
Abstract
Malassezia pachydermatis is part of the normal microbiota of canine skin and external ear canal, and is also associated with otitis externa in dogs. Laboratory detection of Malassezia otitis relies on the presence of elevated numbers of the yeast on cytologic examination of otic exudate. Although cytology has high specificity, it has low sensitivity, resulting in false-negatives and posing a challenge for clinicians to accurately diagnose Malassezia otitis. We developed a quantitative PCR (qPCR) to detect and quantify M. pachydermatis yeasts and validate the method with swabs from external ear canals of dogs. Our qPCR uses the β-tubulin gene, a single-copy gene, as a target. The limit of quantification was established as 0.18 ng/reaction, equivalent to 2.0 × 104 genome equivalents (gEq). Swabs from healthy dogs yielded quantification values of ≤2.7 × 104 gEq in the qPCR, whereas swabs from dogs with otitis yielded quantification values of ≥2.5 × 105 gEq. Our qPCR assay provides accurate quantification of M. pachydermatis yeasts from swab samples from dogs, is more sensitive than cytology, and could be used to monitor response to treatment. Our assay could also be valuable in a research setting to better understand the pathogenesis of M. pachydermatis.
Introduction
The lipophilic yeast Malassezia pachydermatis is a normal inhabitant of healthy canine skin and mucosae. 17 The skin population of M. pachydermatis on dogs can overgrow and act as an opportunistic pathogen, causing dermatitis and otitis externa in animals affected with atopic dermatitis, intertrigo, endocrinopathies, and primary keratinization defects. These concurrent diseases can occur without necessarily favoring the yeast infection. 6 Factors that favor proliferation of M. pachydermatis and the transition from a commensal to a pathogen on canine skin have not been fully clarified, although they seem related to skin disturbances by physical, chemical, and immunologic mechanisms. 36
Otitis externa is a common and multifactorial cutaneous disorder affecting up to 20% of dogs seen by veterinarians. 2 Otitis externa associated with M. pachydermatis is often characterized by the presence of a waxy, moist, and dark exudate, with erythema and pruritus, and lesions that are often colonized by an increased number of M. pachydermatis. 17 Diagnosis of otitis externa caused by M. pachydermatis is based on the observation of compatible lesions, the response to antifungal therapy, and the presence of elevated numbers of the yeast by direct microscopic observation. 6 Microbiologic culture is not routinely performed, and it is required only when direct microscopy is negative in animals with suspected infections. 11 However, quantitative culture has been suggested to have diagnostic value.4,5 Moreover, culture and susceptibility testing allows monitoring for the emergence of azole-resistant strains of M. pachydermatis. 20 A comparison of cytologic examination and fungal culture concluded that cytologic examination has good specificity but low sensitivity. 11 Consequently, there is a need for a specific, sensitive, precise, and rapid method to detect and quantify M. pachydermatis yeasts from dogs with otitis externa.
Several qPCR assays have been developed to detect and quantify the most frequently isolated Malassezia species from human skin, mainly M. globosa and M. restricta.1,15,32,33 Those assays usually amplified rRNA genes, which have been shown to be present in multiple copies in the genomes of ascomycete and basidiomycete fungi,3,28,31 and were designed to study the distribution of Malassezia species on human skin rather than the absolute quantification of these yeasts.
We developed a fast, sensitive, and accurate technique to reliably detect and quantify M. pachydermatis yeasts from the external ear canal of dogs. We selected the β-tubulin gene as an alternative qPCR target to the rRNA genes because it has been shown to be present only in a single copy in closely related species including M. globosa, 38 and it has also been proposed as a phylogenetic marker for the differentiation of Malassezia species. 12
Materials and methods
Strains and samples
We tested 15 M. pachydermatis strains representative of the 8 β-tubulin genotypes 27 of this species with the developed assay (Table 1), in order to ensure amplification of all genotypes of the target gene. Also, 16 strains, each representing a different Malassezia species, were tested to assess the specificity of the qPCR.
Malassezia spp. strains used, including the host species and location from which they were retrieved, β-tubulin sequence, and melting temperature (Tm) of the PCR product.
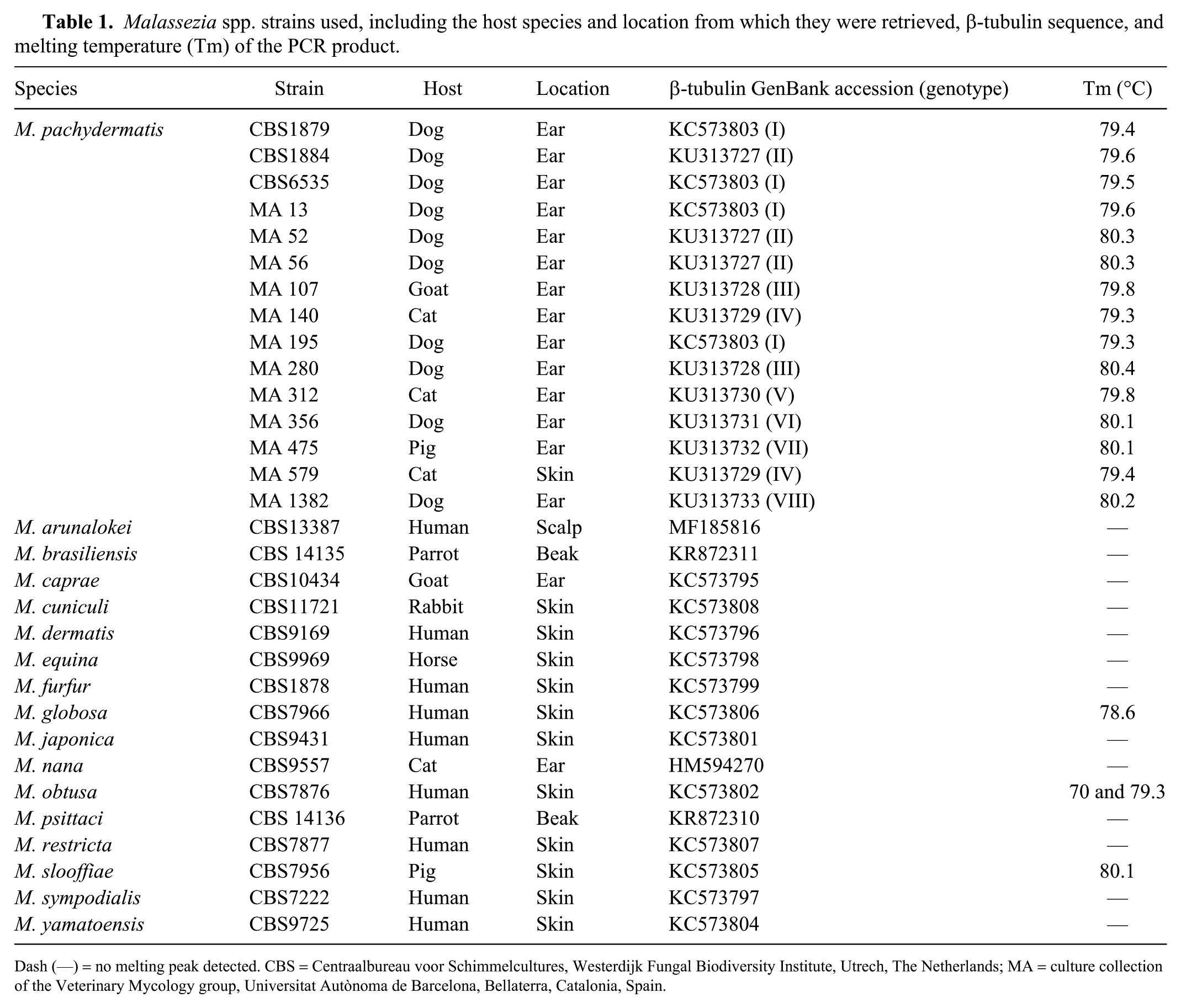
Dash (—) = no melting peak detected. CBS = Centraalbureau voor Schimmelcultures, Westerdijk Fungal Biodiversity Institute, Utrech, The Netherlands; MA = culture collection of the Veterinary Mycology group, Universitat Autònoma de Barcelona, Bellaterra, Catalonia, Spain.
To validate the assay, 24 swabs from the external ear canal of 17 dogs were analyzed. When possible, both ears were sampled regardless of the health status of the ear. Eleven swabs were collected from the external ear canals of 7 healthy dogs with no visible signs of otitis externa. Eleven swabs were obtained from the external ear canal of 9 dogs in which otitis caused by M. pachydermatis was diagnosed by veterinary clinicians based on otoscopic examination (presence of inflammation, a waxy brown exudate, pruritus) and elevated numbers of yeast by direct microscopic observation. Also, 2 swabs from 1 dog with otitis externa caused by M. pachydermatis that had been treated topically for 15 d with miconazole, polymyxin B, and prednisolone (Surolan; Elanco España, Madrid, Spain) were processed. Samples from healthy dogs without otitis were obtained from the dog pound of the Autonomous University of Barcelona (UAB; Spain) following procedures approved by Ethics Committee on Animal and Human Experimentation from UAB and Generalitat de Catalunya (approval CEEAH 1240; date of approval 28 February 2012). Samples from cases of Malassezia otitis were obtained during routine veterinary procedures and with verbal owner consent with the collaboration of veterinary clinicians from the Veterinary Teaching Hospital of the UAB.
Microbiology
In all cases, 2 swabs were obtained from the affected ear of the same dog. One swab was maintained at −20°C and used for qPCR analysis. The second swab was used for classical microbiologic processing and cytologic examination. Briefly, one side of the swab was streaked on a Sabouraud glucose agar (SGA) plate (Oxoid, Madrid, Spain), with 0.05 g of chloramphenicol, and on a 5% sheep blood agar plate (BD Difco, Madrid, Spain). Plates were incubated at 37°C with 5% carbon dioxide for 5 d, and colony-forming units (CFU) were counted. M. pachydermatis was identified by microscopic morphology and by the ability to grow on SGA. 16 The other side of each swab was rolled on a clean glass slide and stained (Diff-Quick stain; Microptic, Barcelona, Spain). The stained slides were examined microscopically (1,000× magnification). Yeast cells with morphology compatible with M. pachydermatis were counted in 10 random fields, and the presence of other fungi and bacteria were also noted.
DNA extraction
M. pachydermatis strains were grown on SGA; strains of the other Malassezia species were grown on modified Dixon agar. 16 After 5 d of incubation at 32°C, DNA was extracted from yeast colonies (FastDNA spin kit protocol, FastPrep FP-24 instrument; MP Biomedicals, Biolink, Barcelona, Spain). DNA was extracted from swabs (UCP Pathogen kit; Qiagen, Madrid, Spain) according to the protocol recommended for swab samples and using pre-treatment with mechanical disruption. A sterile swab was processed under the same conditions as the external ear canal samples in order to control cross-contamination. DNA was stored at −20°C until used as template in the qPCR.
Primer design
Primers were designed to amplify a fragment of the β-tubulin gene of M. pachydermatis. Sequences used for primer design were obtained in a previous study in which 8 genotypes of this gene were described among M. pachydermatis strains. 27 Primers MPFRT-2 (5’-CGGACGAGACGTTCTGCATT-3’) and MPRRT-2 (5’-TTGAGTGTGCGGAAGCAGAT-3’) were designed (Primer Express v.3.0; Applied Biosystems, Foster City, CA). Primer dimers and secondary structure formation were evaluated (Primer Express v.3.0; IDT UNAFold online tool, https://eu.idtdna.com/calc/analyzer). Both primers were checked against the NCBI database using Primer-BLAST to confirm specificity.
qPCR development
We developed our qPCR following the defined criteria of the Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) guidelines. 8 The assays were performed (Applied Biosystems 7500 real-time system; Thermo Fisher Scientific, Waltham, MA) using SYBR Green chemistry with the absolute quantification method.
Optimal concentrations of the forward and reverse primers were determined by testing concentrations of 100–800 nM. Concentrations giving the lowest quantification cycle (Cq) values for the standards were selected: 300 nM for the forward primer and 500 nM for the reverse primer. The optimized reaction mix (20 µL final volume) contained 10 µL of 2× SYBR Green PowerUp PCR master mix (Applied Biosystems, Foster City, CA), 0.45 µL of 13.3 μM forward primer, 0.75 µL of 13.3 μM reverse primer, 3.8 µL of deionized DNase- and RNase-free water, and 5 µL of template DNA. Runs were performed using the following thermal cycling conditions: 1 cycle of 50°C for 2 min, 1 cycle of 95ºC for 2 min, 30 amplification cycles of 95°C for 15 s (denaturing step), and 60°C for 1 min (annealing-extension step and data collection). Next, a melting curve analysis was performed, with a gradual increase of temperature from 60°C to 95°C.
To achieve absolute quantification of the samples, the standard curve method was used. A standard curve of genomic DNA (gDNA) extracted from the neotype strain of M. pachydermatis CBS1879 was constructed, including 8 ten-fold dilutions, from 180 ng to 0.018 pg of DNA. The quantity and purity of the gDNA were determined by spectrometry (NanoDrop 2000; Thermo Scientific, Barcelona, Spain). The amount of gDNA was extrapolated to genome equivalents (gEq), considering that β-tubulin is a single-copy gene in the M. pachydermatis genome, the size of which is ~8.15 Mbp.35,37
Efficiency was assessed in every run, based on the correlation coefficient (r2) and slope values from the standard curve, and sensitivity was evaluated with the limit of quantification (LOQ; the lowest concentration of DNA amplified from the standard curve). The linear dynamic range was determined (highest to lowest gDNA dilution amplified from the standard curve). 8
To test for inhibition of the qPCR, gDNA standards were spiked with randomly selected samples from dogs. The Cq values of spiked standards were compared to the Cq values obtained from the same non-spiked standards to ensure that amplification was equivalent. Non-template controls (NTCs), in which DNA was substituted by water in the reaction mix, allowed verification that no contamination occurred and no primer dimers were formed. In each run, all samples were run in triplicate, including the standards and NTCs.
Statistical analysis
Statistical analyses were performed (Minitab 17 statistical software; Minitab, State College, PA). A multiple regression analysis was used to study the relationship between the qPCR quantification values and values obtained by cytologic examination and plate counts. The Ryan–Joiner normality test was applied to determine whether data followed a normal distribution. Differences in cytologic examination, CFU, and qPCR quantification values between healthy dogs and dogs with Malassezia otitis externa were tested by Kruskal–Wallis test. Samples with a qPCR value less than the detection limit were assigned to 2 × 104 gEq value for statistical analysis. Samples from dog 8 and dog 17 were excluded from the statistical analysis. The significance level for all statistical analyses was set at p ≤ 0.05.
Results
Primer design and specificity
The primers amplified a 61-bp amplicon with no identity to the dog genome. A BLAST search against the NCBI database showed no identity to other commensal and pathogen bacteria or fungi that can be found in the ear canals of dogs. In addition to the β-tubulin gene from M. pachydermatis, the next sequence matches belonged to organisms that are not related with domestic carnivores, such as a mushroom (Coprinopsis argentea), animals (Drosophila navojoa, Cyprinus carpio, Esox lucius), and a plant (Nelumbo nucifera).
The primers consistently amplified the DNA from all tested M. pachydermatis strains. Melting curve analyses yielded a single peak in all M. pachydermatis strains at 79.3–80.4°C depending on the genotype (Table 1). No detectable amplification of other Malassezia species was observed, except for M. globosa, M. slooffiae, and M. obtusa. The M. globosa strain showed a single peak with a clearly different melting temperature (Tm) of 78.6°C; the M. obtusa strain gave a melting curve with 2 peaks. For the M. slooffiae strain, a single dissociation peak with a Tm of 80.1°C was obtained.
qPCR performance
A fragment of the β-tubulin gene from M. pachydermatis was consistently amplified in a linear dynamic range of 4 ten-fold dilutions of gDNA, from 2.0 × 107 to 2.0 × 104 gEq. The standard curve yielded r2 values > 0.990 in all runs, and slope values of approximately −3.40 (Supplementary Fig. 1). All replicates were amplified with high precision, with <1 Cq of variation among them and standard deviation values <0.4. The LOQ was established as 0.18 ng/reaction, equivalent to 2.0 × 104 gEq, which corresponds to the lowest standard dilution amplified and accurately quantified (Cq ≥ 28).
Microbiology and validation of the qPCR using clinical samples
The presence and amount of M. pachydermatis yeasts of samples from the external ear canal of dogs were also assessed by plate counting and cytologic examination (Table 2). Samples from dogs without otitis yielded no growth of M. pachydermatis or plate counts of 0–8 CFU/plate (median: 3 CFU/plate); yeast cytology was negative in most of the samples (8 of 11; range: 0–2; median: 0 cells/field). In samples from dogs with Malassezia otitis, cytology revealed 1–5 M. pachydermatis cells/field (median: 2 cells/field); plate counts were 10–250 CFU/plate (median: 210 CFU/plate). In most of the samples (7 of 11), bacterial colonies also grew on blood agar plates, mostly gram-positive cocci.
Results obtained in samples from dogs with otitis externa by Malassezia pachydermatis and healthy dogs, including cytologic examination, plate counts, and qPCR quantification values, expressed as genome equivalents (gEq).
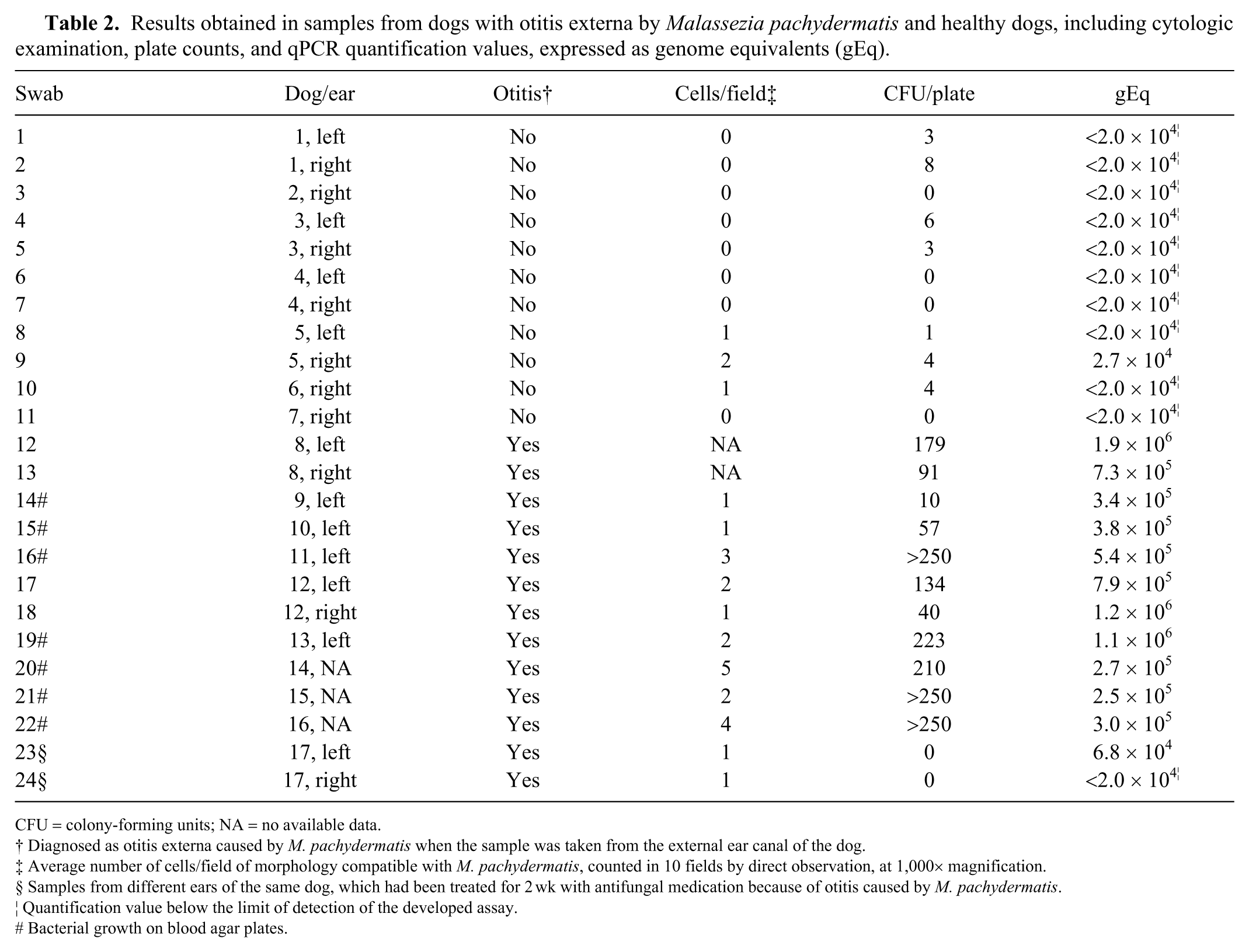
CFU = colony-forming units; NA = no available data.
Diagnosed as otitis externa caused by M. pachydermatis when the sample was taken from the external ear canal of the dog.
Average number of cells/field of morphology compatible with M. pachydermatis, counted in 10 fields by direct observation, at 1,000× magnification.
Samples from different ears of the same dog, which had been treated for 2 wk with antifungal medication because of otitis caused by M. pachydermatis.
Quantification value below the limit of detection of the developed assay.
Bacterial growth on blood agar plates.
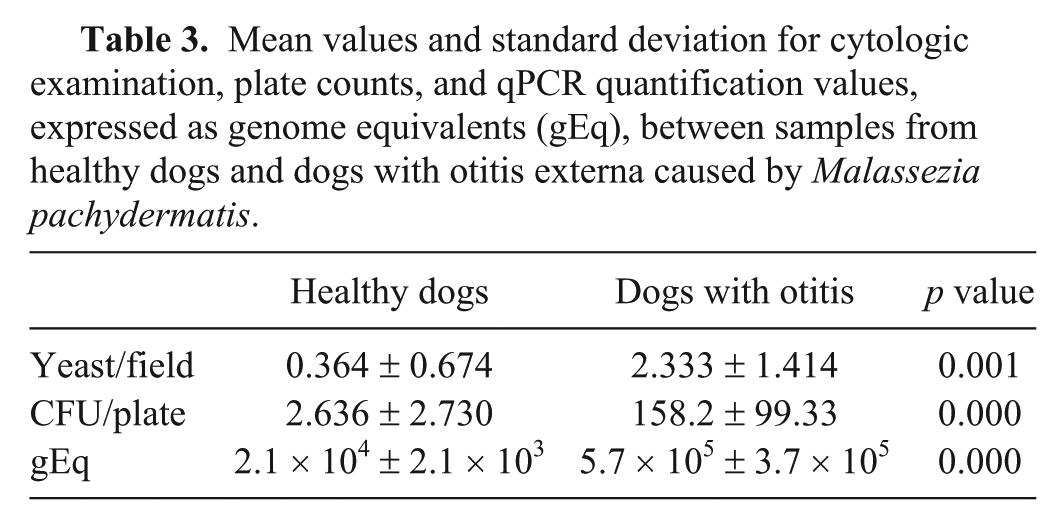
In the developed qPCR, quantification values of 2.0 to 2.7 × 104 gEq were obtained from samples from healthy dogs (median: 2.0 × 104 gEq); values from 2.5 × 105 to 1.9 × 106 gEq (median: 3.8 × 105 gEq) were obtained from samples from dogs with otitis caused by M. pachydermatis (Table 2). Multiple regression analysis showed that the relationship (R2 = 67.0%) between qPCR quantification and cytologic examination and plate counts in the model was statistically significant (p < 0.001). Statistically significant differences in cytologic examination, plate counts, and qPCR quantification values were found between samples from healthy dogs and dogs with otitis externa (Table 3).
Mean values and standard deviation for cytologic examination, plate counts, and qPCR quantification values, expressed as genome equivalents (gEq), between samples from healthy dogs and dogs with otitis externa caused by Malassezia pachydermatis.
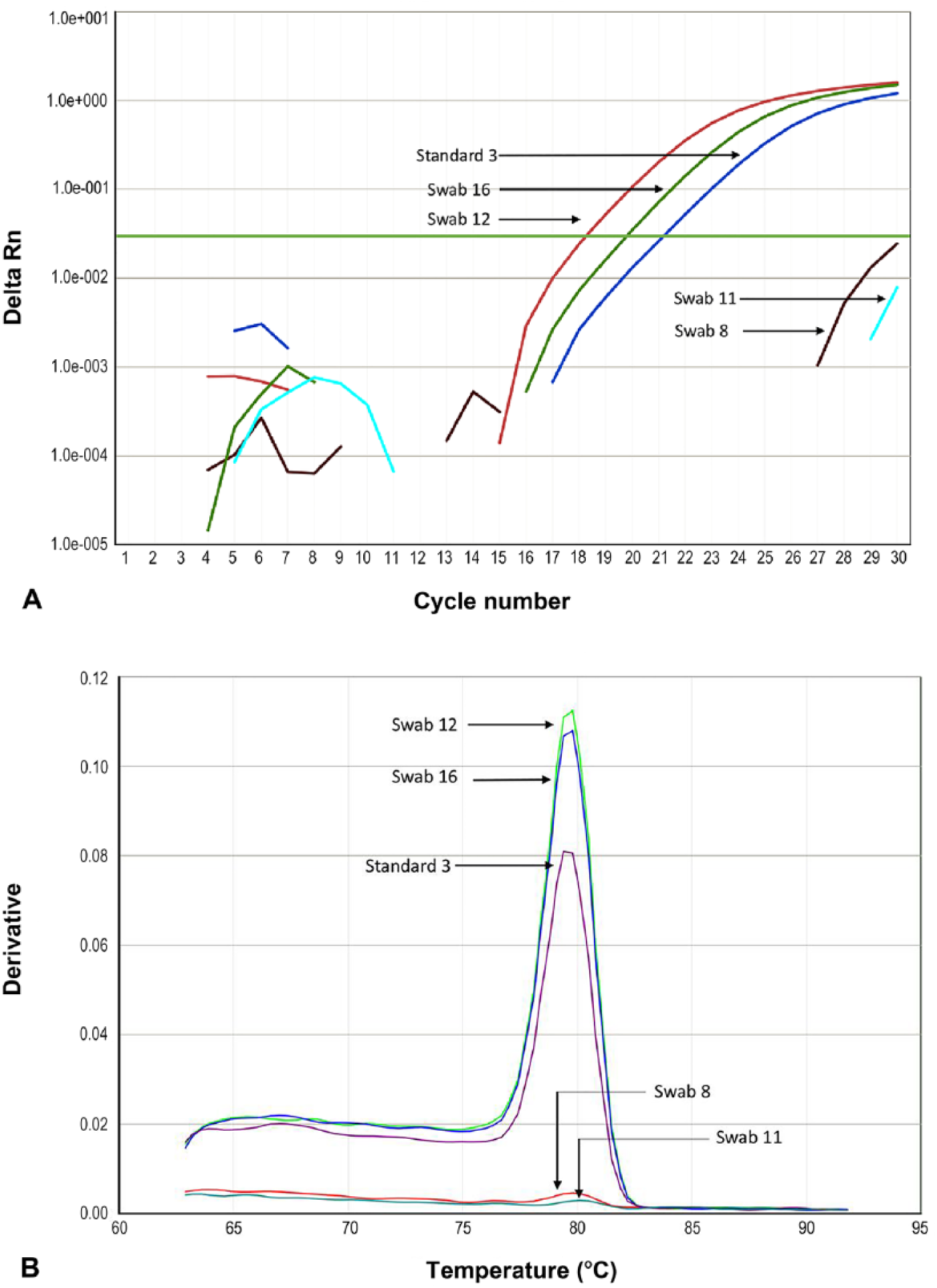
Melting curve analyses provided a single peak in all cases, with Tms of 79.4–80.4°C (Fig. 1). Amplification of NTCs was not detected, and inhibition was not detected in gDNA standards spiked with samples from dogs.
Discussion
The external ear canal of dogs can be colonized by a variety of bacterial and fungal species. In previous massively parallel sequencing studies, it was found that the predominant bacteria of the external ear canal of dogs are members of phyla Proteobacteria, Actinobacteria, and Firmicutes, 30 and some of the predominant fungi correspond to members of genera Alternaria, Cladosporium, Epicoccum, and Cryptococcus, as well as Malassezia spp. 21 A BLAST search showed that our primers did not match any of the deposited sequences of any of the organisms found in dog ear canals in 2 prior studies.21,31 Our designed primers showed amplification of all tested M. pachydermatis strains with consistent Tms within 1°C of variation. Although a slight range of variation can occur in melting curve analyses depending on factors such as SYBR Green or DNA concentrations, an interval variation of ±1°C among Tms is considered to be acceptable for specificity determination.7,22,29 Amplification of the 16 other Malassezia species was not detected, except for M. globosa, M. obtusa, and M. slooffiae. Although these 3 species provided Tms similar to M. pachydermatis strains, M. globosa and M. obtusa could be differentiated by melting curve analysis. Nonetheless, amplification of these species should not be considered as a disadvantage of our technique. M. globosa, M. obtusa, and M. slooffiae have never been isolated from dogs using culture-based methods9,13,34 or detected by molecular-based methods, 19 to our knowledge. There is only one report of M. obtusa associated with canine otitis externa. 14 However, this isolate was identified only by phenotypic characteristics, 14 and a subsequent molecular analysis showed the isolate to be M. nana. 10
Our qPCR assay has a LOQ of 0.18 ng of M. pachydermatis DNA, which corresponded to 2.0 × 104 gEq/reaction. Some qPCR methods have been developed for quantification of different Malassezia species from human skin. In these methods, LOQ ranged from 102 to 105 amplicon copies per reaction depending on the Malassezia species detected.26,33 All of these qPCR assays are based on multi-copy genes and, therefore, are more sensitive than single-copy genes, although quantification is less accurate. 18
Statistically significant quantitative differences were found by all of the methods used (cytologic examination, culture, and qPCR) in samples from dogs according to their health status (healthy or otitis caused by M. pachydermatis). Samples from healthy dogs yielded plate counts ≤8 CFU/plate, a maximum of 2 M. pachydermatis cells/field on cytologic examination, and qPCR quantification values of ≤2.7 × 104 gEq. Samples from dogs with otitis yielded plate counts ≥10 CFU/plate, 1–5 M. pachydermatis cells/field on cytologic examination, and qPCR quantification values of ≥2.5 × 105 gEq. No colonies were grown on plates from the swabs obtained from a dog that had been treated for M. pachydermatis otitis externa, and qPCR quantification values similar to the LOQ were obtained from these samples. M. pachydermatis is considered a secondary cause that contributes to otitis only in the abnormal ear or in combination with predisposing or perpetuating factors. 25
For some authors, the observation of >5–10 cells of morphology compatible with M. pachydermatis in several microscopic fields or >70 CFU per sample from ear specimens of dogs should be considered as an abnormal increase of Malassezia.2,11,17,23 However, these proposed guidelines are not generally accepted because a small population of the yeast might cause disease in sensitized animals, and variations in population sizes have been observed between different breeds. 6 Although it is not known whether there is a threshold population density needed for infection, the population of M. pachydermatis in dogs with skin disease can be increased 100–10,000 fold. 6 An accurate quantification of M. pachydermatis yeasts in swab samples from dogs can be achieved with our qPCR method.
In some cases in our study, results from cytology, culture, and qPCR did not correlate. This disagreement could be explained in part by the fact that cytology has low sensitivity. 11 Also, diverse factors can affect the number of M. pachydermatis colonies grown on a culture plate. As an example, M. pachydermatis has a tendency to form clumps, meaning colonies could have originated from a clump of cells instead of a single yeast cell. 24 Moreover, CFU reflect the number of live yeast, whereas the qPCR will give the total amount of the target gene detected, including both live and dead yeast.
Our qPCR assay provides an accurate quantification of M. pachydermatis yeasts in swabs from the external ear canal of dogs. The sample required for the assay can be obtained non-invasively, does not require any particular transportation condition, and can be stored at −20°C until DNA extraction. Moreover, it is possible to achieve accurate quantification of M. pachydermatis in a few hours. Identification of the primary cause of otitis externa is the most critical factor to address in treating otitis, particularly for chronic cases, but the elimination of the primary cause without an effective management of the concurrent infections typically fails to resolve clinical signs. 2
One of the limitations of our study was the small number of dogs sampled. Another limitation was the lack of data about the severity of otitis and dermatologic histories of the dogs. Further studies are needed, with those studies based on an increased number of dogs, details of clinical scores of otitis, and dermatologic history (to correlate the number of M. pachydermatis yeasts with diagnostic results). Such studies should be able to confirm that our qPCR could improve the diagnosis of otitis by M. pachydermatis in dogs, and consequently lead to more accurate treatment that would enhance the prognosis of the affected animals.
Supplemental Material
DS1_JVDI_10.1177_1040638719840686 – Supplemental material for Quantification of Malassezia pachydermatis by real-time PCR in swabs from the external ear canal of dogs
Supplemental material, DS1_JVDI_10.1177_1040638719840686 for Quantification of Malassezia pachydermatis by real-time PCR in swabs from the external ear canal of dogs by Laura Puig, Gemma Castellá and F. Javier Cabañes in Journal of Veterinary Diagnostic Investigation
Footnotes
Acknowledgements
We thank Mar Bardagí, Laura Ordeix, and Raquel Montoya for the samples kindly provided for our work. We also thank Carolina Gómez from the Veterinary Mycology Group of Universitat Autònoma de Barcelona (UAB) for valuable technical assistance. We thank the anonymous reviewers for their critically reading of our manuscript and their many insightful comments and suggestions.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Financial support for this project came from Servei Veterinari de Bacteriologia i Micologia from the Universitat Autònoma de Barcelona.
Supplementary material
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.