
Abstract
Turkey coronavirus (TCoV) is a member of the Avian coronavirus species with infectious bronchitis virus (IBV), which is considered to be the source of TCoV. These 2 viruses are highly similar in all regions of their genomes, except for the spike gene, which is necessary for virus attachment. Although TCoV causes severe enteric disease in turkey poults, it does not cause clinical disease in chickens. However, considering that TCoV can infect chickens, it is important to distinguish TCoV from IBV in chickens. This is particularly true for chickens that are housed near turkeys and thus might be infected with TCoV and serve as a silent source of TCoV for turkeys. We developed and validated a real-time PCR assay to detect the spike gene of TCoV and sequenced a portion of this gene to evaluate the molecular epidemiology of TCoV infections associated with a commercial turkey premises in the United States in 2020–2021. We identified natural infections of TCoV in chickens, and based on the molecular epidemiology of the viruses detected, these chickens may have served as a source of infection for the commercial turkey premises located nearby.
Turkey coronavirus (TCoV; Coronaviridae, Gammacoronavirus, Avian coronavirus) is an enveloped, single-stranded, positive-sense RNA virus. Other members of the Avian coronavirus species are infectious bronchitis virus (IBV) in chickens, and guinea fowl coronavirus. Based on phylogenetic analysis, TCoV is proposed to have emerged from IBV through a recombination event, resulting in the incorporation of a spike (S) glycoprotein gene of unknown origin into IBV, permitting infection and disease in turkey poults. 7 TCoV and IBV are highly similar in all regions of their genomes, except for the S genes. There are high percent similarities for the S glycoproteins among TCoV isolates within the United States, which are >90%, whereas between TCoV and IBV the similarities are <36–50%.7,8 Epidemiologic evaluation of S genes from European variants of TCoV compared to those from the United States shows that the European viruses are distinct from U.S. variants, suggesting separate evolutionary pathways on the 2 continents. 3
The S glycoprotein is necessary for viral attachment to host cells. The change in the S protein in IBV led to the host shift to turkeys. Along with the host shift, there was a shift in viral tropism from respiratory epithelium for IBV in chickens to intestinal epithelium and bursa of Fabricius for TCoV in turkeys. 5 The turkey is thought to be the only natural host for TCoV, 5 but TCoV has been shown experimentally to infect chickens,6,7 and TCoV can be propagated in embryonated chicken eggs.5,6 TCoV tropism is considered the same in chickens as in turkeys, 5 but antigens and RNA of TCoV have also been detected in chicken respiratory epithelium after experimental infection. 4 Differences in viral tropism of TCoV, depending on the infecting strain in chickens, is likely the result of the strain variations resulting from recombination events that occur for TCoV.2,7 Experimentally, TCoV, despite multiple passages in chickens, failed to produce clinical disease in chickens, but the TCoV isolated from the GI tracts of the chickens did infect and produce lesions in turkey poults. 7
TCoV was first identified in the United States in 1973 and causes poult enteritis complex and poult enteritis mortality syndrome.5,9 TCoV contributes to significant economic losses for the turkey industry because of impaired growth and poor feed conversion. 5 TCoV remains a threat to turkey producers given the difficulty of eliminating the virus from high-concentration premises, lack of effective treatment, and absence of vaccine(s) to prevent disease. 8
TCoV detection has been performed in our laboratory by reverse-transcription real-time PCR (RT-rtPCR) using primers/probes for both the membrane protein (M) gene 10 and the nucleocapsid (N) gene. 9 There is >90% sequence identity for these 2 genes between TCoV and IBV, 5 and these assays were designed to detect both viruses. The N gene assay detected TCoV consistently; however, the M gene assay failed to produce a product for most positive TCoV samples tested in the Animal Disease Diagnostic Laboratory (ADDL) at Purdue University (West Lafayette, IN, USA) from 2018–2021. As stated, the N gene primer/probe set also detects IBV; therefore, when asked to evaluate chickens for TCoV, any positive samples were confirmed as TCoV by amplifying a portion of the S gene (primers S306F and S306R) 8 and sequencing. Hence, we designed primers and a probe specific for the TCoV S gene to be used in conjunction with the primers/probe for the N gene. 9 Both genes were targeted to avoid missing a positive sample resulting from S gene variability between TCoVs. This assay, along with molecular epidemiology of detected viruses, proved that chickens can be naturally infected with TCoV, and the evidence implies, as suggested previously, 6 that chickens may serve as a source of infection for turkeys.
Materials and methods
Samples and nucleic acid extraction
In July 2021, 8 commercial turkey flocks tested positive for TCoV. One of these flocks had a history of an outbreak of TCoV in September 2020, and the company depopulated the affected farm and later restocked. At the time of the second outbreak, the company began looking for a source of the virus for the outbreaks. Samples were submitted for TCoV testing from 16 commercial chicken table egg farms, which included 15 layer farms and 1 pullet farm that were in close proximity to the positive turkey flock. The samples included 16 individual cecal tonsils and 16 pools of 5 ceca each (1 tonsil and 1 pool from each farm). Samples were also submitted from neighboring turkey farms. All of the farms (chicken and turkey) were located within a 3.2-km (2-mi) radius.
Other samples used for our study included 82 known TCoV-positive samples (7 individual turkey intestines, 5 pools of 5 turkey intestines, 70 environmental drag swabs) and 31 known TCoV-negative samples (1 turkey intestine, 30 environmental drag swabs), based on testing using N 9 and M 10 targets and TCoV S gene sequencing. 8 These samples had been submitted for TCoV testing from U.S. commercial turkey farms from 2018–2022. Samples from 2022 from one farm that had a drop out of the N gene target with a published RT-rtPCR assay 9 were also included. Additionally, 24 IBV RT-rtPCR–positive samples 1 (oropharyngeal and tracheal swabs), submitted for IBV testing from backyard chicken flocks, were included.
Nucleic acids were extracted (KingFisher flex purification system, MagMAX CORE nucleic acid purification kit-complex workflow; Thermo Fisher). The internal positive control RNA (VetMAX Xeno; Thermo Fisher) was used according to the manufacturer’s instructions. Nucleic acids were stored at −70°C prior to testing.
Primers and probe design
TCoV S gene sequences from samples collected in the United States were obtained from GenBank (https://www.ncbi.nlm.nih.gov/genbank/). All of the sequences were aligned with MAFFT using the default parameters in Geneious Prime v. 2021.1.1 (Biomatters), and conserved areas were chosen for primer/probe development, which was performed with Primer3 (https://bioinfo.ut.ee/primer3-0.4.0/). Primer and probe sequences used in our study include those previously published (for N gene detection 9 and S gene sequencing 8 ) and those specifically developed for our work (Table 1).
Primers and probes used in our study of turkey coronaviruses (TCoVs) from turkey and chicken farms.

Nucleotide location based on TCoV/IN-517/94 (GenBank GQ427175).
Primers and probe were developed for a RT-rtPCR assay to detect the S gene of TCoV. Additionally, primers were developed to sequence the portion of the TCoV S gene containing the real-time primer/probe binding regions in contemporary variants from 9 TCoV-positive clinical samples that had been submitted for testing during 2021–2022 to the ADDL from 3 different companies among 3 states. Primers were also developed to sequence the portion of the N gene containing the primer/probe binding regions for the RT-rtPCR assay used in our laboratory to sequence a set of samples from 2022 from one farm that had a drop out of the N gene target with the RT-rtPCR assay. 9
Sequencing of RT-rtPCR primer/probe binding regions
RT-rtPCR reactions were performed (One step RT-PCR kit; Qiagen) in 50-µL total volume reactions containing 75 μmol of each primer. PCR parameters included 30 min at 50°C, 15 min at 95°C, and 40 cycles of 1 min at 94°C, 1 min at 60°C (S gene) or 65°C (N gene), and 1 min at 72°C, followed by 10 min at 72°C. PCR products were purified (ExoSAP-IT PCR product cleanup reagent; Thermo Fisher). Sanger sequencing was performed (Eurofins Genomics, Louisville, KY). The partial S gene sequences were submitted to GenBank (ON868855– ON868863). Sequenced regions were aligned (Clustal Omega, using default parameters in Geneious Prime) and compared with the primer and probe sequences for the S gene and N gene RT-rtPCR assays.
Multiplex RT-rtPCR assay
A multiplex RT-rtPCR assay targeting the S and N genes of TCoV was developed. Following optimization of the RT-rtPCR assay, each 25-µL RT-rtPCR reaction contained 6.25 µL of master mix (TaqMan fast virus 1-step; Thermo Fisher), 0.25 µL of DNA polymerase (AmpliTaq 360; Thermo Fisher), 10 μmol of each primer, and 5 μmol of each probe (IDT; Table 1, S and N RT-rtPCR), 1 µL of internal positive control (VetMAX Xeno LIZ assay; Thermo Fisher), and 8.5 µL of nuclease-free water. Seven µL of template nucleic acid was added to each reaction. A positive amplification control (nucleic acid from a known positive sample), negative extraction control (Xeno), and a negative template control (nuclease-free water) were used with each RT-rtPCR run. RT-rtPCR reactions were performed (ABI 7500 fast thermocycler; Thermo Fisher), and reaction conditions were 50°C for 10 min, 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 60 s.
Method validation
Limit of detection was evaluated using a 10-fold serial dilution series of a laboratory-validated positive control (confirmed by sequencing a portion of the S gene) 8 and by comparing the Ct values obtained for the S gene with Ct values obtained previously with the N gene primers/probe. 9 A standard curve generated with the dilution series was used to determine linearity and efficiency of the S gene primers/probe. Analytical specificity was evaluated in silico using BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) of the primer/probe sequences and compared specifically to IBV (taxid 11120) and to guinea fowl coronavirus (taxid 2341010).
Repeatability was established through repetitive testing of positive samples, 5 replicates in a single run (intra-assay repeatability), over 6 runs (inter-assay repeatability) with sample Ct values over the linear range of testing (Ct values at 19, 23, 26, 28, 31, 34). CVs were calculated for each set of results.
Positive predictive value (PPV) and negative predictive value (NPV) were evaluated with 82 known TCoV-positive samples and 31 known TCoV-negative samples We also tested 24 IBV-positive samples with the TCoV assay.
A portion of the S gene was sequenced from positive samples and from samples from the outbreaks in 2020 and 2021 using primers described previously 8 (Table 1). Sanger sequencing was performed (Eurofins Genomics). Sequences were deposited in GenBank (ON868842–ON868853). The sequences were aligned (MAFFT; Geneious Prime) to sequences obtained from other commercial turkey flocks, and a phylogenetic tree was prepared in Geneious tree builder, HKY genetic distance model, neighbor-joining method with no outgroup, and bootstrapping with 1,000 replicates. Clades with bootstrap values ≥70% were considered significant.
Results
Primers and probe design and binding region sequencing
The primer/probe binding regions for the S gene RT-rtPCR sequenced in the contemporary variants showed some single nucleotide changes in these regions compared to older variants found in GenBank (Fig. 1). Sequences obtained for the primer/probe binding regions of the N gene from the sample with N gene drop out in the assay had a single nucleotide change in the probe binding region and 2 nucleotide changes in the reverse primer binding region (data not shown).

Primer/probe binding regions for our turkey coronavirus (TCoV) S gene RT-rtPCR assay, with comparison between design and sequences of contemporary strains. Compared to contemporary variants (5–13), primers RT-rtPCR S gene F (14) and R (4) and the probe (1) each only have 1 potential mismatch with the corresponding binding region. To date, none of these changes have been shown to have an effect on our assay. Older TCoV variants are represented by GenBank GQ427175 (2) and KF652237 (3).
Method validation
Both S and N gene targets could be detected to a dilution of 106, representing Ct values between replicates of 36–37, suggesting 37 as a cutoff Ct value for a positive sample. Based on BLAST results, the designed primers for the S gene were specific for TCoV and bound to conserved areas of the gene. CVs for intra-assay and inter-assay testing were 0.2–0.7% for the designed S gene primers/probe. The slope of the line generated from the standard curve for the S gene target was −3.46, the efficiency of the reaction was 95%, and the R2 was 99%.
All positive and negative results were reproduced with the new assay (100% PPV, 100% NPV). Both the N and S gene targets were positive for all 82 positive samples, and the Ct values were 9.8–36.5 (x– Ct value 23.9). All IBV-positive samples were positive for the N gene target but were negative for the S gene target.
Of the 16 cecal tonsils tested for TCoV from chicken layers, 5 were positive for TCoV. The Ct values were 18.5–28.9 for the S gene and 16.4–23.2 for the N gene. A portion of the S gene was sequenced from 4 of these samples (layers 1–4_Farm 1; Fig. 2). For the intestinal pools from chicken pullets, 5 of 16 pools were positive for TCoV (Ct values for S gene 28.9–33.9). One of these samples was sequenced (Chicken pullets_Farm 2; Fig. 2). None of these chickens had signs consistent with TCoV.
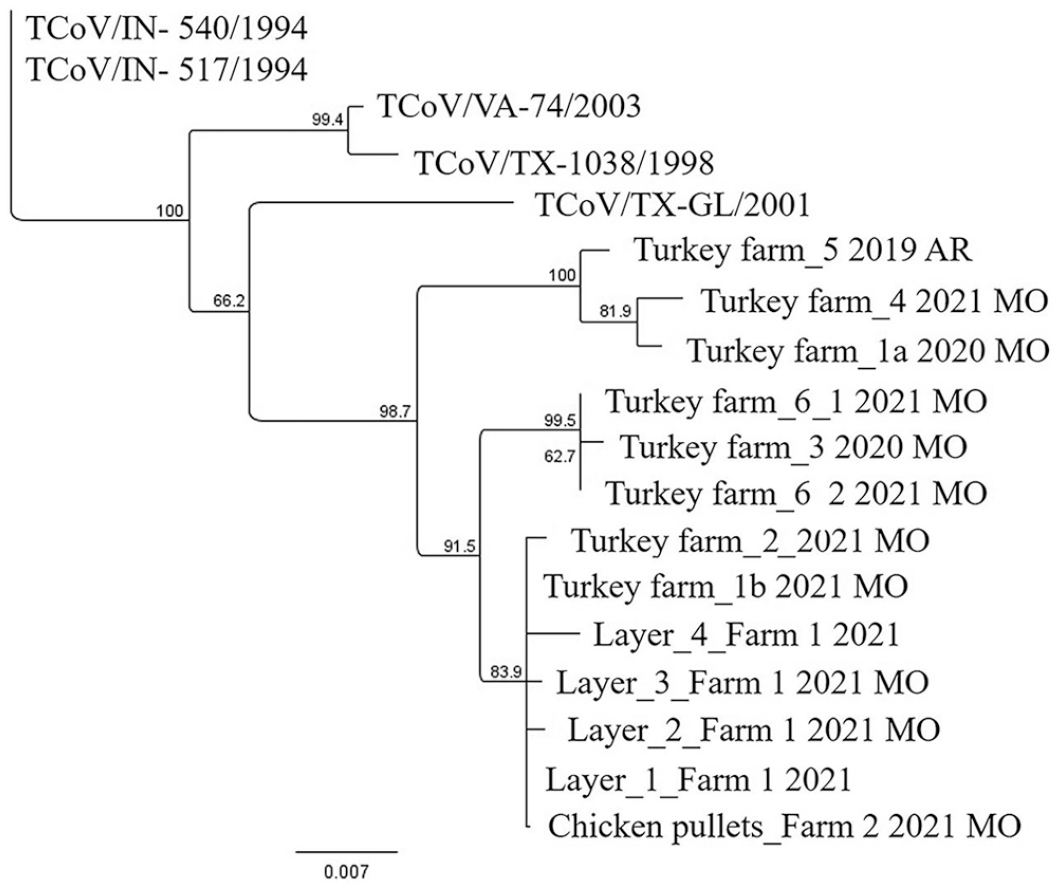
Neighbor-joining tree, with 1,000 bootstrap replicates, generated from portions of the turkey coronavirus (TCoV) S gene obtained from samples from commercial turkey and chicken flocks. Tree also includes older sequences from GenBank: TCoV/IN-540/1994 (EU022525), TCoV/IN-517/1994 (GQ427175), TCoV/VA-74/2003 (GQ427173), TCoV/TX-1038/1998 (GQ427176), and TCoV/TX-GL/2001 (GQ427174). Clades with bootstrap values ≥70% were considered significant.
Based on the phylogenetic analysis (Fig. 2), the outbreaks on the turkey farm in 2020 (Turkey farm_1a) and 2021 (Turkey farm_1b) were caused by 2 different TCoV variants. The virus from the first outbreak grouped with viruses detected from another turkey farm with the same company (Turkey farm_4; Fig. 2) and from a turkey farm associated with a different company in a different state (Turkey farm_5; Fig. 2). However, there is some variation within this group of viruses, which, based on our data, is related to geographic location. The second outbreak was associated with a virus that grouped with the TCoV viruses from the chicken farms but also with a second turkey farm with the same company (Turkey farm_2; Fig. 2) that is located in the same geographic location as turkey farm 1. This clade containing the TCoV viruses from the chickens is not fully resolved, but there are some genetic differences between the “chicken TCoV variants” between neighboring farms within the same company. Turkey farms 3 and 6 are with 2 different companies, but the viruses from these farms have similar sequences. The currently circulating variants differ from previously sequenced isolates collected in the mid-to-late 1990s and early 2000s.
Discussion
Our assay developed for detection of the TCoV S gene used in conjunction with primers for the N gene was highly sensitive and specific. Our assay is based on use of a variable gene, and although necessary, it creates an increased need to monitor for changes in the primer/probe binding regions that will occur over time for this RNA virus. Based on sequences from these binding regions in contemporary variants versus older variants in GenBank, there have been a few changes in the binding regions, but to date, these changes have occurred in areas shown not to have an effect on our RT-rtPCR assay.
Inclusion of the N gene (highly conserved in TCoV and IBV) in the assay should prevent a false-negative result that would occur from changes in the S gene in primer/probe binding regions creating a drop out of S gene detection. Interestingly, one variant detected in 2022 had an N gene drop out with our RT-rtPCR assay. Although none of these changes were at or near the 3′-end of the primer, which is considered most likely to affect the assay, 2 of the changes were A to G and G to A, which are considered to have a severe impact on amplification. 11 In this particular case, detection of the S gene maintained the ability of the assay to detect the positive sample. Other than drop out of a target, comparison of the Ct values for each target may allow detection of noticeable shifts to the right for the S gene (or less likely the N gene) Ct value, suggesting changes to the binding regions and reduced primer/probe binding. Of note, our assay is specific for U.S. variants of TCoV. Variants detected in other countries have been shown to have an S gene with significant sequence variation from U.S. strains. 3 Our data also confirm that sequences of the S gene from contemporary variants differ from those obtained in the mid-to-late 1990s and early 2000s, as would be expected, based on replication errors known to occur with RNA viruses and the continued evolution of the viral attachment protein.
Our assay differentiated TCoV from IBV in samples from chickens that were naturally infected with TCoV. Sequencing confirmed that the same TCoV variants can circulate on both turkey and chicken commercial farms that are located within a close geographical location (within a 3.2-km [2-mi] radius). This suggests that infected chickens without clinical signs of disease may be a source of TCoV infection for turkeys. Variants found in the chickens from different farms also differed somewhat from each other, suggesting the potential for evolution of this virus within chickens, or multiple introductions with different viruses. Considering that these chicken farms are all part of the same commercial entity, it is more likely the virus has evolved while maintained within the population.
Similar TCoV variants can be found on turkey farms that are part of different commercial entities, and even across different states. It is unknown how these particular variants moved from one farm to another, but this suggests similar sources for these variants on multiple farms that then continued to evolve independently. Biosecurity for the turkey industry needs to consider not just transfer of TCoV from one turkey farm to another but potentially also transfer from chicken farms to turkey farms or vice versa.
Footnotes
Acknowledgements
We thank the staff of the chicken farms associated with our work for providing birds for sample collection and testing, the veterinarians for collecting and submitting the samples, and the technicians in the Animal Disease Diagnostic Laboratory (Purdue University, West Lafayette, IN, USA) for running the real-time PCR assays and assisting with assay validation.
Declaration of conflicting interests
Brian Wooming is a veterinarian employed by Cargill. The other authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.