
Abstract
We examined the pathogens, morphologic patterns, and risk factors associated with bovine respiratory disease (BRD) in 136 recently weaned cattle (“weanlings”), 6–12 mo of age, that were submitted for postmortem examination to regional veterinary laboratories in Ireland. A standardized sampling protocol included routine microbiologic investigations as well as polymerase chain reaction and immunohistochemistry. Lungs with histologic lesions were categorized into 1 of 5 morphologic patterns of pneumonia. Fibrinosuppurative bronchopneumonia (49%) and interstitial pneumonia (48%) were the morphologic patterns recorded most frequently. The various morphologic patterns of pulmonary lesions suggest the involvement of variable combinations of initiating and compounding infectious agents that hindered any simple classification of the etiopathogenesis of the pneumonias. Dual infections were detected in 58% of lungs, with Mannheimia haemolytica and Histophilus somni most frequently recorded in concert. M. haemolytica (43%) was the most frequently detected respiratory pathogen; H. somni was also shown to be frequently implicated in pneumonia in this age group of cattle. Bovine parainfluenza virus 3 (BPIV-3) and Bovine respiratory syncytial virus (16% each) were the viral agents detected most frequently. Potential respiratory pathogens (particularly Pasteurella multocida, BPIV-3, and H. somni) were frequently detected (64%) in lungs that had neither gross nor histologic pulmonary lesions, raising questions regarding their role in the pathogenesis of BRD. The breadth of respiratory pathogens detected in bovine lungs by various detection methods highlights the diagnostic value of parallel analyses in respiratory disease postmortem investigation.
Introduction
The term bovine respiratory disease (BRD) encompasses pneumonias in cattle caused by an array of infectious agents and environmental factors, resulting in a complex range of pulmonary lesions.24,41,45,55,60 Outbreaks of BRD result in significant economic loss worldwide.8,52 In Ireland, BRD is the most common cause of death recorded in cattle >1 mo of age (2012 and 2013 AFBI/DAFM all-island animal disease surveillance reports. Available at: goo.gl/ZNCtNg). Infectious causes include viral, bacterial, and mycoplasmal agents, some of which can be isolated from normal tissue of the bovine respiratory tract.22,24,25 However, the complex and confounding aspects of the pathogenesis of BRD (such as the role played by host or environmental factors) are illustrated by the fact that not all of the viruses and bacteria identified in pneumonic lungs are consistently implicated as either causative or contributory.41,43 Primary viral infection impairing host defenses and predisposing to secondary bacterial infection 31 is a preeminent pathogenesis, but viral, 43 bacterial, 18 mycoplasmal, 40 and parasitic 17 infections alone, as well as combinations thereof, have also been implicated.19,21,32
BRD in northern European countries such as Ireland is seasonal in occurrence (goo.gl/ZNCtNg) with a peak of cases between September and February. 41 This seasonality is related to the often complex interaction between climatic conditions and the effect on overall immunity of potentially stressful husbandry practices such as comingling and housing of recently weaned cattle.15,29,38,39,53
Despite the huge economic impact of BRD in Ireland and beyond,23,52 there are few detailed large-scale studies19,27 assessing the relative frequency with which the various bovine respiratory pathogens occur and the morphologic patterns of pneumonia that result under such climatic and husbandry conditions.6,9,10,41 We describe and compare the pathogens, patterns of pneumonia, and associated epidemiologic risk factors of BRD in cohorts of affected and unaffected weaned cattle presented for postmortem examination.
Materials and methods
Weanling selection and clinical information
A 13-wk study was conducted from September 16 to December 15, 2013 including cattle 6–12 mo of age (i.e., “weanlings”) that had been submitted for postmortem examination to any of the 6 Regional Veterinary Laboratories (RVLs) in the Republic of Ireland and that were deemed suitable for full diagnostic workup. An accompanying clinical history covering herd demographics, treatment history, vaccination history, husbandry conditions, and the clinical signs observed antemortem was obtained from herd owners on submission of the carcass. Using the tag and herd number of the submitted carcass, further herd- and animal-level data, including confirmation of age, was obtained by interrogation of the Animal Identification and Movement (AIM) system and Irish Cattle Breeders Federation databases. These data were collated with those provided by the herd owner.
Gross pathology
All carcasses were subjected to a standardized postmortem examination of the heart and lungs, and all analyses deemed appropriate to confirm a diagnosis of BRD were carried out. Regardless of the presence or absence of grossly visible pulmonary or cardiac lesions, a standard examination and tissue sampling protocol for respiratory disease diagnostic purposes were employed. Briefly, the trachea, lungs, and heart were removed, the lungs were palpated and serially incised, and the trachea, bronchi, and pulmonary arteries were incised longitudinally. The heart chambers were opened and examined and a left ventricular papillary muscle incised. All grossly visible lesions were described, and pulmonary lesions were mapped graphically on a diagram of the lungs (Fig. 1).
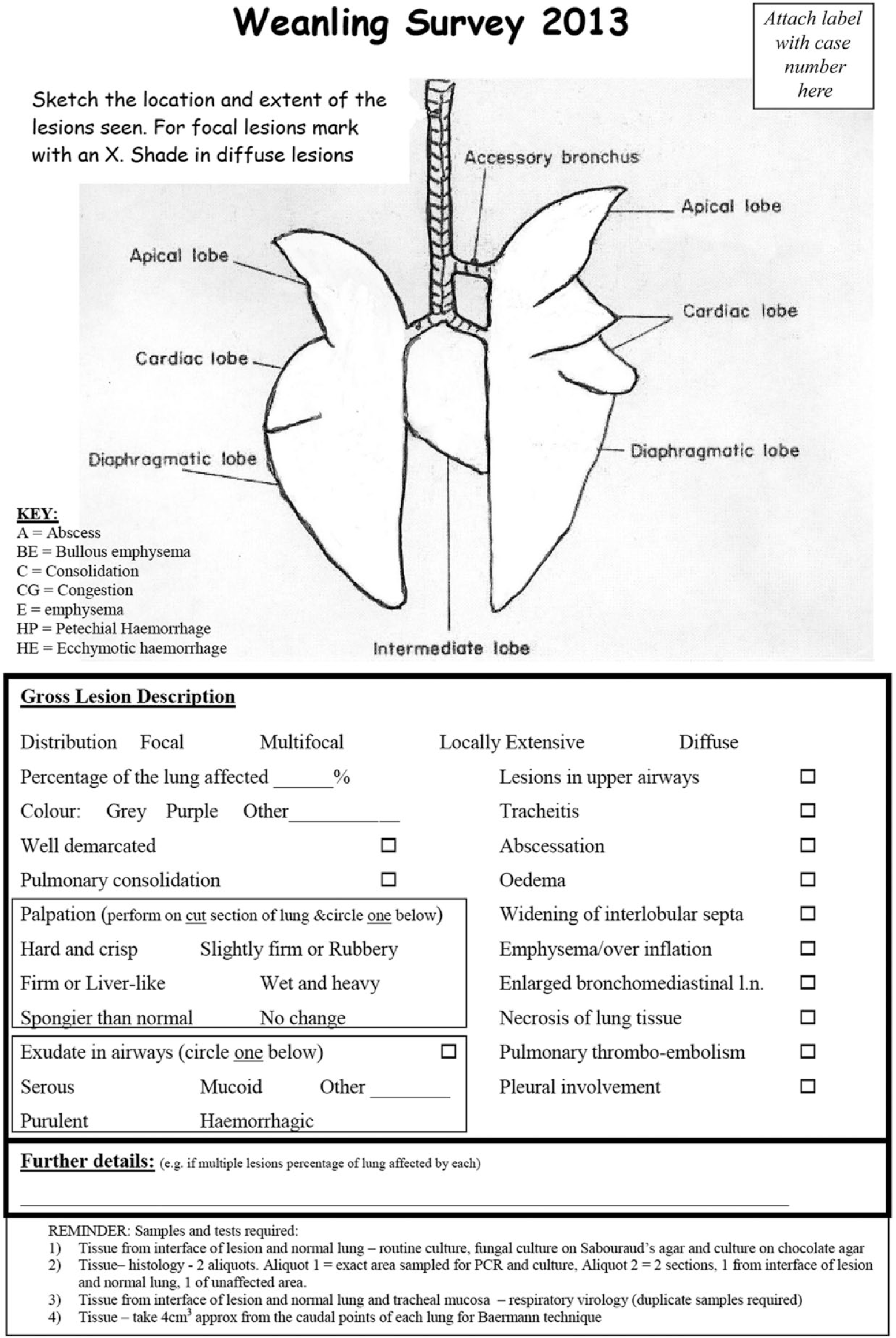
The working document used by pathologists on which grossly visible lesions in bovine lungs were mapped graphically and recorded categorically and by free-text.
Bacteriology, fungal culture, and virology
Aerobic bacteriology on blood agar was performed on all lung samples. In carcasses with grossly visible lung or heart lesions (n = 66), single lung samples from the border of lesioned and nonlesioned lung were submitted for culture on blood agar (37°C for 2 d), fungal culture on Sabouraud agar (37°C for 2 d), and chocolate agar culture (8% CO2 at 37°C for 3 d). Plates were read at 24-h intervals. For carcasses in which grossly visible lung or heart lesions were absent (n = 70), a single lung sample from a standardized location at the junction of the cranial and middle lobes was taken. Lung and tracheal mucosa were submitted for polymerase chain reaction (PCR) analysis for the detection of Mycoplasma bovisa,49 (n = 136), Histophilus somnia,6 (n = 133), Mannheimia haemolyticab–e,34 (n = 120), and Pasteurella multocidab–e,35 (n = 127), which was performed as previously described. Real-time PCRb,f,g analysis to identify Bovine herpesvirus 1 (BoHV-1; n = 136) and reverse transcription PCR h analysis to identify Bovine respiratory syncytial virus (BRSV), Bovine parainfluenza virus 3 (BPIV-3), Bovine coronavirus (BCoV), and Bovine viral diarrhea virus (BVDV; n = 136) were performed individually as described previously. 41 DNA or RNA extracted from laboratory strains of the target organisms using commercial DNA or RNA isolation kits (for M. bovis, i H. somni, i and respiratory virusesf,j,k) or specific commercially manufactured oligonucleotides e (for P. multocida and M. haemolytica) were used as positive controls; reaction mixtures without the template DNA or RNA were used as negative controls in all amplifications for each PCR.
Parasitology
Lung samples (n = 135) of ~4 cm3 from the caudal lobes of each lung were subjected to modified Baermann testing.5,28 Briefly, 8 cm3 of lung was placed in a blender l for 30 s and then suspended in a double layer of cheesecloth for 24 h in a funnel with lukewarm water. Five milliliters of fluid from the stem of the funnel were aliquoted into a test tube and left to sediment for at least 30 min. A drop of Gram iodine was added to a drop of sediment on a cover slide and examined for Dictyocaulus viviparus larvae.
Histopathology and immunohistochemistry
Lung samples (n = 199) were taken for histopathology from 133 animal carcasses. These samples included a single tissue section (n = 133) taken from the same location as the lung sampled for the microbiologic analyses described above. In those carcasses with grossly visible lung or heart lesions (n = 66), a second section was taken, which included tissue from the border of lesioned and non-lesioned tissue and from the least affected part of the lung. Sections were fixed in 10% neutral-buffered formalin for 4 d, embedded in paraffin wax, cut with a microtome, and stained with hematoxylin and eosin. Immunohistochemistry (IHC) for the detection of M. bovism–o,48 (n = 203 from 132 carcasses), H. somnim–o,9 (n = 204 from 132 animal carcasses), and M. haemolyticam–o (n = 195 from 130 animal carcasses) was performed using standard methods. Briefly, paraffin-embedded sections were cleared, and sections were incubated with 0.5% H2O2 in methanol for 20 min, followed by treatment with 0.1% protease type XIV m at room temperature for 5 min. Sections were then washed in Tris-buffered saline (TBS) and incubated in a solution of 0.1% bovine serum albumin (BSA) in TBS for 20 min. The sections were then placed in a protein block o for 5 min. Following washing in TBS for 5 min, sections were then incubated at 1/3,000 in polyclonal antibody overnight at room temperature. Sections were then treated with a blocking reagent for endogenous peroxidase p and a high-sensitivity 3-amino-9-ethylcarbazole (AEC+) chromogenic substrate system at room temperature. The sections were incubated in a vector peroxidase substrate q for 5 min, after which they were counterstained with hematoxylin.
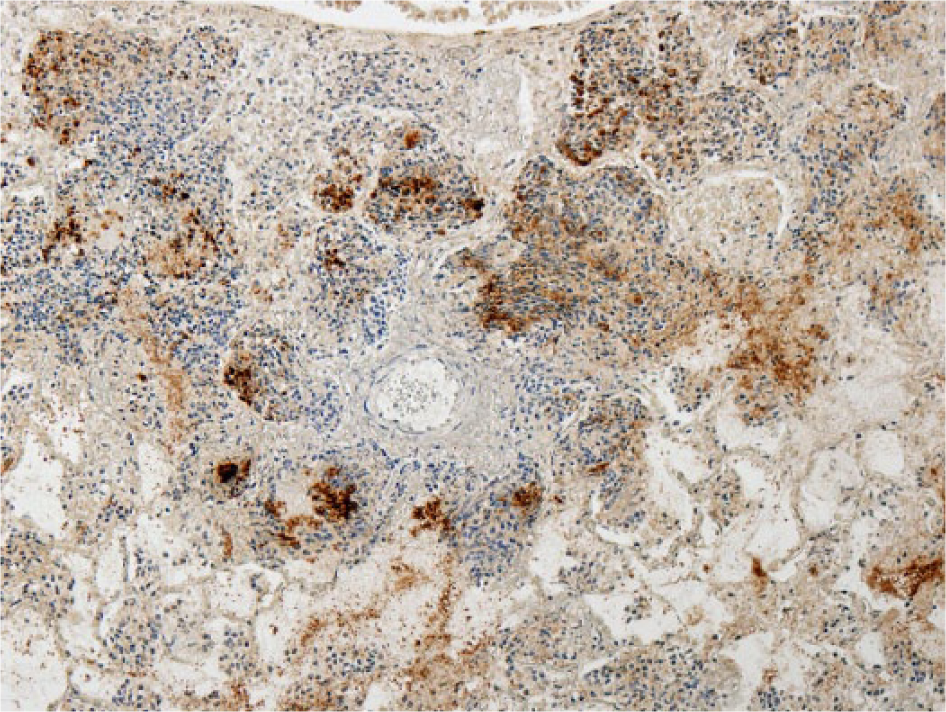
The primary antibodies and antisera employed for all 3 IHC tests were produced in-house, and the specificity of each for their respective pathogens was evaluated previously.4,9,48 Briefly, M. bovis antigen was detected using a monoclonal antibody (mAb5A10) raised against an M. bovis–specific 30 kDa protein. mAb5A10 did not cross-react with lung tissue infected with M. haemolytica, P. multocida, or Mycoplasma mycoides ssp. mycoides small colony. 48 Omission of primary antibodies and the use of lung tissue infected exclusively with M. bovis (confirmed by culture and PCR) constituted negative and positive control procedures, respectively. Histophilus somni antigen was detected using a polyclonal antiserum raised against the H. somni–specific 270K Fc receptor antigen. This antiserum did not cross-react with P. multocida, M. haemolytica biotype A serotypes 1 and 2, or Trueperella pyogenes antigens. 9 Omission of primary antibodies and the use of lung tissue infected exclusively with H. somni (confirmed by culture) constituted negative and positive control procedures, respectively. Mannheimia haemolytica antigen was detected using a polyclonal antiserum raised against the outer membrane protein of serotype A2. The specificity of the antisera was tested by indirect hemagglutination and by rapid plate agglutination. A high homologous titer (≥1:256) with negligible cross-reaction (<1:16) was obtained by indirect hemagglutination. The low-level cross-reactivity of the antisera to the A-type strains by rapid plate agglutination was successfully eliminated by dilution. 4 Omission of primary antibodies and the use of lung tissue infected exclusively with M. haemolytica (confirmed by culture) constituted negative and positive control procedures, respectively. Histologic examination of IHC sections was undertaken with reference to positive and negative controls. Histologic examination of all sections (n = 204) was performed by 1 operator (GM Murray), and morphologic diagnoses determined by the predominant pattern were based on the 6 morphologic patterns outlined in Table 3.
Epidemiologic analysis
The “straight-line” distance of the submitting herd from the RVL was calculated using the X and Y coordinates of the RVL and herd as recorded on the Land Parcel Identification System for 121 of the 124 herds. For the remaining 3 herds, for which the exact location was unknown, these calculations were based on the X and Y coordinates of the centroid for the district electoral division in which the farm was located. Where available, farmer-reported information about animal vaccine status (n = 108) and antemortem antimicrobial usage (n = 119) was obtained. In Table 1, “respiratory vaccination” refers to vaccination against BoHV-1, BRSV, BPIV-3, BVDV, BCoV, M. haemolytica, P. multocida, or any other recognized primary bovine respiratory pathogen, either alone or in combination. Data were obtained from the AIM database on the total number of on-farm deaths and the average monthly herd size of submitting herds during 2013. A crude annual mortality rate was calculated by dividing the annual number of on-farm deaths for 2013 by the average monthly herd size over the entire year. The number of deaths in a respiratory disease outbreak was calculated as the total number of on-farm deaths of all ages in the submitting herd, including the submitted animal, during the 30 d preceding and 30 d subsequent to the death of the submitted animal.
Epidemiologic categorical variables of weaned cattle (n = 136), submitted to the Regional Veterinary Laboratories in the Republic of Ireland, both with (n = 66) and without (n = 70) visible lung or heart lesions. As heart lesions were not recorded in any animal, visible lesions refer to lung lesions only.
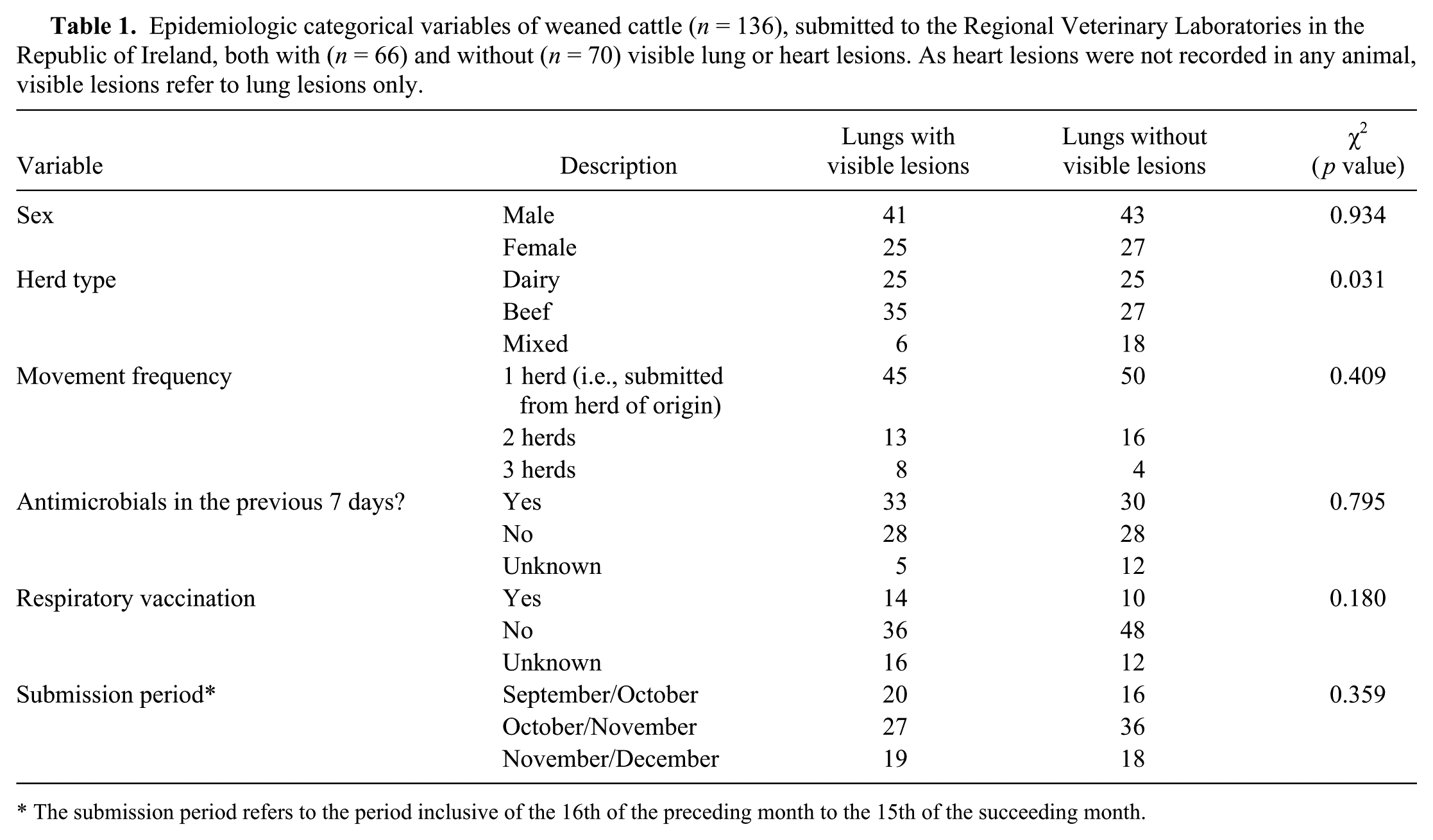
The submission period refers to the period inclusive of the 16th of the preceding month to the 15th of the succeeding month.
Statistical analysis
A statistical software package r was used for all data management and statistical analyses. The association between the presence of lung lesions and specific categorical variables was tested using a chi-square test, unless the numbers within a category were less than 5 in which case the Fisher exact test was used.
Results
Descriptive epidemiology of the study population
A total of 136 cattle from 124 herds were examined postmortem (Table 1). Two animals had been euthanized prior to submission; the remaining 134 animals had succumbed to clinical disease. Eighty animals (59%) were submitted without any clinical histories. Twenty-three animals (17%) had histories of respiratory signs (only 6 of which were reported to have exhibited signs for more than 7 d before death), 19 animals (14%) were reported to have died suddenly without clinical signs, and 9 animals (7%) were reported to have clinical signs suggestive of enteric disease. All 136 animals were born and spent their entire lives in the Republic of Ireland. There was a significant ( p = 0.03) association between herd type and the occurrence of lung lesions in the study population. Mixed herds had a lower incidence of lung lesions than expected, whereas beef herds had a higher incidence than expected. Carcasses were submitted from herds in 20 of the 26 counties in the Republic of Ireland (Fig. 2). The mean herd size of those animals with visible lung lesions was 171 animals, and the average annual crude herd mortality rate (total fallen animals within 12 mo/average herd size over 12 mo) within these herds during the 12 mo prior to the death of the animal was 8.2%. The mean number of bovine deaths recorded in submitting herds during respiratory disease outbreaks was 3.6.
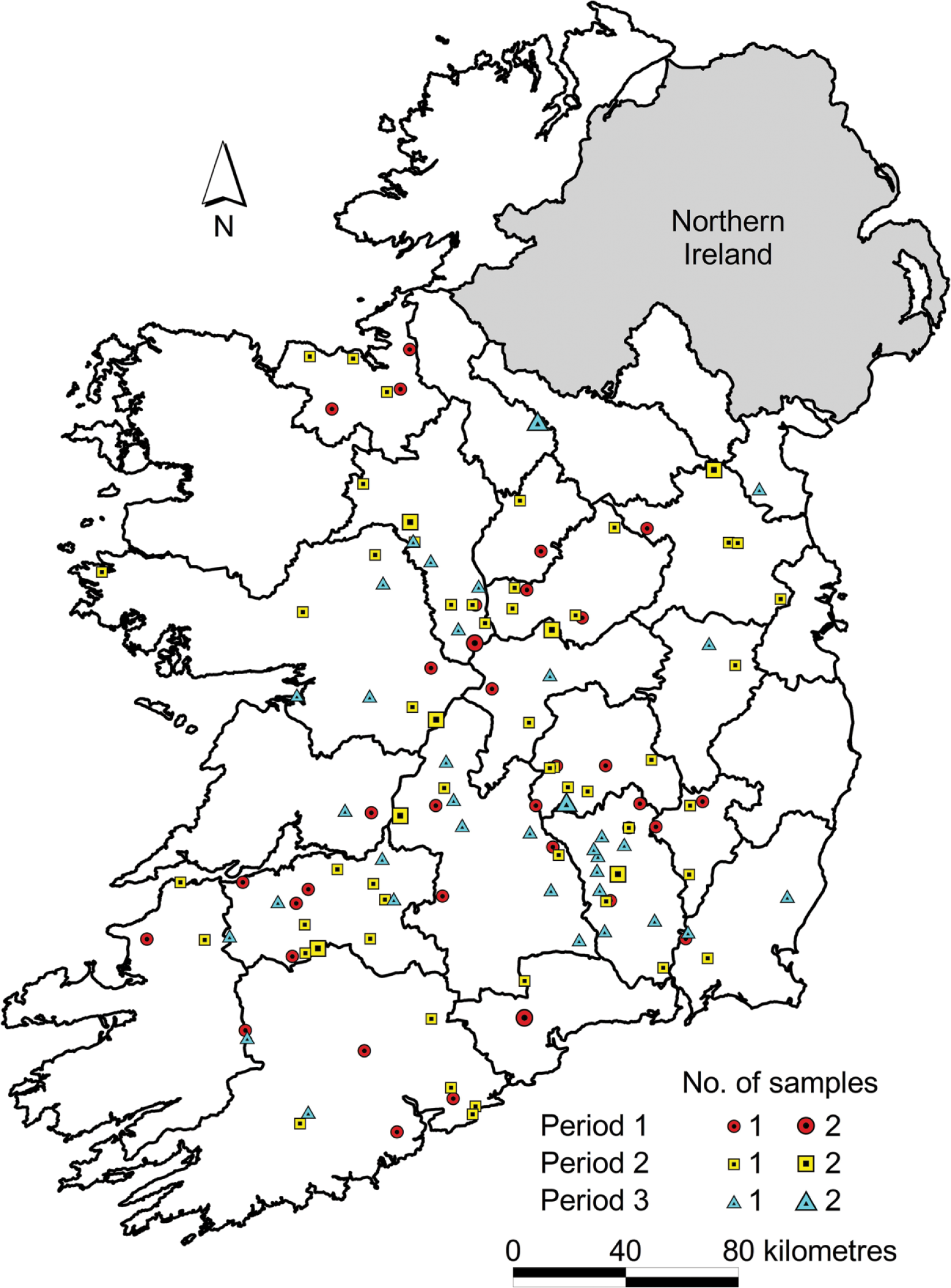
A map of Ireland showing the temporal and spatial distribution of 136 weanling bovine carcass submissions to the Regional Veterinary Laboratories between September 16, 2013 and December 13, 2013. Period 1 = 16 Sept–15 Oct; Period 2 = 16 Oct–15 Nov; Period 3 = 16 Nov–15 Dec.
Gross pathology
Grossly visible pulmonary lesions were recorded in 66 animals. Microscopically visible lesions, with interstitial pneumonia predominating (11 animals), were identified in an additional 15 animals in which grossly visible pulmonary lesions were not recorded. Gross lesions were not detected in the heart of any animal. Of these 81 animals in which histologic pulmonary lesions were recorded, respiratory disease was identified as the primary cause of death in 51. Gastrointestinal disease (10 animals), septicemia (6 animals), and blackleg (5 animals) were the other commonly recorded causes of death. Purple, firm, well-demarcated consolidation was a typical finding in the 40 animals in which bronchopneumonia was recorded. Only 1 of these 40 animals had histologic but not grossly visible lesions. In 19 of the 39 animals with grossly visible lesions, the lesional proportion of lung was >66%; in only 9 animals were lesions confined to the cranial and middle lobes. Lesions were most commonly recorded as locally extensive in distribution (17 of 39).
Abscessation (i.e., an accumulation of liquefied-to-more-inspissated exudate within a tissue space) was a particular feature of caseonecrotic bronchopneumonias. Grossly visible lesions were recorded in the lungs of all 5 cases recorded, with a locally extensive distribution most commonly recorded (4 of 5). Cranial and middle lobes had lesions in all 5; 3 animals also had lesions in the caudal lobes. Only 2 animals had a lesional proportion of >50%.
Emphysema was a frequent finding in lungs in which bronchointerstitial pneumonia was identified. Of the 25 animals in which bronchointerstitial pneumonia was recorded, 23 had grossly visible lesions, with a lesional proportion of ≥80% in 14 of these 23. Lesions were distributed across cranial, middle, and caudal lobes in 19 animals.
Many interstitial pneumonic lungs were purple and rubbery. Eleven of the 38 lungs in which interstitial pneumonia was recorded histologically did not have grossly visible lesions. A diffuse pulmonary lesional distribution was recorded in 17 of these 27 animals in which grossly visible lesions were recorded, although in only 10 animals was the lesional proportion of lung >80%. The cranial lobes had lesions in all 27 animals, although in only 2 animals were lesions confined to the cranial lobes.
Prevalence of selected potential respiratory pathogens in lung tissue
Pasteurella multocida was isolated by bacterial culture of the lung in 8 animals with grossly visible lung lesions, 6 of which were also positive on PCR. Pasteurella multocida nucleic acid was detected by PCR in lung tissue from a further 17 animals from which P. multocida was not isolated on bacterial culture (Table 2).
The frequency of detection of specific pathogens by different means of detection, in the lungs of bovine weanlings with (n = 66) and without (n = 70) lung lesions on postmortem examination.*
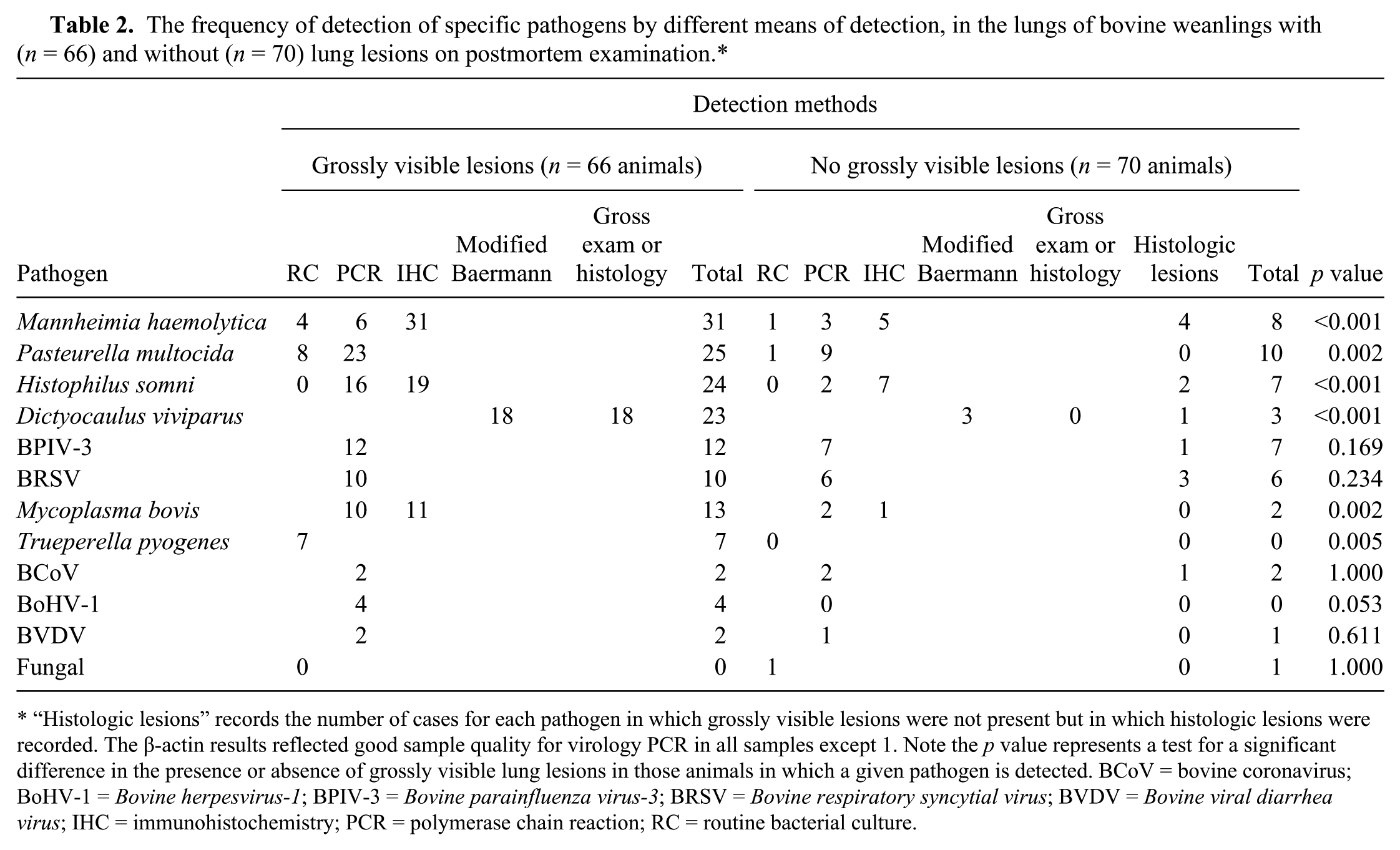
“Histologic lesions” records the number of cases for each pathogen in which grossly visible lesions were not present but in which histologic lesions were recorded. The β-actin results reflected good sample quality for virology PCR in all samples except 1. Note the p value represents a test for a significant difference in the presence or absence of grossly visible lung lesions in those animals in which a given pathogen is detected. BCoV = bovine coronavirus; BoHV-1 = Bovine herpesvirus-1; BPIV-3 = Bovine parainfluenza virus-3; BRSV = Bovine respiratory syncytial virus; BVDV = Bovine viral diarrhea virus; IHC = immunohistochemistry; PCR = polymerase chain reaction; RC = routine bacterial culture.
In 41 carcasses, no potential respiratory pathogen was identified in the lungs; 25 of these animals had neither gross nor histologic lung lesions on examination, whereas 8 animals had histologic but not grossly visible lesions. Detection of multiple respiratory pathogens in the lungs was a common feature (data not presented), with coinfections of H. somni and M. haemolytica (18 animals) and P. multocida and M. haemolytica (17 animals) recorded with greatest frequency. In 42 animals, only a single respiratory disease pathogen was detected in the lungs. P. multocida was the pathogen identified alone in lungs with the greatest frequency (9 animals). Of 66 animals recorded with grossly visible lung lesions, 28 animals had 3 or more potential respiratory pathogens detected. The most frequently detected viral and bacterial co-infections were P. multocida and BPIV-3, H. somni and BPIV-3, or H. somni and BRSV (each detected in the lungs of 7 weanlings).
Histopathologic findings
The morphologic categorization of the pulmonary pathology and the corresponding pathogens identified are presented in Table 3. More than 1 specific histologic category of pneumonia was identified in some lungs.
The gross postmortem findings and respiratory pathogens identified in bovine weanling lungs recorded as exhibiting histologic lung lesions (n = 81), categorized according to the histologic findings recorded. Note that n = 66 had grossly visible lung lesions and n = 15 did not have grossly visible lung lesions. More than 1 specific histologic category or pathogen was identified in some lungs.*
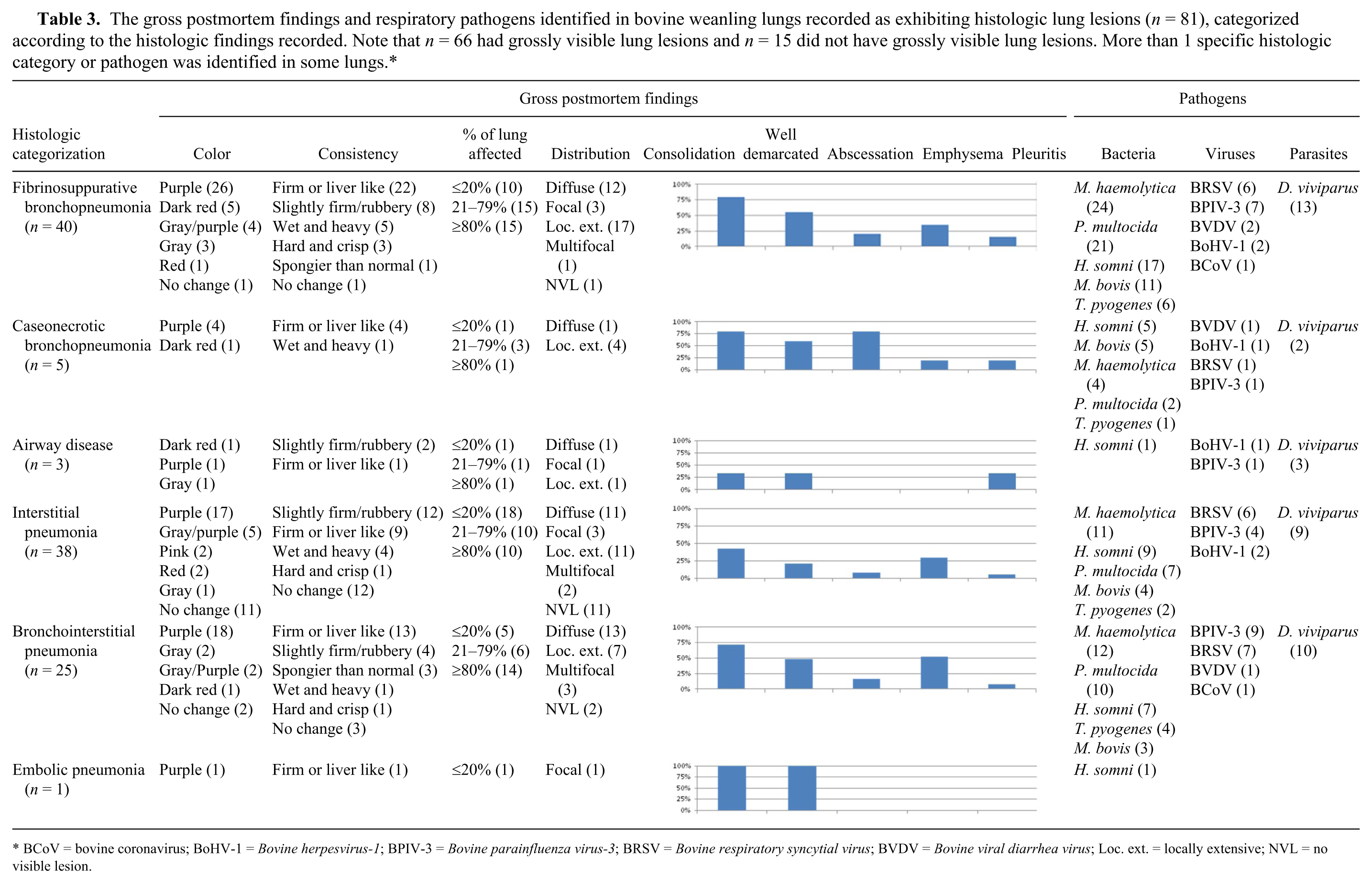
BCoV = bovine coronavirus; BoHV-1 = Bovine herpesvirus-1; BPIV-3 = Bovine parainfluenza virus-3; BRSV = Bovine respiratory syncytial virus; BVDV = Bovine viral diarrhea virus; Loc. ext. = locally extensive; NVL = no visible lesion.
Pneumonia was categorized as “bronchopneumonia” (Fig. 3) when inflammation of the lung tissue was characterized by exudate in the airspaces extending typically from the bronchoalveolar junction. Lesions of fibrinous and/or suppurative bronchopneumonia were grouped together as fibrinosuppurative bronchopneumonia, which was the largest morphologic category recorded (40 animals) and was almost consistently associated with grossly visible lesions (39 of 40). Fibrin deposition (39 of 40 animals), blood vessel or lymphatic thrombosis (19 of 40), and fibrinous pleuritis (2 of 40) were features observed in cases of bronchopneumonia. Of the lungs in which blood vessel or lymphatic thrombosis were identified, M. haemolytica (4) was identified in 14, H. somni in 12, P. multocida in 9, and M. bovis in 7. Chronic lesions were identified in 28 of the 40 animals with fibrinosuppurative bronchopneumonia, with obliterative bronchiolitis (24) the most commonly recorded chronic lesion; peribronchial fibrosis was identified in 19 animals. In total, 30 lungs in which fibrinosuppurative bronchopneumonia were recorded were detected with either M. haemolytica (n = 10) or P. multocida (n = 7), or both (n = 13). Of these 30 animals, “oat cells” (Fig. 4) were identified in the lungs of 10 animals, 6 of which proved positive for M. haemolytica only and 4 which were positive for both pathogens. Five (2 with chronic lesions) of these 10 animals had not been treated with antimicrobials in the 7 d before death; antimicrobials had been administered to 3 animals (1 with chronic lesions). “Oat cells” were recorded in only 1animal in which neither M. haemolytica nor P. multocida were identified; this animal, diagnosed with acute fibrinosuppurative bronchopneumonia, had not been treated with antimicrobials.
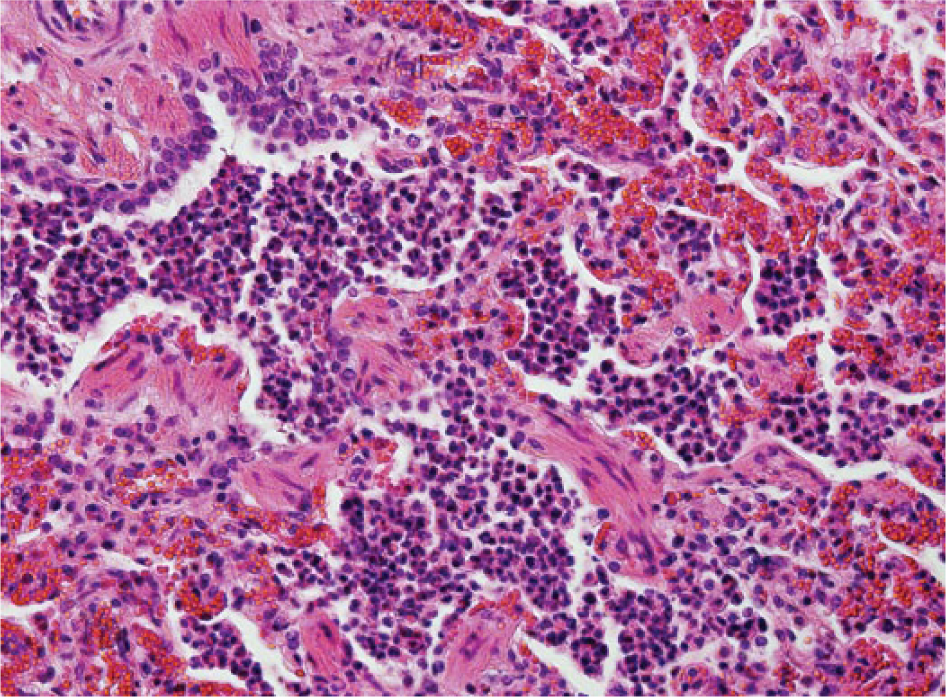
Bovine lung with acute diffuse suppurative bronchopneumonia characterized by abundant neutrophils in the lumen of bronchioles and at the bronchoalveolar junction. Alveoli contain viable, and some degenerate, neutrophils as well as intact alveolar macrophages. Hematoxylin and eosin. 200×.
Bovine lung with immunohistochemical staining specific for Mannheimia haemolytica within multifocal aggregates of degenerate leukocytes with nuclear streaming (“oat cells”) in the terminal bronchioles and alveoli. 100×.
Mannheimia haemolytica (n = 24), P. multocida (n = 21), and H. somni (n = 17) were detected in cases of fibrinosuppurative bronchopneumonia with greatest frequency; BPIV-3 and BRSV were the viral agents detected most frequently (7 and 6 animals, respectively).
Mycoplasma bovis was detected in 11 animals in which fibrinosuppurative bronchopneumonia was recorded, but only 5 of these had concurrent caseonecrotic foci (Fig. 5). Ten of these 11 animals were also positive for M. bovis antigen on IHC (Fig. 6). All 5 cases of caseonecrotic bronchopneumonia were positive for H. somni antigen (Fig. 7) or nucleic acid (4 of 5 on IHC, 4 of 5 on PCR) and M. bovis (5 of 5 on IHC and PCR). In the 6 cases of fibrinosuppurative bronchopneumonia without caseonecrotic foci, M. bovis antigen was primarily located in degenerate inflammatory cells in the bronchiolar lumen. Two lungs without histologic lesions were positive for M. bovis nucleic acid by PCR, in 1 of which M. bovis antigen was detected by IHC in a localized area of the alveolar wall and lumen.
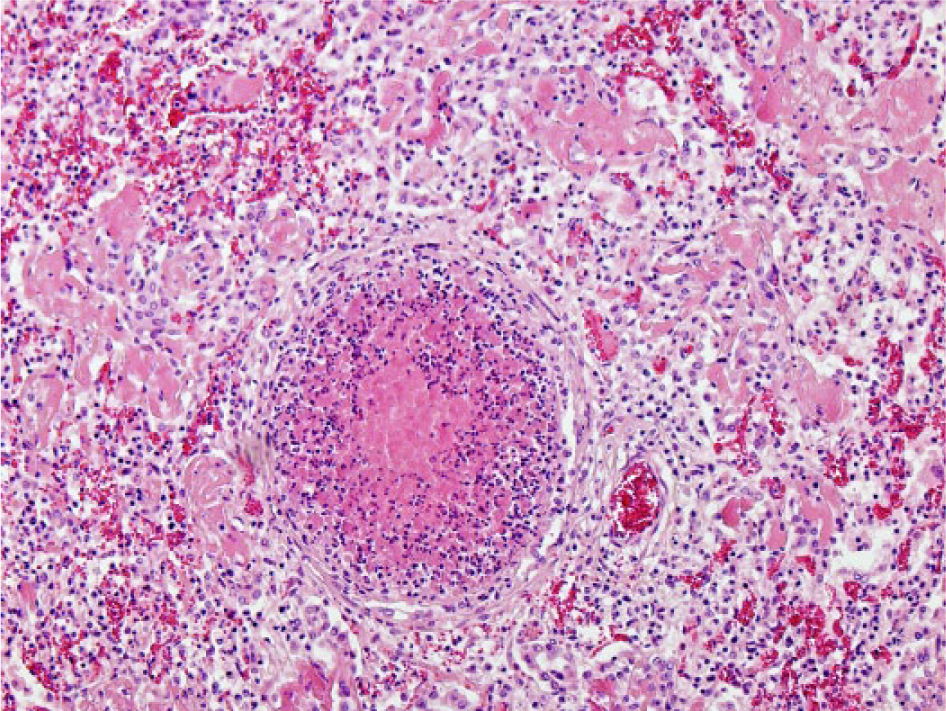
Bovine lung with caseonecrotic bronchopneumonia characterized by an ectatic bronchiole filled with amorphous eosinophilic material and demarcated by degenerate inflammatory cells and remnants of necrotic bronchiolar epithelium. Hematoxylin and eosin. 100×.
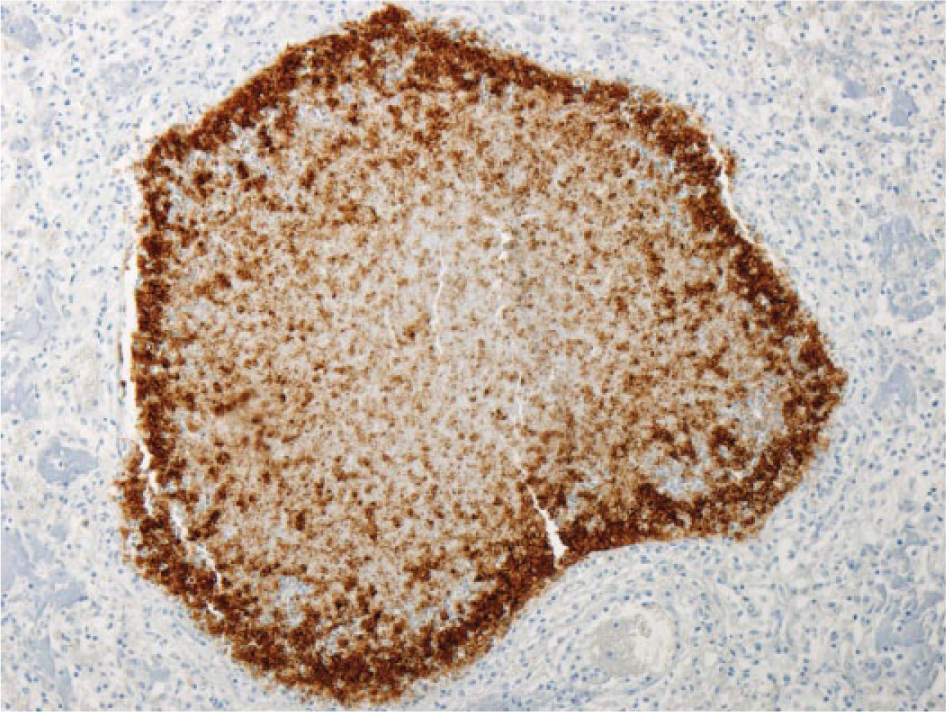
Bovine lung with immunohistochemical staining specific for Mycoplasma bovis in bronchiolar epithelium and within degenerate material within the bronchiolar lumen. 100×.
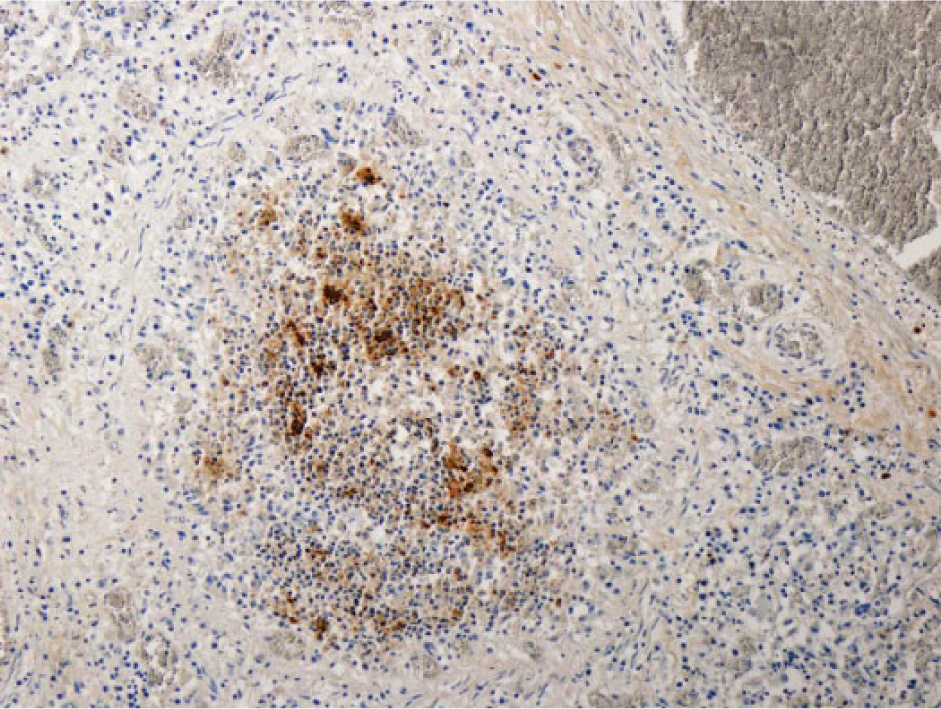
Bovine lung with immunohistochemical staining specific for Histophilus somni in macrophages, degenerate neutrophils, and necrotic bronchiolar epithelial cells in an ectatic bronchiole. 100×.
Histophilus somni antigen was detected in 13 lungs in which fibrinosuppurative bronchopneumonia was recorded. Nine of these 13 animals had abundant antigen primarily in macrophages and neutrophils in the bronchiolar lumina, in bronchiolar epithelial cells, and in the alveolar macrophages. The remaining 4 lungs recorded scant antigen primarily located in the alveolar macrophages. Seven of the 13 animals were also positive for H. somni nucleic acid on PCR. Histophilus somni antigen was detected in the bronchiolar epithelium of 5 lungs without histologic lesions; the detection of antigen in 3 of these 5 lungs was scant. Only 1 of these 5 lungs was also positive for H. somni nucleic acid by PCR. Thirteen animals were positive for the presence of H. somni nucleic acid by PCR and H. somni antigen by IHC. Among these 13 animals, 4 different histologic patterns of pneumonia were recorded: fibrinosuppurative bronchopneumonia (n = 7), interstitial pneumonia (n = 6), bronchointerstitial pneumonia (n = 4), and caseonecrotic bronchopneumonia (n = 3). Abundant H. somni antigen was present in 9 of these 13 animals, primarily located in the bronchiolar lumen within neutrophils and macrophages, in bronchiolar epithelium, and in the alveolar macrophages. Four or more respiratory pathogens were detected in 11 of these 13 animals. One animal without histologic lesions was also positive for H. somni nucleic acid by PCR and H. somni antigen by IHC. There was scant antigen in the lungs of this animal. The histologic categorization of “interstitial pneumonia” was given to inflammation of the “nonairway” components of the lung (i.e., inflammation that is focused primarily on the alveolar walls; Fig. 8), and was recorded in 38 animals. Hyaline membrane formation (15 animals) and type 2 pneumocyte hyperplasia (epithelialization; 11 animals) were regularly identified features in the lungs of these animals. Eleven of the 38 animals in which interstitial pneumonia was recorded did not have grossly visible lung lesions. Eleven separate diagnosed causes of death were recorded in these animals, including septicemia, cellulitis, meningitis, and nephritis.
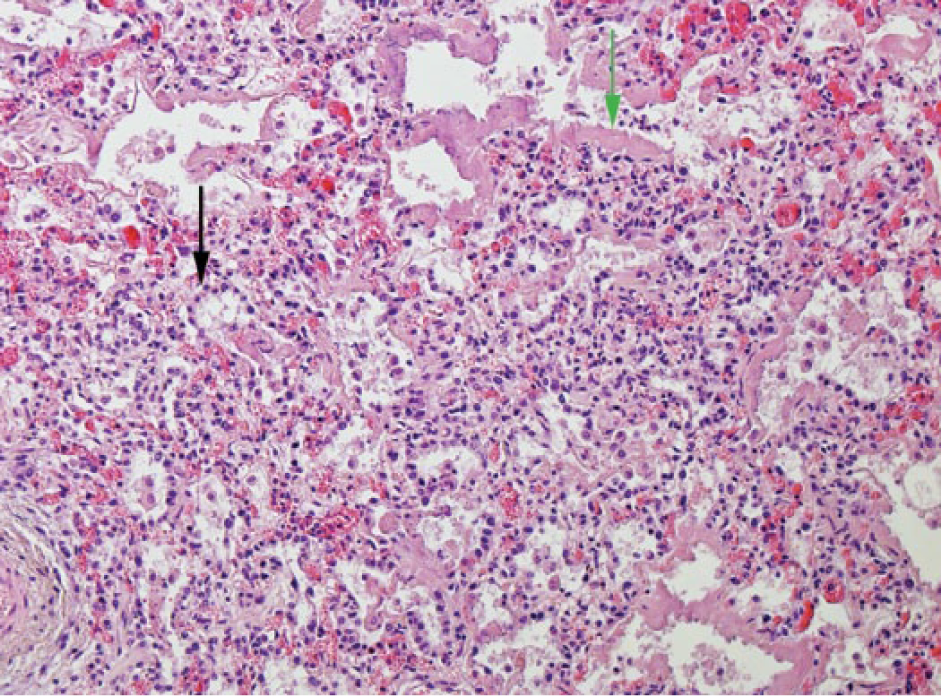
Bovine lung with acute diffuse interstitial pneumonia, with edema, hyaline membranes (green arrow), type II pneumocyte hyperplasia (black arrow), congestion, and expansion of the alveolar walls by a mild infiltrate of mononuclear inflammatory cells. H&E. 100×.
“Bronchointerstitial pneumonia” (Fig. 9) was the histologic categorization given to inflammation of the lung resulting from injury to both the bronchiolar and alveolar epithelium and was diagnosed in 25 animals, 23 of which had grossly visible lung lesions. Fibrin (23 animals), hyaline membranes (n = 18), and syncytial cells (n = 11) were common histologic features.
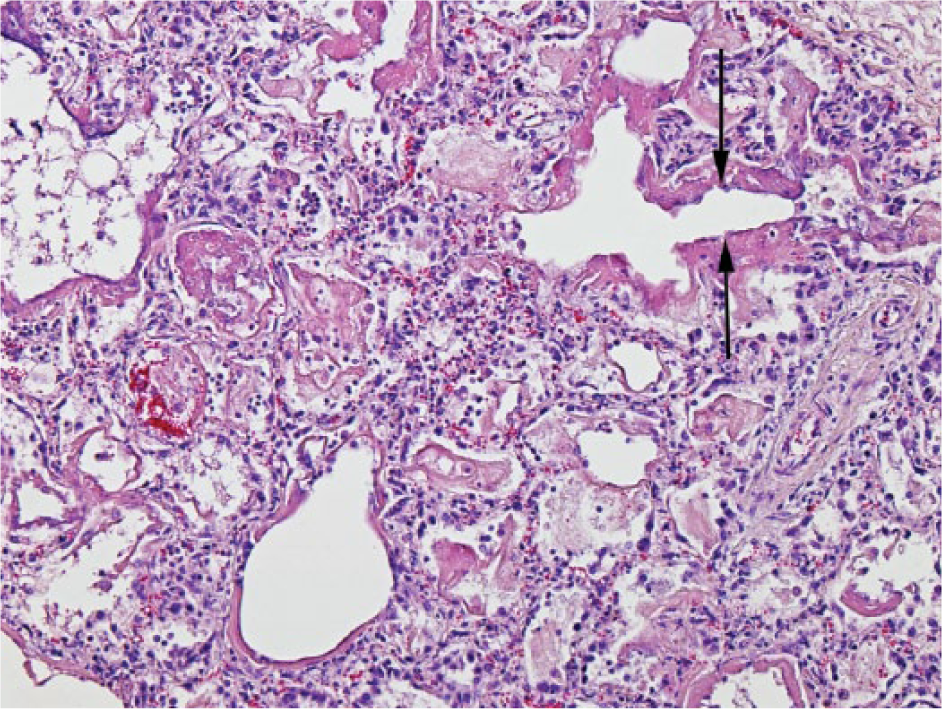
Bovine lung with bronchointerstitial pneumonia characterized by necrotic and attenuated bronchiolar epithelium (arrows). There is type II pneumocyte hyperplasia evident in surrounding alveoli, fibrin deposition, and hyaline membranes. There are occasional prominent alveolar macrophages in the lumen. H&E. 100×.
Although exudate in the airway lumens is the primary feature of bronchopneumonia, epithelial necrosis is rarely seen. In contrast, bronchointerstitial pneumonia is characterized primarily by epithelial necrosis with occasional mild exudate in the airspaces. Lesions of both bronchointerstitial pneumonia and bronchopneumonia were identified in 12 weanlings. The bronchopneumonia was identified as fibrinosuppurative in all 12 animals; additional lesions of caseonecrotic bronchopneumonia were identified in 1 weanling.
Airway disease was diagnosed in 3 cases. This refers to a morphologic pattern of bronchitis, bronchiolitis, or both, without the involvement of alveolar epithelial cells.
Lesions of chronic pneumonia such as epithelial hyperplasia, pulmonary fibrosis, obliterative bronchiolitis, bronchiectasis, abscessation, or sequestration were identified in 40 weanlings. Obliterative bronchiolitis was identified in 34 animals. Seven of these animals only had lesions of bronchopneumonia, 7 animals exhibited lesions of bronchointerstitial pneumonia only, and 8 animals exhibited lesions of both bronchopneumonia and bronchointerstitial pneumonia. Twenty-seven of the 40 animals with chronic lung lesions had been treated with antimicrobials within the 7 d before death, whereas 12 had not. In 3 of these 40 animals, a respiratory pathogen was not detected in the lungs; 17 animals had 3 or more pathogens detected. Pasteurella multocida was detected in 22 animals; M. haemolytica was detected in 21; H. somni (n = 14) and D. viviparus (n = 14) were detected with lesser frequency. BPIV-3 (n = 10) and BRSV (n = 5) were the viruses most frequently detected in chronic cases.
Mannheimia haemolytica antigen was identified by IHC in the lungs of 25 animals that were negative for M. haemolytica nucleic acid on PCR. Antigen was plentiful in 17 of these lungs, primarily located in macrophages and neutrophils in the bronchiolar lumen and occasionally detected in bronchiolar epithelium or alveolar macrophages.
Discussion
The aim of this study was to address deficits in our knowledge of the pathogens, patterns of pneumonia, and associated epidemiologic risk factors of BRD in weaned cattle presented for postmortem examination during outbreaks of respiratory disease. The various morphologic patterns of lesions found in the lungs of calves that succumbed to pneumonia suggest involvement of variable combinations of initiating and compounding infectious agents. This complex etiology, taken together with the likely variable chronology with which the various pathogens participate in tissue injury, inevitably hinder any simple classification of the pneumonias present in terms of etiopathogenesis. 56 The time taken for pulmonary injury and inflammation to progress to the point where calves develop significant respiratory distress and die also likely means some loss in the diagnostic specificity of the morphologic changes as the lesions progress to “end stage.” 19 The variation in morphologic patterns of pneumonia within a single lung also present challenges to the categorization of pneumonia based on 1–2 histologic sections. However, despite these confounding events and recognized limitations, we were able to make some associations between particular pathogens and well-documented patterns of pneumonia.
As would be expected, based on previous studies of pneumonia in this age category of calf, 56 cases of fibrinosuppurative bronchopneumonia predominated and, although causation cannot be assumed, we found these were associated particularly with M. haemolytica (24 of 40), P. multocida (21 of 40), and H. somni (17 of 40). All 5 cases of caseonecrotic bronchopneumonia were associated with H. somni and M. bovis. Interstitial pneumonia was commonly linked with M. haemolytica (12 of 39), H. somni (9 of 39), and D. viviparus (9 of 39), and bronchointerstitial pneumonia was diagnosed in 25 animals, in which M. haemolytica (12 of 25), P. multocida (10 of 25), and D. viviparus (10 of 25) were most frequently encountered. The most exclusive association between specific pathogen(s) and pneumonia subcategory was in caseonecrotic bronchopneumonias in which H. somni and M. bovis were consistently identified, and in airway disease in which D. viviparus was constantly implicated. Histophilus somni was the only pathogen identified in each of the 5 histologic categories of pneumonia recorded. Embolic pneumonia, represented by only 1 case, was the only category in which H. somni nucleic acid was identified by PCR, but H. somni antigen was not identified by IHC.
Dual infections were detected in 47 of the 81 lungs (58.0%) in which lesions were recorded either on gross or histologic examination, which was more than the 45.3% recorded in a 2014 study confined to the detection of bacteria in lung tissue by PCR and culture. 6 The 2014 study identified co-infections most frequently between M. haemolytica and P. multocida; coinfections between P. multocida and M. bovis were detected marginally less frequently. In our study, M. haemolytica and H. somni were most frequently identified together, with either of these pathogens also frequently found in concert with P. multocida or M. bovis. There is evidence of synergistic relationships between pathogens causing BRD: P. multocida and M. bovis,14,58 M. bovis and M. haemolytica,11,23 BVDV and M. bovis,26,33 BVDV and BRSV, 36 BoHV-1 and BVDV, 44 and BRSV and H. somni. 12 The frequency of detection of H. somni in our study (32.1% of animals with grossly or histologically visible lesions) exceeds that recorded in postmortem submissions in Northern Ireland 6 (24.7%) and the United States 59 (19%). Furthermore, the detection rate far exceeds that recorded in feedlot populations in the United States 19 (10%) and Canada 21 (14.1%), although bacterial culture only was employed in both of these studies. Although bacterial culture failed to isolate H. somni from any of the lungs in our study, this pathogen was identified by PCR and IHC, a scenario similar to previous studies.6,42,43 Histophilus somni nucleic acid as well as H. somni antigen was detected in the lungs of 13 animals. In only 1 of these 13 (the single animal without histologic lesions) was H. somni detected alone without co-infecting pathogens. Histophilus somni can be detected in the lungs of healthy animals 1 ; however, the marked abundance of H. somni antigen in lesioned lung compared to non-lesioned lung in our study supports a pathogenic role in these animals. Furthermore, our findings challenge the view that H. somni pneumonia is primarily a disease of intensive cattle husbandry systems 51 such as feedlots.
In our study, M. haemolytica (43%) was the most frequently detected respiratory pathogen in the lungs of weanlings with either histologically or grossly visible lesions; P. multocida was detected at lesser frequency (31%). IHC accounted for many detections of M. haemolytica antigen in lungs in which nucleic acid was not detected on PCR. The PCR screens samples for virulence genes for leukotoxin (lktC) and UDP-N-acetyl-
Using a combination of PCR and IHC, M. bovis was identified from lungs with either histologic or grossly visible lung lesions at a frequency similar (16.0%) to previous studies, 10 but in our study all pneumonic lungs in which M. bovis was detected were also coinfected with at least 1 other respiratory pathogen. The detection of M. bovis in lung tissue in our study was significantly associated with lung lesions.
Respiratory viruses play an important role in the pathogenesis of BRD in compromising pulmonary defenses thereby facilitating bacterial colonization. 3 BPIV-3 and BRSV were the viruses most frequently detected in lungs with either grossly or histologically visible lung lesions—16% each—whereas BoHV-1 (4.9%), BCoV (3.7%), and BVDV (2.5%) were detected considerably less frequently. The role of BVDV in BRD in an Irish context does not appear to be as significant as previously assumed. 41 This is a likely consequence of the industry-led national BVD eradication program launched in 2012. The role of BCoV in the development of BRD is also unclear. 43 Within specific outbreaks, mortality resulting from respiratory BCoV infection can be high 54 ; however, the finding of our study supports the view 37 that BCoV may cause high morbidity 41 but is not a significant cause of mortality in cattle.
The detection of potential respiratory pathogens (particularly P. multocida, BPIV-3, and H. somni) in the lungs of 35 animals that had neither gross nor histologic pulmonary lesions is an interesting finding. Many bacterial pathogens are commensals that may preferentially colonize the lower respiratory tract.14,21–23 There are conflicting accounts of the roles of bacterial pathogens such as P. multocida1,2,14 (Maheswaran SK, et al. Current knowledge on the pathogenesis of lung injury caused by Mannheimia haemolytica and Pasteurella multocida in the bovine. Proc XXII World Buiatrics Congress; 2002 Aug 18–23; Hannover, Germany) and M. bovis11,20,33 as either primary causative or opportunistic pathogens in BRD pathogenesis. The ability of bovine paramyxoviruses (particularly BRSV) to persist in cattle has been proposed,7,57 and BPIV-3 can be isolated from individual calves for months following natural infection. 16 Previous resolved BPIV-3 infection may provide an explanation for our findings in some of these weanlings.
The adapted Baermann technique used in our study is useful in detecting D. viviparus in its prepatent phase.5,28 Dictyocaulus viviparus was the sole pathogen identified in 7 animals with grossly visible lesions. Our data indicate that this parasite was not significant in the development of pneumonia in all cases in which it was detected; larvae were found in 2 lungs in which grossly or histologically visible lesions were not identified.
The temporal proximity of weaning of beef calves, their sale through markets, and the housing of both dairy and beef calves are recognized stressors 15 and contribute to outbreaks of BRD in the autumn which precipitated the decision to perform this study during these months. A significant association was identified in our study between herd type and the occurrence of pulmonary lesions. No significant association was identified between other selected potential herd or animal BRD risk factors and the occurrence of pulmonary lesions. Although the sample size in our survey is comparable to other similar prevalence studies,21,56 it is smaller than many studies investigating risk factors for respiratory disease in cattle13,46 and may have contributed to the failure to identify a significant association between many of these factors and respiratory disease. Notwithstanding this, there were some interesting findings in our study relevant to respiratory disease control and detection on farms.
Our study identified that rates of vaccination against the pathogens associated with BRD were low and that almost one-third of animals with chronic lung lesions had not been given antimicrobials in the week prior to death, which suggests that clinical signs went undetected. It is also noteworthy that, of animals vaccinated against BoHV-1 and BRSV in this study, 7 and 5 different pathogens, other than those against which vaccination was performed, were detected, respectively. These findings highlight the need for both improved disease detection and earlier intervention as well as the need for further scrutiny of the criteria used by herd owners in deciding against which pathogens calves should be vaccinated. As data on the timing of vaccination were not collected, we acknowledge that close proximity of vaccination with a modified-live viral vaccine and PCR testing could contribute to positive results in some cases. It must also be stated that vaccines against P. multocida, H. somni, and M. bovis were not available for use in Ireland at the time of this study.
We acknowledge that factors such as the number of deaths experienced on farm, the cost of the service, and the distance of the farm from the laboratory could have exerted a selection bias on the carcasses submitted for postmortem examination in this study.47,50 Equally, it is acknowledged that the relative merits and target of specific methods of detection may introduce some bias into the results. The effect on bacterial culture of the dose, timing, and efficacy of antimicrobials that were administered to almost half of the study population within 7 d of death is impossible to predict. The use of PCR or IHC should circumvent this effect; however, bacterial pathogens in which detection was reliant on culture only (e.g., T. pyogenes) may be underrepresented. 6 Nevertheless, in our study, for the 4 bacterial pathogens typically implicated in BRD (H. somni, M. bovis, P. multocida, and M. haemolytica),30,55 both PCR (for all) and IHC (for all, except P. multocida) were employed in their detection.
The objectives of this research were to not only address deficits in our knowledge of, but also to compare, the pathogens, patterns of pneumonia, and associated epidemiologic risk factors of BRD in cohorts of affected and unaffected weaned cattle presented for postmortem examination. Cases of BRD in recently weaned calves were categorized into internationally accepted morphologic categories of pneumonia in an attempt to identify links between these subcategories and specific pathogens. Although a novel finding was the identification, more frequently than expected, of H. somni in all histologic categories of pneumonia, particularly fibrinosuppurative and caseonecrotic bronchopneumonia, the precise role of H. somni in contributing to specific diverse histologic patterns is difficult to discern. Significant associations between recognized animal and herd risk factors and BRD in bovine weanlings were not identified in this study. The breadth of respiratory pathogens detected in bovine lungs by various detection methods highlights the diagnostic value of parallel analyses in respiratory disease postmortem investigation. Our findings underline the scale and complexities of pulmonary inflammation associated with BRD and facilitate an evidence-based approach to devising methods of disease control.
Footnotes
Acknowledgements
We thank the staff in the 6 Irish RVLs: Athlone, Cork, Dublin, Kilkenny, Limerick, and Sligo, and in Bacteriology, Virology, and Pathology Divisions in the Central Veterinary Research Laboratory. The assistance of colleagues in the Agri-Food and Biosciences Institute, Northern Ireland with selected PCR analysis is also gratefully acknowledged. The authors also thank Dr. Jennifer McClure, Irish Cattle Breeders Federation, and Mr. Daniel Collins, Centre for Veterinary Epidemiology and Risk Analysis.
Authors’ contributions
GM Murray contributed to conception and design of the study; contributed to acquisition, analysis, and interpretation of data; drafted the manuscript; and agreed to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved. SJ More, D Sammin, RG O’Neill, MJ Casey, MC McElroy, WJ Byrne, and JP Cassidy contributed to conception and design of the study, and to acquisition, analysis, and interpretation of data. B Earley contributed to design of the study and to acquisition, analysis, and interpretation of data. TA Clegg contributed to design of the study and to analysis and interpretation of data. H Ball and CJ Bell contributed to acquisition and interpretation of data. All authors critically revised the manuscript and gave final approval.
a.
JumpStart Taq ReadyMix for quantitative PCR, Sigma-Aldrich Ireland, Arklow, Ireland.
b.
LSI VetMAX screening pack real-time PCR kit, Thermo Fisher Scientific, Waltham, MA.
c.
E.Z.N.A. tissue DNA kit, VWR International, Dublin, Ireland.
d.
MaximaProbe qPCR master mix, Thermo Scientific, Dublin, Ireland.
e.
Metabion International, Munich, Germany.
f.
MagNA Pure 96, Roche Life Sciences, Manheim, Germany.
g.
Stratagene Mx3005P, Agilent Technologies, Santa Clara, CA.
h.
QScript XLT one-step RT-PCR master-mix, VWR International, Radnor, PA.
i.
QIAamp DNA mini kit, Qiagen, Manchester, United Kingdom.
j.
QIAamp RNA mini kit, Qiagen, Manchester, United Kingdom.
k.
DNeasy blood and tissue kit, Qiagen, Manchester, United Kingdom.
l.
Stomacher lab blender, Seward Medical, London, United Kingdom.
m.
Sigma P5147-5G, Sigma-Aldrich, Arklow, Co. Wicklow, Ireland.
n.
UltraView universal DAB detection kit, Roche Diagnostics, Mannheim, Germany.
o.
DAKO X0909 serum free, Agilent Technologies, Santa Clara, CA.
p.
DAKO Envision kit (K5007), Agilent Technologies, Santa Clara, CA.
q.
DAB substrate kit SK-4100, Vector Laboratories, Burlingame, CA.
r.
Stata version 11, StataCorp, College Station, TX.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.