
Abstract
Scalable purification platforms have been developed for adeno-associated virus (AAV) processing to support large-scale vector manufacturing. The ability of column chromatography to recover packaged vectors and remove empty capsids has been well-established, but knowledge gaps remain for understanding process parameter impacts on impurity retention. In this work, we examine the impacts of two key process parameters—the harvest method and affinity resin selection—on residual host cell protein (HCP) retention. Sequential window acquisition of all theoretical fragment ion mass spectra (SWATH-MS) proteomics is applied to comprehensively profile residual HCPs in affinity chromatography (AC) elution pools from four AAV serotypes (AAV2, −5, −8, and −9) produced by suspension HEK293 cells. Vectors were purified from cell culture lysates and from supernatants using POROS™ CaptureSelect™ AAVX (AAVX) and one additional serotype-specific affinity resin—Capto™ AVB (AVB), POROS CaptureSelect AAV8 (PAAV8), or POROS CaptureSelect AAV9 (PAAV9). Significant divergence in residual HCP profiles was observed with the use of different affinity resins, with AVB and PAAV9 showing reduced residual HCP content in elution pools compared with AAVX and PAAV8. Processing of null culture lysates with fresh resins and resins with digested single-domain antibody fragments (sdAbs) shows that differences in resin performance are driven by variable nonspecific sdAb association with cellular impurities. Proteomic analysis of vector preparations from lysates compared with supernatants demonstrates product quality advantages of designing a media-only harvest process, specifically for AAV8, which was measured to contain an average of 66% of total vector genome content in the cell culture media. This work highlights the importance of serotype-specific tailoring of AAV downstream process design for improved product quality attributes to support clinical manufacture and small-scale analytics workflows.
Keywords
INTRODUCTION
Recombinant adeno-associated virus (AAV) is the most prevalent in vivo gene therapy platform for delivery of curative treatments to patients suffering from genetic diseases. 1 The safety and long-term efficacy of AAV therapies have been well established with over 200 completed or in-progress clinical trials and seven FDA-approved products. 2 One major hurdle impacting the availability and cost of AAV therapies is the difficulty in scaling up current manufacturing platforms.3,4 Recent advances in HEK293 suspension culture systems and the advent of stable packaging and producer cell lines have greatly improved vector production capacity.5–7 These higher-titer and larger-volume production systems require new purification methods for recovery of packaged capsids, removal of empty capsids, and robust clearance of process- and product-related impurities.8,9 Affinity chromatography (AC) has emerged as a preferred method for scalable primary capture of AAV capsids with the development of heparin sulfate,10,11 immunoaffinity, 12 and peptide ligands. 13 These AC processes can easily be scaled up and fit into multistage chromatography platforms for vector recovery and full capsid enrichment.
Despite extensive demonstrations of column chromatography for AAV purification, there remains a significant gap in the understanding of process parameter impacts on critical quality attributes (CQAs) for these systems. One CQA for AAV purification is residual host cell protein (HCP) content, which can impact transduction efficiency and may pose additional risks to product stability and safety.14–16 In this work, we use sequential window acquisition of all theoretical fragment ion mass spectra (SWATH-MS) proteomics to examine the residual HCP impacts of two key parameters in scalable AAV purification systems—the culture harvest method and the AC resin selection. Four AAV serotypes produced from transient transfection of suspension HEK293 cell culture were split at harvest into lysate and supernatant lots prior to affinity purification. AC was performed using POROS CaptureSelect AAVX (AAVX) in addition to one serotype-specific affinity resin for each material lot. Significant divergence in residual HCP profiles was observed based on affinity resin and harvest feed stream, suggesting that serotype-specific tailoring of purification can offer notable advantages for reducing residual impurity levels. Overall, the results of these experiments inform downstream process design for scalable AAV systems and highlight the importance of product-specific purification development for the improvement of both analytical workflows and large-scale vector production.
MATERIALS AND METHODS
AAV production
AAV vectors were produced by transient transfection of EXPI293™ cells (Thermo Fisher Scientific) grown in suspension culture with EXPI293™ Expression Medium (Thermo Fisher Scientific). Transfections were performed as described in Leibiger et al. 17 Transfection and cell culture process parameters are summarized in Supplementary Table S1. Two different vectors—one packaged with enhanced green fluorescent protein (EGFP) transgenes (AAV-EGFP) and one empty capsid lot (AAV-Empty)—were produced for AAV2, −5, −8, and −9, respectively. 2 × 1 L flasks were transfected for AAV-EGFP lots of each AAV serotype, and 1 × 1 L flask was transfected for AAV-Empty lots of each serotype using delivery of pRepCap and pHelper only. An additional 1 × 1 L flask was transfected with only delivery of the pEGFP plasmid for production of “EGFP control” material that had no AAV production or viral helper genes delivered to cells. 17 The transfection procedures were performed twice for generation of biological duplicate lots. 17 In total, 26 × 1 L flasks were transfected across the experiment (two biological replicates of: eight AAV-EGFP, four AAV-Empty, and one EGFP control).
AAV purification
AAV lots were harvested 72-h post transfection using methods and equipment described in Leibiger et al. 17 Shake flasks (N = 2) were pooled for AAV-EGFP lots at harvest within each biological replicate. Supernatant and lysate lots were separated for AAV-EGFP harvest materials for AAV5, −8, and −9 to compare purification performance from each harvest stream. AAV2-EGFP supernatant was not purified due to a low vector genome (VG) titer measurement. A detailed experimental workflow is visualized in Fig. 1.
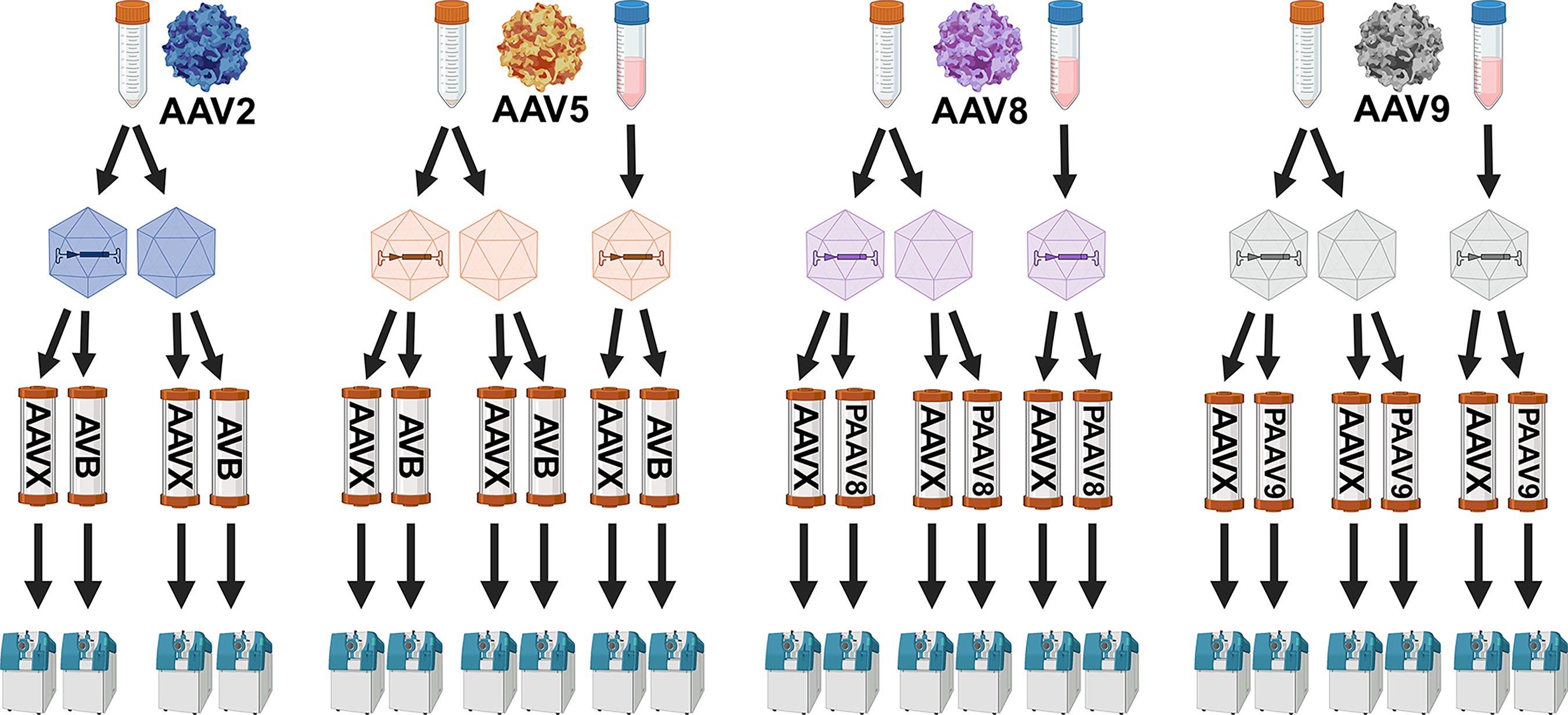
Downstream experimental workflow. Two different vector types were produced for each AAV serotype—one packaged with EGFP transgenes and one empty vector lot. Lysates from each of the lots were purified using POROS™ CaptureSelect™ AAVX (AAVX) resin and one additional affinity resin—Capto™ AVB (AVB), POROS CaptureSelect AAV8 (PAAV8), or POROS CaptureSelect AAV9 (PAAV9). Supernatants from the EGFP-packaged vector lots of AAV5, −8, and −9 were also purified using AAVX and one additional serotype-specific affinity resin. Eluates from each purification cycle were characterized using SWATH-MS. SWATH-MS, sequential window acquisition of all theoretical fragment ion mass spectra. AAV, adeno-associated virus.
Affinity purification was performed using the same “base-case” AC process described in Leibiger et al., 17 except the workflow was expanded for purification using different affinity resins and different load materials. AAV2-EGFP-lysate and AAV2-Empty-lysate lots were purified using AVB and AAVX resins. AAV5-EGFP-lysate, AAV5-Empty-lysate, and AAV5-EGFP-supernatant lots were purified using AVB and AAVX resins. AAV8-EGFP-lysate, AAV8-Empty-lysate, and AAV8-EGFP-supernatant lots were purified using PAAV8 and AAVX resins. AAV9-EGFP-lysate, AAV9-Empty-lysate, and AAV9-EGFP-supernatant lots were purified using PAAV9 and AAVX resins. EGFP control lots were purified using each of the four affinity resins. Purification of lysates was performed as described previously. 17 Supernatants were 2-fold concentrated using AMICON Ultra-15 10 kDa spin filters (Sigma) prior to loading. Lysates and supernatants for each serotype replicate group were loaded to the two different affinity resins from the same pool, with load amounts for each cycle corresponding to 100 mL of culture volume. This setup allowed for controlled capsid loading within each replicate group; however, capsid loading amounts varied across serotypes based on cell culture productivities. Capsid loading is reported for each purification cycle (Supplementary Table S2).
Immunoaffinity resin screening—fresh versus Proteinase K-treated resin
The same base-case affinity process was applied with the EGFP lysate load containing no AAV capsids for 1 mL of fresh resin and 1 mL of resin that was subjected to Proteinase K (Thermo Fisher Scientific) treatment. For Proteinase K treatment, 1 mL of each fresh resin (AVB, AAVX, PAAV8, and PAAV9) was washed twice by centrifugation at 3,000 RCF, followed by decanting and resuspension of resin pellets in 4 mL equilibration buffer (20 mM Tris-HCl, 0.1 M NaCl, pH 7.2). Resins in 4 mL equilibration buffer were treated with 150 µL of 20 mg/mL Proteinase K (Thermo Fisher Scientific) and incubated in an Eppendorf ThermoMixer C at 1,000 RPM, 56°C for 16 h. Two additional washing cycles were performed prior to the use of ligand-digested resins.
AAV analytical characterization
Biolayer interferometry (BLI) using AAVX biosensors with an Octet R8 (Sartorius) instrument was performed for each load and elution material to calculate capsid recoveries. Methods for BLI data collection and analysis are described in Leibiger et al. 18 Elution pools were measured for protein concentration using Coomassie Plus (Bradford) Assay Reagent (Thermo Fisher) with an albumin standard curve (Thermo Fisher) as previously described. 17 SDS-PAGE and negative stain transmission electron microscopy (nsTEM) were performed using methods, reagents, and equipment detailed in Leibiger et al. 17
Liquid chromatography-tandem mass spectrometry sample preparation
Briefly, 50 µg protein for each sample was buffer exchanged into 100 µL of 100 mM triethylammonium bicarbonate and was digested as previously detailed. 17 Reduction was performed with the addition of 2.5 µL 100 mM Tris(2-carboxyethyl)phosphine (Thermo Fisher Scientific) followed by 1 h incubation at 60°C. Alkylation was achieved with the addition of 5 µL 150 mM iodoacetamide (Sigma-Aldrich) and 30 min incubation in the dark. Digestion with sequencing-grade trypsin (Promega) was carried out at 37°C for 16 h at an enzyme-to-substrate ratio of 1:50. Digestion steps were scaled accordingly for samples with less than 50 µg total protein available. Sample cleanup was performed as previously detailed using OMIX C18 tips (Agilent), and eluates were dried with a SpeedVac (Thermo Fisher Scientific). 17
Liquid chromatography-tandem mass spectrometry data acquisition and processing
Dried samples were resuspended in 2% acetonitrile (ACN) and 0.1% formic acid (FA). Samples for triplicate liquid chromatography-tandem mass spectrometry (LC-MS/MS) injection were spiked with a target of 5 fmol/µL pre-digested yeast alcohol dehydrogenase (ADH), and samples equivalent to 5 µg digested protein were injected for each LC-MS/MS analysis. Then, 0.25 µL of retention time standard (Biognosys) was added to each injection. LC-MS/MS was performed as described previously using a TripleTOF 6600 (Sciex), Eksigent Nano LC 425, and ChromXP C18CL column (3 mm, 120 Å, 150 × 0.3 mm; Sciex). 17 Mobile phases of 0.1% FA in water (A) and 0.1% FA in ACN (B) were used with a program of 3–25% mobile phase B across 68 min, 25 − 35% mobile phase B across 5 min, 35 − 80% mobile phase B across 1 min, and 80% mobile phase B for 3 min. A flow rate of 5 µL/min was used.
Data-dependent acquisition (DDA) was performed as previously described for spectral library construction. 17 Triplicate DDA data were acquired for all biological replicate R2 samples (N = 26) except for five samples that were run in duplicate and one sample that was run in one injection due to protein limitations (Supplementary Table S3). DIA SWATH-MS was performed for all samples (N = 52) as previously described with an MS1 full scan followed by 64 MS/MS variable size window acquisitions. 17 Triplicate SWATH-MS injections were performed for all samples except for two samples that were run in duplicate and five samples that were run as single injections due to protein limitations (Supplementary Table S3).
Database searches were carried out as previously detailed using ProteinPilot version 5.1 (Sciex) and a local copy of the NCBI:Hu_RefSeqGRCh38 database. 17 A ProteinPilot group file was imported to Skyline (version 20.2.0.343) for consolidated library construction using BiblioSpec with a 95% peptide identification confidence threshold.17,19 SWATH-MS data extraction was carried out as previously described. 17 Peak integrations were performed in MSstats (version 4.8.0). Protein peak areas were normalized against ADH to calculate relative HCP amounts (ng) in each sample with the assumption that all proteins yield a consistent response. Protein identification in each sample required peptide detection (q < 0.01) in all replicate SWATH-MS data acquisitions.
RESULTS
AC capsid yields across the sample set ranged from 42.93% (AAV2-EGFP-lysate) to 98.61% (AAV8-EGFP-sup; median = 81.08%; Fig. 2, Supplementary Table S2, and Supplementary Fig. S1). Paired analyses of capsid yield data showed no statistically significant differences between recoveries for serotype-specific resins compared with the AAVX resin (Wilcoxon test, p > 0.05). Additionally, no significant differences were measured for capsid recoveries from the empty capsid lot compared with the EGFP-packaged lot and for material processed from lysate (EGFP-lysate) compared with material purified from supernatant (EGFP-sup; Wilcoxon test, p > 0.05).
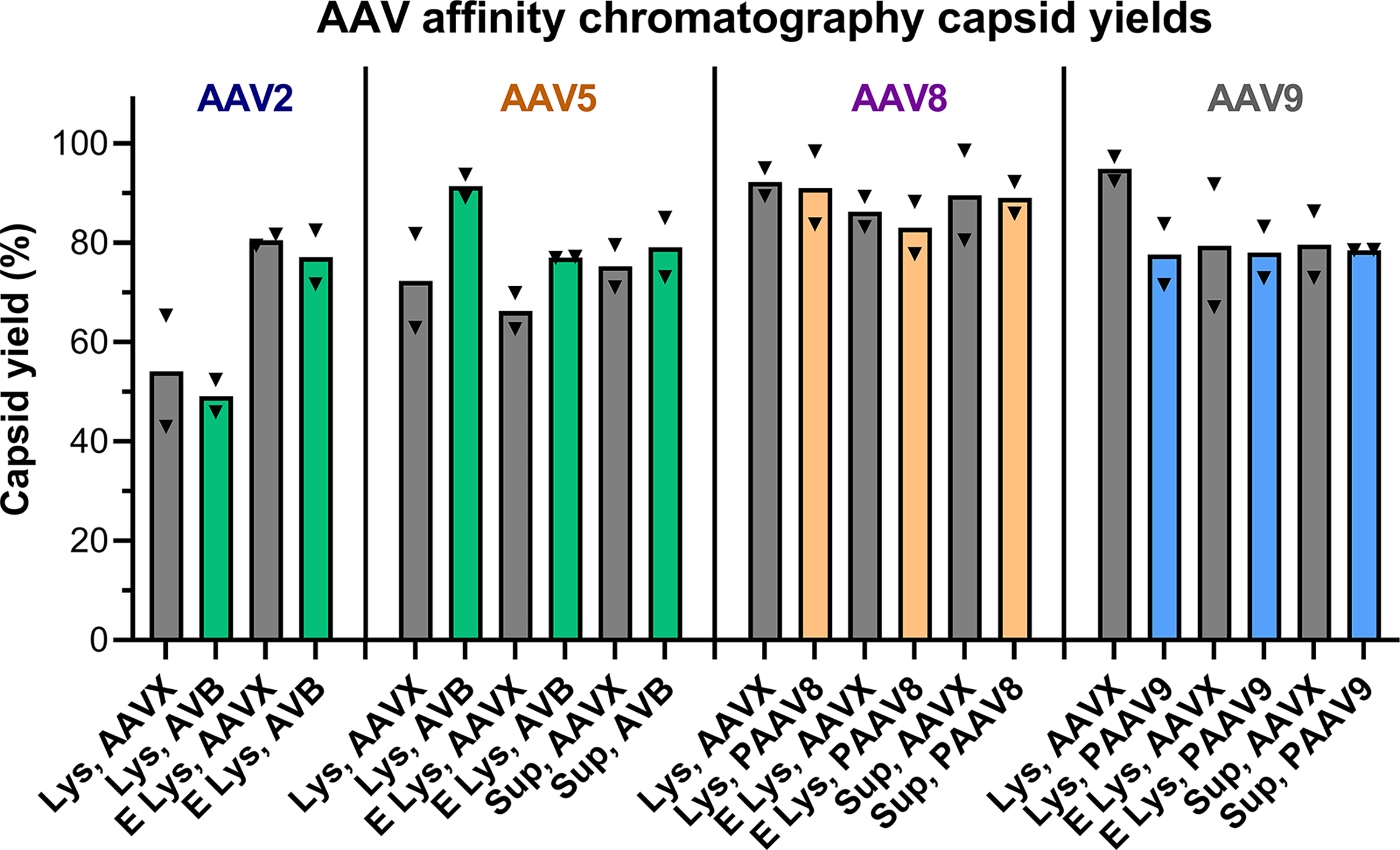
Affinity chromatography capsid yields measured by AAVX-Octet for all cycles, including AAV-EGFP lysates (Lys), AAV-Empty lysates (E Lys), and AAV-EGFP supernatants (Sup). Gray bars correspond to purification by POROS CaptureSelect AAVX (AAVX). Green bars correspond to purification by Capto AVB (AVB). Orange bars correspond to purification by POROS CaptureSelect AAV8 (PAAV8). Blue bars correspond to purification by POROS CaptureSelect AAV9 (PAAV9). Replicate cycles are shown by triangles with bar heights equal to mean capsid recovery.
Harvest material impacts on HCP retention
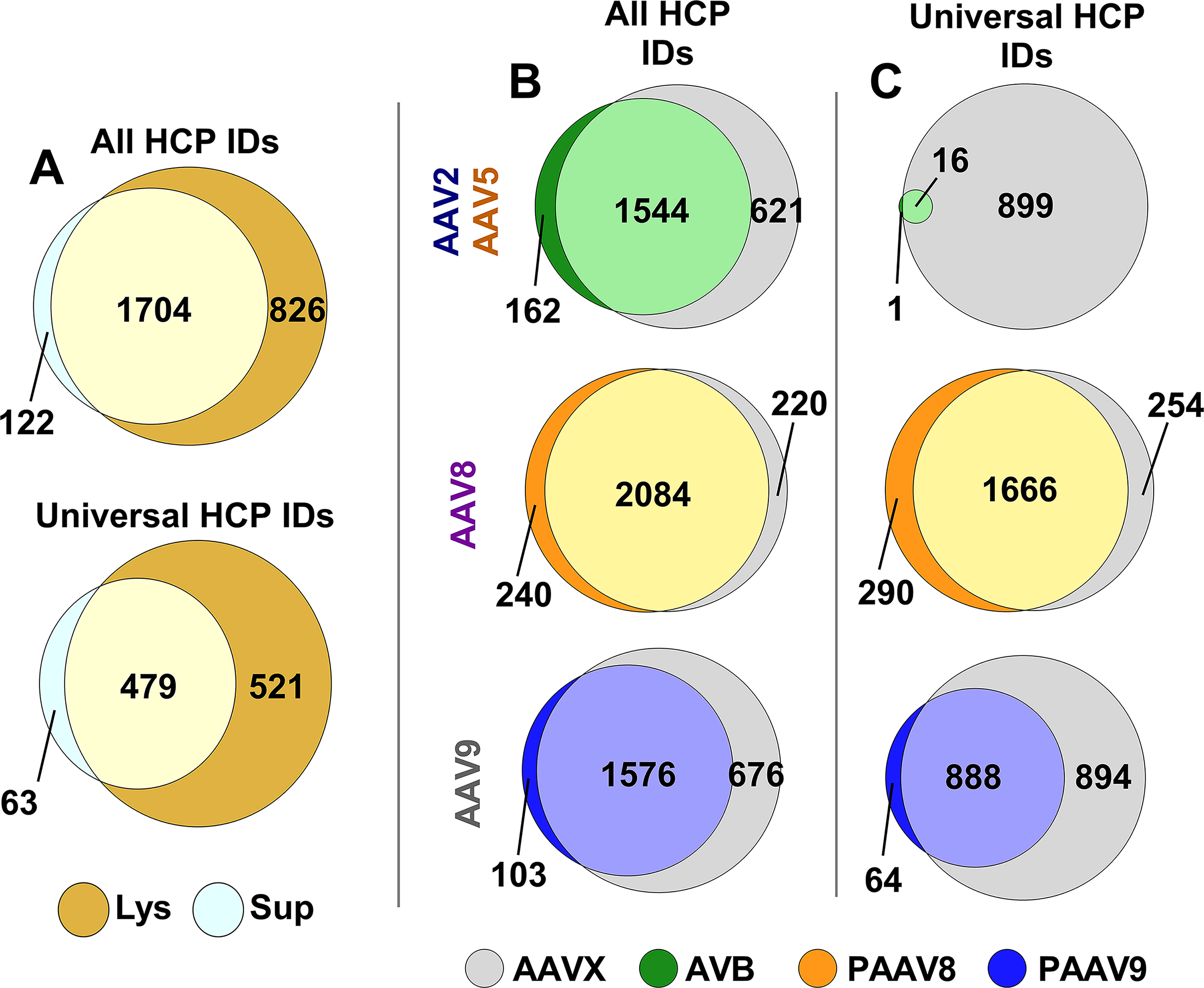
VG titer measured in culture lysates compared with culture supernatants varied by AAV serotype (Supplementary Fig. S2). AAV2-EGFP supernatant was measured to contain less than 1% of the total VG titer for all cultures (N = 4). AAV8-EGFP supernatant contained an average of 66% of total VG titer measured from the cultures (range of 63 − 68% for N = 4 cultures). AAV5-EGFP and AAV9-EGFP showed intermediate levels of VG titer in culture supernatants, averaging 33.9% and 30.2% of total VGs, respectively (N = 4 each). There was a greater number of total and universal HCP identifications for material processed from lysate compared with supernatant (Fig. 3A). Purity visualization with SDS-PAGE shows material harvested from culture supernatant for AAV5, −8, and −9 containing fewer impurities compared with lysate-harvested material from the same flasks and purified with the same resins (Fig. 4). Divergence in vector purity after AC is further shown by nsTEM images where AAV8-EGFP harvested from the culture supernatant contains less debris compared with lysate-harvested material after both lots were subjected to the same AAVX purification process (Fig. 5A). AAVX purification conditions were used as a reference to identify material-specific residual HCPs that appeared in all elution pools for one load material type but are absent in all elution pools for the other material type (Supplementary Table S4). There were 347 residual HCPs identified in all samples after lysate purification that did not appear in any supernatant-purified samples. Comparatively, 11 residual HCPs were identified in all samples after supernatant purification that did not appear in any lysate-purified samples.

SYPRO Ruby-stained protein gels of AAV2 and AAV5 (gel 1, left) and AAV8 and AAV9 (gel 2, right) samples purified using affinity chromatography. Eluates from the purification of AAV-EGFP lysates (Lys) and AAV-Empty lysates (E Lys) are shown for each serotype. Eluates from purification of AAV-EGFP supernatants (Sup) are shown for AAV5, −8, and −9 and are marked with a red S. Gel lanes are colored at the top by affinity resin, with gray rectangles corresponding to POROS CaptureSelect AAVX (AAVX), green rectangles corresponding to Capto AVB (AVB), orange rectangles corresponding to POROS CaptureSelect AAV8 (PAAV8), and blue rectangles corresponding to POROS CaptureSelect AAV9 (PAAV9). Biological replicate R1 samples are shown for all gel lanes.

Resin impacts on HCP retention
The percentage of total protein composed of HCPs in the eluates for the same load materials was used as a resin performance metric, which showed that affinity resin selection has a strong influence on vector purity (Supplementary Figs. S3 and S4). Purification of AAV2 and AAV5 with AVB resin gave a significant reduction in residual HCP content compared with the AAVX resin (Wilcoxon test, p < 0.05). Similarly, AAV9 purification with PAAV9 resin showed a significant reduction in relative HCP content versus AAVX resin (Wilcoxon, p < 0.05). Interestingly, there was no difference observed between HCP retention with PAAV8 resin compared with AAVX resin for AAV8 purification. SDS-PAGE and nsTEM imaging support SWATH-MS data, showing improved vector purity with serotype-specific resins AVB and PAAV9 and similar levels of impurity retention for PAAV8 and AAVX (Figs. 4 and 5B). In addition to differences in relative HCP content, there were notable differences in diversity of HCP species detected from purification with the different affinity resins (Fig. 3B). AVB and PAAV9 resins show a reduction in total and universal HCP identifications across the samples compared with AAVX resin. PAAV8 shows a similar number of unique and universal HCP species compared with AAVX resin. Lysate load material was used as a reference to identify resin-specific residual HCPs that appeared in all purification conditions for a given resin but were absent after purification with other resins. There were no universal unique HCPs identified for AVB resin. AAVX, PAAV8, and PAAV9 resins showed 18, 131, and 15 universal unique HCP hits, respectively (Supplementary Table S5).
sdAbs nonspecific binding drives HCP retention
Comparisons between EGFP lysate cycling for fresh resins and resins with sdAbs digested by Proteinase K indicate that differences in resin performance are driven by divergent levels of nonspecific binding of sdAbs to background impurities. AAVX and PAAV8 show notable binding and elution of background impurities, indicated by prominent UV 280 nm absorbance peaks and elevated elution pool protein concentration for cycling of EGFP lysate material that contains no AAV (Fig. 6 and Supplementary Fig. S5). AVB and PAAV9 resins show much lower levels of binding and elution of HCPs based on chromatograms and elution pool protein concentrations (Fig. 6). Nonspecific binding of background impurities is substantially lower for AAVX and PAAV8 resins after digestion of sdAb fragments by Proteinase K (Fig. 6).
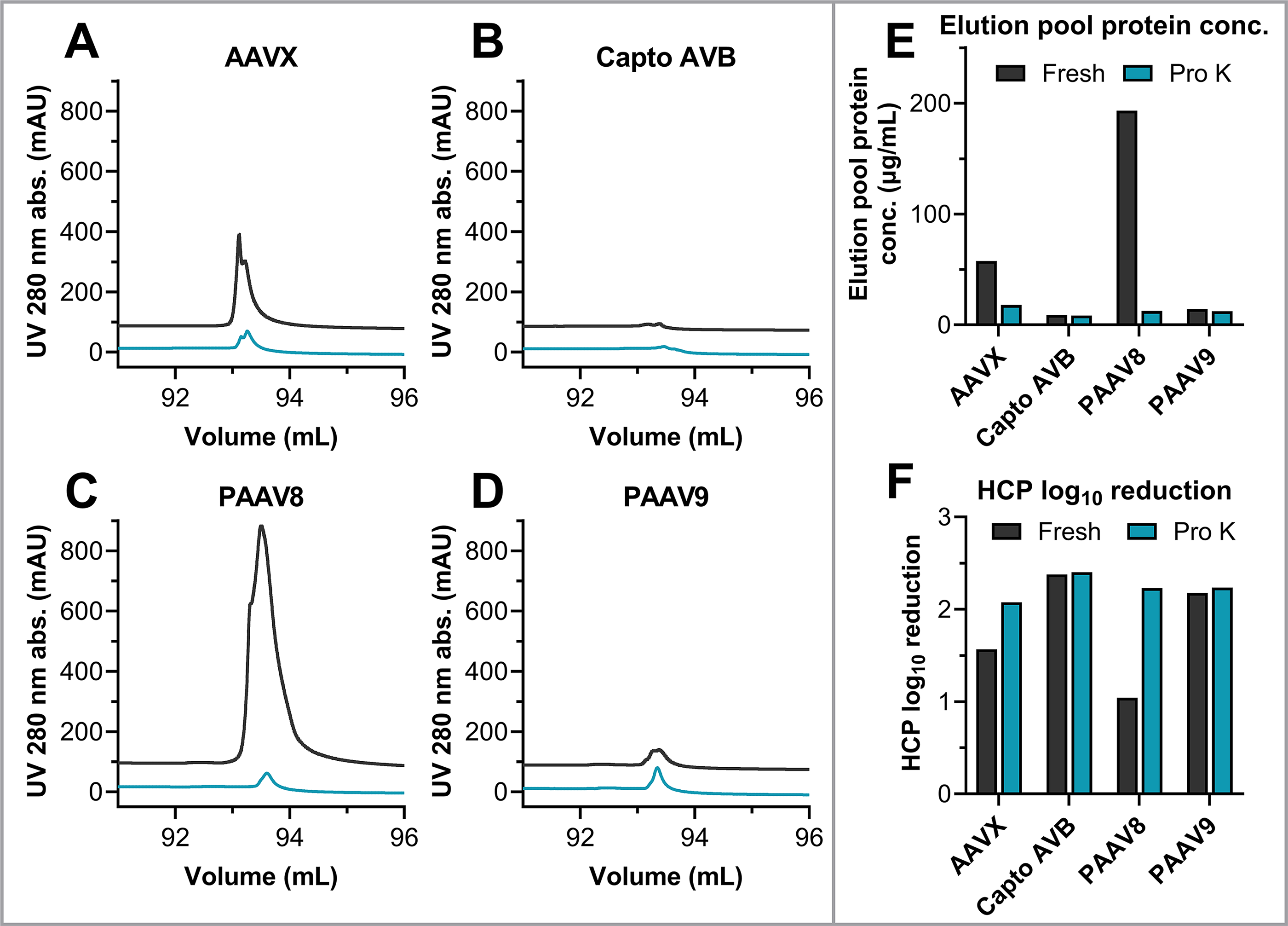
Zoomed in elution peaks for affinity chromatography performed with EGFP lysate load material using
DISCUSSION
Scalable purification platforms are needed to process AAV vectors from next-generation cell culture systems. 8 Multistage chromatography strategies have been demonstrated, which consist of AC coupled with anion exchange chromatography for the removal of empty capsids. 8 These purification systems show high capsid recovery and full capsid enrichment combined with flexibility for scale-up to support clinical manufacture and adaptability to high-throughput workflows for analytical characterization. Studies have been reported examining process-related impurity clearance for AAV AC, but these evaluations have not characterized how downstream process parameters influence HCP retention.20–22 In this work, we show that both harvest strategy and affinity resin selection strongly influence intermediate vector purity.
Affinity resins show varying levels of nonspecific impurity binding
The commercial marketing of different AAV affinity resins allows for serotype-specific resin selection to support small-scale vector preparation or for large-scale purification process development.12,23 Through in-depth and controlled analysis of residual HCP retention, we observed significant differences in purification quality based on AC resin selection. AAVX offers the advantage of broad binding tropism with affinity for all naturally occurring and most synthetic AAV serotypes. 12 However, purification with AAVX resin gives a higher amount (HCP mass/total protein mass) and a greater number of unique residual HCPs compared with AVB and PAAV9 resins. This divergence in resin performance was measured by elution pool protein concentration and SWATH-MS and is visually supported by SDS-PAGE and nsTEM imaging. Digestion of sdAb fragments of immunoaffinity resins decouples impurity interactions with the resin matrix and the ligand to show that observed differences in HCP retention are specifically driven by off-target sbAb binding. Interestingly, PAAV8 resins show similar levels of nonspecific binding to HCPs compared with AAVX, which highlights the need for new and improved AC resins for AAV purification. There is a significant process performance advantage for choosing AVB resins for AAV2 or AAV5 purification and PAAV9 resin for AAV9 purification instead of AAVX with no observed impacts to process yield. Parameters such as cell culture viability, harvest procedures, and capsid:HCP content likely influence overall results but are not directly evaluated here.
Supernatant-only harvest can substantially improve intermediate vector purity
Methods for AAV cell culture harvest must be evaluated on a product- and process-specific basis. Cell lysis is required for systems with low levels of product in the supernatant, which has been widely shown for AAV2 due to heparin affinity. 24 Other AAV serotypes are present in culture media to a varying degree. AAV8 has been shown to be present in large quantities in culture media, which is supported by results obtained in this work. 24 Piras et al. demonstrated that AAV8 distribution in the culture medium increases throughout culture duration, with the proportion of viral particles in the media reaching as high as 94% by day 7. 25 Additionally, these researchers noted that the full-to-empty capsid ratio of AAV8 was similar in the lysate and the culture media, further allowing for simplification of harvest by only processing the culture media while discarding cells. Results obtained here suggest that there is an advantage in intermediate vector purity for shifting to a media-only harvest process. AAV8 HCP content relative to capsid content decreased approximately 2-fold after affinity purification for supernatant harvested material compared with lysate harvested material. This reduction in impurity content can allow for enhanced quality of final vector products for clinical applications and improved vector purities for small-scale preparations in research settings.
Unique profiles of HCPs associated with lysate and supernatant processed material are likely driven by varying levels of protein secretion from the host cell during vector production; however, reduced culture viabilities can contribute to the release of additional cellular proteins. Proteins secreted into the culture medium at higher proportions may interact with viral particles in the media or carry through the downstream process due to nonspecific interactions with affinity resins. Phospholipase B-like 2 (PLBL2) was one of 347 HCPs uniquely detected in all lysate-purified vector samples but absent in supernatant-processed material. PLBL2 has been previously shown to copurify with monoclonal antibodies and pose significant immunogenicity risks to patients. 15 LC-MS/MS results indicate that retention of this “high-risk” HCP in intermediate vector preparations can be avoided by shifting to a supernatant-only harvest process.
CONCLUSIONS
We show here through controlled vector production and purification combined with SWATH-MS proteomic profiling that affinity resin selection and harvest process design have crucial impacts on AAV HCP content across multiple serotypes. sdAbs for AVB and PAAV9 give reduced nonspecific interactions with HCPs compared with AAVX and PAAV8 resins. Processing advantages for media-only harvest of AAV8 have been previously discussed, and we show the additional benefit of HCP reduction for processing AAV8 directly from the culture media for both EGFP packaged vectors and empty capsids. The results obtained in this work contribute to the improved understanding of AAV downstream parameter impacts to vector CQAs, facilitating the design of scalable next-generation purification platforms to deliver safe and efficacious therapies to patients.
Statistical analysis
Statistical comparisons between harvest lots (lysate vs. supernatant), vector transgenes (EGFP vs. empty), and affinity resins (AAVX vs. serotype-specific) were performed using nonparametric paired analyses (Wilcoxon test), with p < 0.05 considered statistically significant.
DATA AVAILABILITY
All data generated, discussed, or analyzed as part of this work are available upon reasonable request.
AUTHORS’ CONTRIBUTIONS
T.M.L. conceptualized the research, designed and performed the experiments, analyzed the data, created the figures, and wrote the original draft. L.M. generated LC-MS/MS data and edited the draft. K.H.L. edited the draft, secured funding, and provided supervision.
Footnotes
ACKNOWLEDGMENTS
The authors thank Shannon Modla in the Delaware Biotechnology Institute for supporting nsTEM imaging.
AUTHOR DISCLOSURE STATEMENT
The authors declare no competing interests.
FUNDING INFORMATION
This work was supported in part by financial assistance awards 70NANB17H002, 70NANB20H037, and 70NANB21H085 from the US Department of Commerce, and the
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.