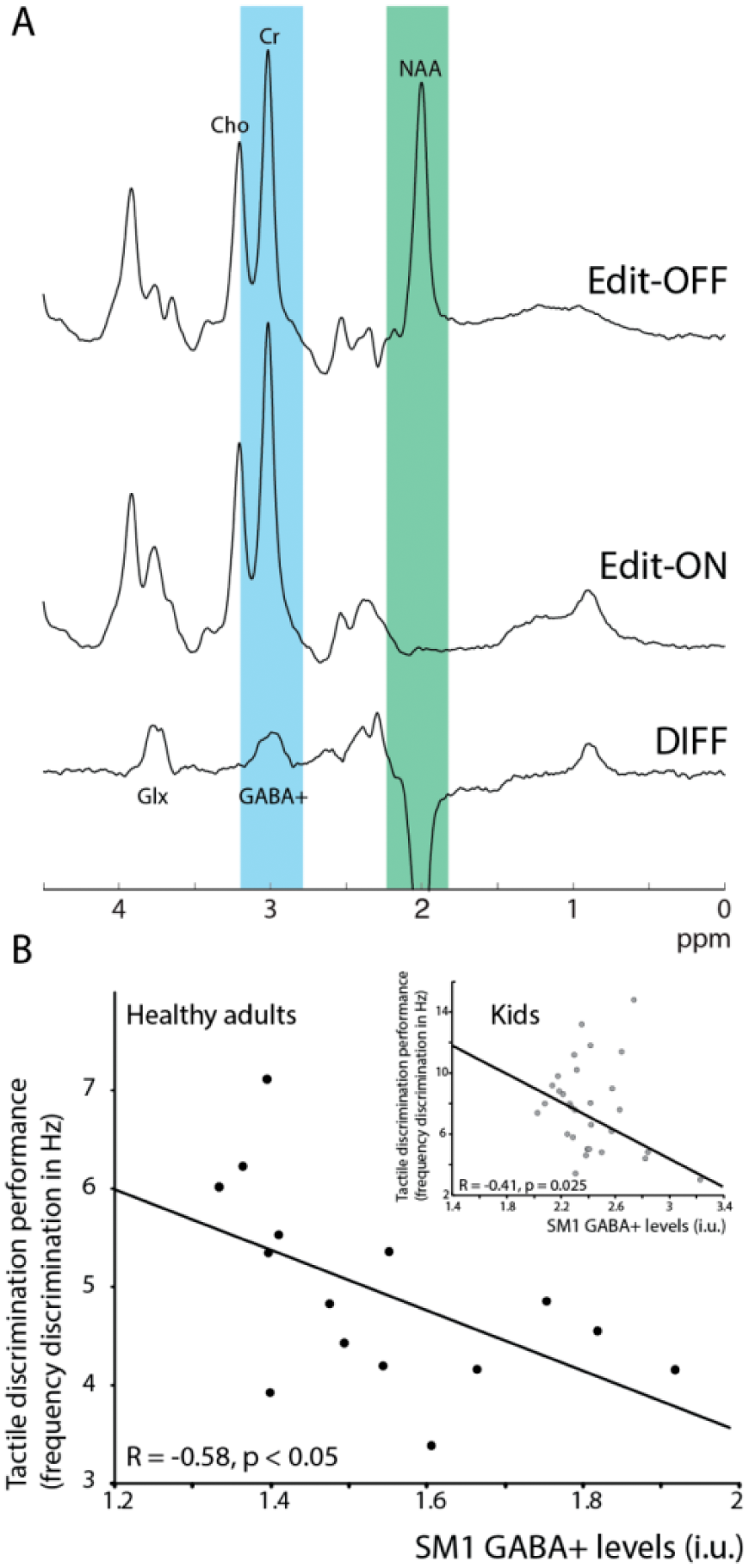
GABA difference editing using MEGA-PRESS. (A) GABA has signals at 1.9, 2.3, and 3 ppm, which are masked by larger concentration metabolites such as N-acetyl aspartate (NAA), creatine, and choline, and is therefore not visible in a typical unedited magnetic resonance (MR) spectrum at 3 T. In MEGA-PRESS, during the Edit-OFF condition, editing pulses are placed off resonance, and signals evolve as normal. In the Edit-ON condition however, editing pulses are placed at 1.9 ppm, selectively affecting the GABA signal at 3 ppm (masked by the larger creatine signal) and suppressing the NAA signal. As the creatine and choline signals were not affected in either condition, but GABA was, the difference spectrum (DIFF) contains only those signals that were affected by the frequency selective editing pulse at 1.9 ppm. The creatine signal is edited out, and a quantifiable GABA signal is now visible at 3 ppm. Because of the low concentration, the edit-OFF and edit-ON conditions are typically repeated 160 times to obtain sufficient signal to noise. (B) Sensorimotor GABA levels correlate negatively with tactile frequency discrimination in healthy adults and children (8-12 years old; inset), showing that participants with more GABA are better at the task. This is reflective of other work showing that GABA levels in a region related to a certain behavioral function are correlated with performance in that behavior in healthy participants. Data were presented in Puts and others (2011) and Puts and others (2017).
Because of the limited sensitivity of MRS and the low concentration of GABA, edited measurements are usually relatively long (~10 minutes) and have a poor spatial resolution (~27 mL measurement volume). The most widely used implementation of editing is MEGA-PRESS (Mescher and others 1998).
It is a major limitation that edited MRS does not only edit GABA signals—the edited signal at 3 ppm contains contributions from other GABA-like metabolites such as homocarnosine and a substantial contribution from macromolecules (MM). Co-editing of MM signal occurs because there is an MM signal at 1.7 ppm (close to the GABA signal at 1.9 ppm that editing pulses are applied to) that is coupled to an MM signal at 3 ppm (underneath the edited GABA signal). Thus, MM co-edits with GABA, and the edited GABA signal is usually referred to as GABA+. MM-suppressed editing schemes have been proposed to address this issue (Henry and others 2001), but they suffer from increased variance due to scanner instability and subject motion (Edden and others 2016).
J-difference editing with MEGA-PRESS is an inherently wasteful technique, aimed at acquiring edited signal from just one metabolite (GABA) in one brain region at a time. The long duration of measurements makes this inefficiency particularly stark, and it is not uncommon for an hour-long MRI protocol to only make GABA measurements from three brain regions. There is therefore strong interest in improving the acquisition efficiency (or temporal signal-to-noise ratio) of measurements by multiplexing editing of multiple metabolites and/or multiple regions.
Hadamard Encoding and Reconstruction of MEGA-Edited Spectroscopy (HERMES) performs MEGA-edited measurements of more than one metabolite at the same time, by using orthogonally applied editing timings for each metabolite (Chan and others 2016). For example, whereas the GABA-encoding across averages might be ON-OFF-ON-OFF . . ., an alternative editing target (Harris and others 2017) such as glutathione (GSH) might be encoded as ON-ON-OFF-OFF. Thus, the GABA-OFF scans can be subtracted from the GABA-ON scans to give the GABA-edited spectrum and the GSH-OFF scans can be subtracted from the GSH-ON scans to give the GSH-edited spectrum without compromise of the editing selectivity (Saleh and others 2016). Simultaneous editing of GABA in more than one voxel has also been implemented using parallel reconstruction (Oeltzschner and others 2017) and spatially selective Hadamard editing (Chan and others 2017). The ability to acquire more information within the same amount of time will change the types of studies that can be planned in the future, and allows for the investigation of relationships within and between biochemical systems.
Magnetic Resonance Spectroscopy of GABA and Cognition
GABA plays an important role in regulating brain activity and shaping the neuronal responses to incoming sensory input. As such, GABA is also critically involved in encoding sensory information. Using MRS of GABA, it has been shown that individual differences in occipital GABA levels are correlated with healthy participants’ ability to discriminate visual orientation, linking known animal work to human perception (Duncan and others 2014). Similarly, sensorimotor GABA levels were correlated with healthy participants’ ability to discriminate tactile frequencies (Duncan and others 2014; Puts and others 2011), a finding later replicated in typically developing children, but not in children with neurodevelopmental disorders (Puts and others 2017) (Fig. 6B).
Several other studies have shown links between GABA and cognition, including studies linking dorsolateral prefrontal cortex GABA to working memory (Duncan and others 2014; Yoon and others 2016) and impulsivity (Boy and others 2011; Yoon and others 2016), and supplementary motor area GABA levels to motor distraction (Duncan and others 2014; Mullins and others 2014). Other studies have found that GABA levels change with age, and reductions in GABA with age have been linked to alterations in cognitive processing (Harris and others 2017; Porges and others 2017). One key observation across these various studies is that MRS measurements of GABA are regionally and functionally specific. Furthermore, interindividual differences in baseline GABA levels from a region relevant to a certain task can predict behavioral performance on that task. Interestingly, and perhaps surprisingly, these macro-level findings often reflect animal/cellular metrics of GABA function, providing a link between micro-level biochemistry and macro-level behavioral function (Duncan and others 2014).
While the aforementioned studies focus on baseline GABA levels—that is, GABA levels measured at rest—other studies have investigated GABA under conditions of plasticity. In one of the first studies applying MRS of GABA, it could be shown that during an ischemic block, motor cortex (M1) GABA levels significantly decreased as a function of time, reverting after ischemic block was released, possibly reflecting a release of inhibition to allow for plastic changes to take place (Levy and others 2002). Floyer-lea and colleagues (2006) showed that M1 GABA levels decrease during a motor learning task. In several studies, it has been shown that GABA levels can be altered using non-invasive brain stimulation; concurrent TMS over M1 increases GABA levels, whereas both anodal and cathodal transcranial direct current stimulation reduces GABA levels (Stagg 2014). Finally, it could be demonstrated that baseline GABA levels in the sensorimotor cortex (preintervention) predicted performance improvement on a two-point discrimination task after repetitive electrical stimulation of the fingers, interestingly individual GABA levels changed in a homeostatic manner rather than on a group-average level (Heba and others 2016).
There is an increasing interest in studying GABA levels in disease (Puts and Edden 2012), for example, in neurodevelopmental disorders, neurodegenerative diseases and chronic pain. Previous work has suggested altered GABAergic function in autism spectrum disorder (ASD), attention deficit hyperactivity disorder (ADHD), and Tourette syndrome (TS). MRS studies of GABA in these disorders corroborate this previous work, showing reduced GABA levels in children with ASD and ADHD (for reviews, see Ford and Crewther 2016; Schür and others 2016) although this may be region-dependent (occipital GABA appears unaffected in ASD). Although altered GABA has been shown in TS (Draper and others 2014; Freed and others 2016; Puts and others 2015), the work is inconsistent, with some studies showing increased and others decreased GABA levels. These differences might be related to regional cortical differences, cohorts studied, or methodological considerations, and are perhaps more difficult to interpret.
Linking MRS to brain function and cognition has limitations. Associations between GABA levels and cognitive and behavioral measurements at rest or after learning, that are present in healthy participants, are not always easy to interpret and may not be present in disease. Interpretation of the functional relevance of the presence, or absence of these associations is often based on links with cellular and animal work. Further studies are necessary to determine the link between MRS of GABA and inhibitory processing on a mechanistic level, which may be particularly relevant for studying the GABA system in disease. Furthermore, findings are not always replicated (Cousijn and others 2014; Harris and others 2015), limiting the reproducibility and interpretability of these studies and are likely due to experimental and biological effects, such as small cohort size and differences in cohorts. In summary, MRS of GABA allows for functionally relevant in vivo measurements of the role that GABA plays in driving behavior and cognition in health and disease, and is an important contribution to the study of GABA in health and disease.
Neurochemical Imaging of Chronic Pain
Over the past 15 years, the application of MRS in the investigation of chronic pain has gained traction and recent studies have suggested that GABA and glutamate are strong correlates of the subjective pain experience. In healthy participants, thalamic GLX (combined glutamate and glutamine) levels predicted pain thresholds (Zunhammer and others 2016). MRS has also been applied to study GABA and GLX levels in patients with chronic pain. GLX levels have been shown to be elevated in fibromyalgia patients within the posterior insula (Harris and others 2009). Elevated GLX levels were associated with evoked pain sensitivity, suggesting that glutamatergic activity in this region of the brain might be in part responsible for the “gain setting” on central neural pain processing. In a longitudinal trial of acupuncture and sham acupuncture, it could be demonstrated that changes in GLX levels, specifically within the posterior insula cortex, tracked with changes in both experimental and clinical pain (Harris and others 2008). In that study, reductions in GLX were associated with greater improvements in both clinical and experimental pain. Importantly, changes in insular GLX were also associated with concomitant changes in the brain’s functional response to evoked pressure pain during fMRI, suggesting that brain GLX levels may be associated with neuronal activity. As such increased GLX levels could enhance excitatory neurotransmission and subsequent pain. Interestingly increased GLX levels have been reported in animal models and have been proposed in fibromyalgia (Zhuo 2008). On the other hand, GABA levels were shown to be reduced in fibromyalgia patients within the anterior insula but not in the posterior insula (Foerster and others 2012) (Fig. 7A and B).
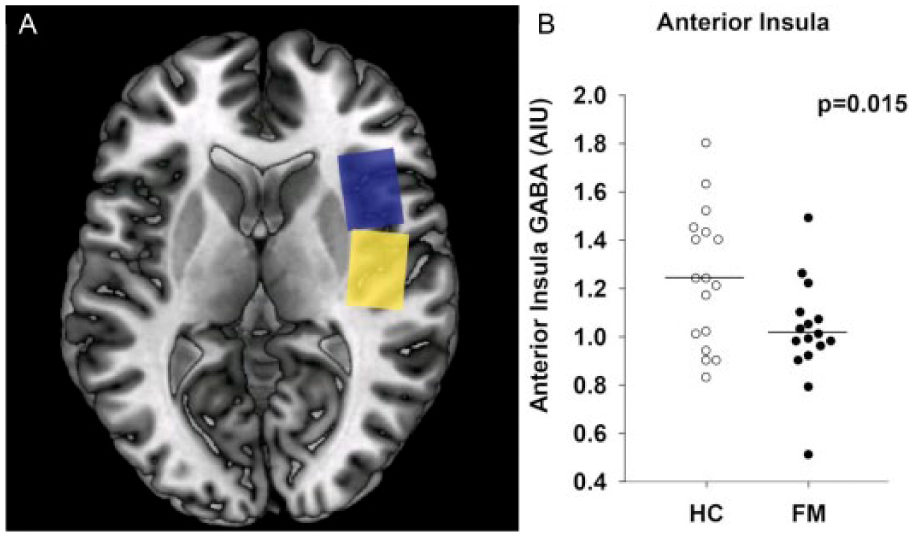
GABA voxel placement in the anterior and posterior insular cortex. (A) Axial T1-weighted brain image showing single-voxel placement in the anterior insula (blue box) and the posterior insula (yellow box). (B) Levels of GABA within the right anterior insula of individual patients with fibromyalgia (FM; solid circles) as compared with healthy controls (HC; open circles). Fibromyalgia patients have reduced concentrations of GABA in the right anterior insula. There were no between-group differences in the right posterior insula or occipital cortex (data not shown). Circles represent individual subjects; horizontal bars indicate the mean. AIU = arbitrary institutional units.
Importantly, lower GABA levels within the posterior insula were associated with greater pain sensitivity. In patients with chronic neuropathic pain after spinal cord injury GABA levels within the thalamus were shown to be decreased as compared with healthy controls and patients after spinal cord injury, but without pain (Gustin and others 2014), suggesting that thalamic disinhibition might play a role in the genesis of neuropathic pain. In patients with osteoarthritis pain, it was inversely correlated with GABA in the anterior cingulate cortex (Reckziegel and others 2016). In aggregate, these findings suggest a neurotransmitter imbalance within regions critically involved in pain processing, in terms of increased glutamate and decreased GABA levels. Pharmacologic studies using neurochemical imaging derived outcomes have also been reported in chronic pain. Preclinical work with pregabalin, an efficacious drug for fibromyalgia, suggested that the mechanism of action of this compound was to reduce the release of glutamate into the synaptic cleft (Micheva and others 2006). Of note, an MRS study found that pregabalin reduced GLX levels in the posterior insula and higher pretreatment levels of GLX in this region were associated with greater subsequent reductions in experimental pressure pain sensitivity (Harris and others 2013).
Future studies in chronic pain will need to better link neuroplastic brain changes with both pain and nonpain clinically relevant outcomes. Many chronic pain patients report fatigue, cognitive deficits, mood disturbance, and poor sleep and these outcomes should also be explored with MRS.
Conclusion
In this review article, we sought to bring together some important aspects of the role and function of the main inhibitory neurotransmitter GABA from researchers in different sub-fields, working at different levels of neural organization and using different methods to investigate GABAergic systems, that is, tetrode recordings, TMS, EPs, EEG, and MRS. Linking the function of specific cell populations and/or neurotransmitter systems to perception, cognition and behavior is one of the main goals and at the same time greatest challenges of modern neuroscience. Different methods reveal different aspects of what we hypothesize to be the same system, yet on different scales of complexity and spatial resolution. Scientific progress very much depends on the range and accuracy of the methods applied, and methodological specialization allows for a high degree of focus within specific subfields. The downside of specialization is that scientists with a common interest, for example, on GABA neuroscience, yet from different backgrounds, risk losing (or not even establishing) contact. As such the distinction between different subtypes of GABAergic interneurons is currently of little relevance to MR spectroscopists. Likewise, the poor resolution of GABA spectroscopy combined with the lack of site specificity (intracellular, extracellular, etc.) leaves the basic scientist somewhat puzzled.
Among researchers (methodological) reductionism stating that entire systems can be explained in terms of their constituent parts and their interactions is common. As such there is the basic assumption that findings in single cells and small cell assemblies translate not only into local but also into far-ranging, large-scale networks, which finally help to explain higher cognitive functions. It is an epistemological, rather than a scientific question as to what actually defines complexity, for example, the number of neurons and their interactions, the method applied and its outcome parameters, or the hypothesis to be verified (or falsified). As such the term complexity does not necessarily refer to the world per se, but to the way we formulate and address a specific question. It will be of critical importance to integrate results yielded by a specific method into a conceptual framework that is accessible by researchers from other subfields. In the case of GABA, one candidate topic to provide such a conceptual framework is oscillatory activity. In the mature brain, GABA strongly shapes cortical gamma and hippocampal theta oscillations. These oscillations have the potential to be the heuristic backbone to both link microcircuitry to larger networks, accessible to EEG, EP, and MRS, and also to link neurophysiology to behavior. Furthermore, computational neuroscience holds promise to generate common denominators and even models that have the potential to bridge these gaps. However, an approach that could be called “integrative neuroscience” is probably needed to fully understand how a four-carbon molecule shapes complex behavior.
Footnotes
Acknowledgements
The current article was inspired by a conference on the role of GABA on different levels of neural organization and complexity: GABA—From Molecule to Cognition, held at the Ruhr University of Bochum in June 2016 and supported by the SFB 874 Integration and Representation of Sensory Processes. We thank Niklas Wulms for technical support and valuable discussion.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Tobias Schmidt-Wilcke currently receives funding from the SFB 874 (to Project A8).