
Abstract
The aim of our study was to determine the association of Helicobacter spp. that had flexispira morphology with ovine abortion, and to understand the importance of these organisms as a cause of ovine abortion in New Zealand. A retrospective diagnostic survey was carried out on laboratory submissions from ovine abortion outbreaks. A comparison was made of the proportion of laboratory submissions where Helicobacter spp. were detected from flocks that had no other agent identified (group A) with a group that had a known cause of abortion identified (group B). This latter group was considered to be a negative control, given the premise that Helicobacter spp. were not causing abortions and that Helicobacter spp. should be present at a lower rate in the group. Where no diagnosis had been made, aborted material was positive for Helicobacter spp. with flexispira morphology in 8 submissions (20%, 8/40) from 5 of the 31 survey farms (16%, 5/31). Helicobacter spp. were not detected in any of the 18 submissions from the 17 control farms (group B). Helicobacter spp. were confirmed by 16S ribosomal RNA sequencing of 3 of the Helicobacter spp. isolated by culture from the livers of aborted sheep fetuses, and 7 of the 8 where samples were positive in a Helicobacter PCR assay. The Helicobacter spp. were identified as Helicobacter trogontum (Flexispira taxon 5 genotype) and Helicobacter bilis (Flexispira taxon 8 genotype). The findings support Helicobacter spp. being a probable causative agent of ovine abortions in New Zealand.
Introduction
New Zealand is free from the major causes of abortion in sheep, such as Chlamydophila abortus and Salmonella enterica subsp. enterica serovar Abortusovis, that cause considerable losses worldwide. 22 In New Zealand, the agents most commonly diagnosed in sheep abortion are campylobacters, toxoplasmas, and Salmonella enterica subsp. enterica serovar Brandenburg with these 3 agents reported to account for ~80% of sheep abortions where a causal agent is identified. 22 Other minor causes of infectious abortion that are diagnosed sporadically include Border disease virus (commonly known as hairy shaker disease virus), Listeria spp., Bacillus spp., Salmonella spp., Yersinia spp. and Brucella ovis. However where laboratory diagnosis is undertaken, the agents causing abortion are often not determined. 22 It is also possible that, for many outbreaks of sheep abortion, a veterinary diagnostic investigation is not performed.
In the early 1990s, a large outbreak of abortions in mature ewes in Canterbury in the central-eastern South Island of New Zealand was extensively investigated. 13 In this outbreak, 10% of 2,950 ewes aborted over a 50-day period. These ewes had been previously vaccinated against campylobacteriosis and toxoplasmosis. Gross and histologic examination of the liver and placentas from many of the aborted lambs showed lesions that were identical to those seen in campylobacter abortion outbreaks but cultures for this bacterium and others were negative. However, weakly staining, Gram-negative, beaded, tapered curved bacilli were identified in the stomach contents of many of the aborted lambs. The study 13 suggested that this bacterium may have been Fusobacterium nucleatum, which had been reported in similar but small abortion outbreaks in ewes in North America. 17
Since this abortion outbreak in the early 1990s, similar, occasionally large outbreaks of last trimester abortions, stillbirths, and the birth of weak nonviable lambs have been reported. These occurred annually in most cases, in vaccinated, intensive ewe flocks in the regions of Canterbury, Otago, and Southland in the South Island of New Zealand. Aborted fetuses were characterized by liver lesions that were grossly and microscopically identical to campylobacter infection but with no evidence of common etiological agents identified. Identification of the agent has been pursued, but, to date, only presumptive characterization of a bacterial agent has been made 13 (Smart JA, Gill JM. Fusobacterium like abortions—an overview. Proceedings of the Society of Sheep and Beef Cattle Veterinarians of the New Zealand Veterinary Association; 1999).
In 2009, as part of a Ministry for Primary Industries (MPI) investigation, 5 lambs from a large abortion outbreak in Southland underwent postmortem examination. In 2 lambs, histology identified large areas of coagulative necrosis, with silver staining showing faintly curved bacilli clustered in the canaliculi of intact hepatocytes, along the edge of the necrotic liver lesions. Electron microscopy of a small section of formalin-fixed liver from a paraffin block revealed large numbers of fusiform-shaped bacteria with periplasmic fibers in the biliary canaliculi (Bingham P. Quarterly report of investigations of suspected exotic diseases. Surveillance 2010;37:35–36). 9 The ultrastructure of these bacteria resemble bacteria previously identified in aborted lambs in North America as nonclassified, microaerophilic bacteria with spiral morphology, periplasmic fibers and bipolar tufts of sheathed flagella designated “Flexispira rappini.”14–16
Subsequent studies indicated that organisms with this ultrastructure were members of the genus Helicobacter and represented at least 10 different taxa.3,18 These taxa were given the designations “F. rappini” taxon 1 through taxon 10. In phylogenetic trees based on 16S ribosomal RNA (rRNA) sequences, the “F. rappini” taxa are scattered among other Helicobacter species, but a study using more information-rich 23S rRNA sequencing found that all bacteria with “F. rappini” morphology fall into a single monophyletic group within the Helicobacter genus. 4 “F. rappini” taxa 2, 3, 8 and 9 have been proposed as Helicobacter bilis, and “F. rappini” taxa 1, 4, 5, and 6 as Helicobacter trogontum.10,11 Individual taxa have been associated with animal species sources. Although combining “Flexispira” taxa into named Helicobacter species was useful for simplifying Helicobacter taxonomy, it obscures the fact that 16S rRNA sequences for different “Flexispira” taxa are distinguishable and reflect animal origin. For example, “F. rappini” taxa 2, 4, and 5 have been previously isolated from sheep, “F. rappini” taxon 3 from pigs, 3 “Flexispira” taxon 8 from dogs and humans, 3 and “Flexispira” taxon 6 from rats. 3 It thus appears useful to note if a H. trogontum isolate is similar to pig (taxon 1), sheep (taxon 4 or 5), or rat (taxon 6) genotypes.
Since the earlier detection of the organism with “F. rappini” morphology in aborted New Zealand sheep, 9 methods for their detection and classification have improved. The aim of our study was to determine if there was an association of Helicobacter spp. that had flexispira morphology with sheep abortions, to determine Helicobacter spp. present in various sheep flocks, and to ascribe their importance as a cause of ovine abortion in New Zealand.
Materials and methods
Survey design
A retrospective survey was undertaken to compare the proportion of routine regional laboratory submissions where Helicobacter spp. were detected from flocks that had no agent of abortion identified (group A; 31 farms, 40 submissions) compared to a group that had a known cause of abortion (group B; 17 farms, 18 submissions). This latter group (group B) was considered to be a negative control group given the premise that Helicobacter spp. were not causing abortion and that this agent should be present at a lower rate in this group. Group B consisted of submissions actively collected from farms experiencing perinatal mortality resulting from a non-infectious etiology such as inclement weather and mismothering, and from routine abortion submissions where a definitive diagnosis had been made of an agent normally associated with ovine abortion (Campylobacter spp., Toxoplasma spp., Salmonella spp., and Listeria spp.). The study was carried out in sheep flocks experiencing abortions during the spring of 2012 in the southern South Island of New Zealand.
Routine abortion testing
In general, routine laboratory tests were used to determine the presence of endemic agents (Campylobacter spp., Listeria spp., Salmonella Brandenburg, Toxoplasma gondii, and Border disease virus) in all submissions included in the survey. a A minimum set of data was collected for each abortion case. The data included age, class of sheep affected, start and end date, number and percent aborting, and ewe vaccination history.
Aborted lambs were autopsied. a If liver lesions were identified, the lesions were fixed in 10% neutral buffered formalin, embedded in paraffin by conventional methods, routinely sectioned at 4–6 μm, and stained with hematoxylin and eosin and a silver stain (Warthin–Starry). In a small number of these cases, sections of lung, brain, and very occasionally placenta were also similarly fixed, processed, and examined.
Heart blood taken at autopsy was tested for antibodies to T. gondii by a latex agglutination test. b Stomach contents from the aborted lambs were cultured on campylobacter isolation blood free agar c and identified using methodology from a previous study. 19 Stomach contents were also cultured aerobically on blood agar and incubated for 24 h at 42°C and, from selected lambs, also plated onto Rambach medium d and also incubated for 24 h at 42°C. Suspect Salmonella spp. isolates were then plated onto Colombia sheep blood agar and MacConkey agar, c and colonies were subject to an antisera agglutination test. e Any presumptive Salmonella spp. isolates were placed onto nutrient agar slopes and sent to the Institute of Environmental Science and Research, Enteric and Leptospira Reference Laboratory f for serotyping.
Fetal liver samples were collected from 58 aborted fetuses into brain–heart infusion broth (BHI) with 20% glycerol and stored at −20°C. The samples were transported on dry ice to 2 different laboratories, the Animal Health Laboratory (AHL), Ministry for Primary Industries (Wallaceville, Wellington, New Zealand) and the Division of Comparative Medicine, Massachusetts Institute of Technology (MIT; Cambridge, Massachusetts), for parallel testing. Helicobacter spp. culture and PCR testing were conducted at MIT, whereas only PCR was performed at AHL.
Helicobacter culture
Tissue samples, measuring ~5 mm × 5 mm × 5 mm, were cut into small pieces in freeze media, homogenized with tissue grinders, and 100 µL of the supernatant placed onto CVA plates (with cefoperazone, vancomycin, and amphotericin B) g ; another 500 µL of the supernatant was passed through a 0.65-µm syringe filter onto a blood agar plate with 5% sheep blood. g The plates were incubated at 37°C microaerobically in a vented jar filled with mixed gas containing 80% N2, 10% H2, and 10% CO2. After 2–5 days of incubation, the primary isolates with visible colonies or film-like growth were Gram stained and plated on blood agar plates for further analysis.
DNA extraction
Genomic DNA from fetal liver of sheep and bacterial isolates was extracted using a commercial kit h following the manufacturer’s protocol at MIT. At AHL, each fetal liver sample was removed from the BHI with 20% glycerol and dabbed dry before removing ~20 mg of tissue from the center of the sample. Genomic DNA was extracted on a commercial instrument i using DX reagent pack and DX tissue digest reagent following the manufacturer’s protocol, with minor modifications: the tissue was digested with 420 µL of DX tissue digest (containing 1% v/v DX digest enzyme) and incubated overnight in a heat block at 56°C. The digested tissue was then centrifuged at 2,500 × g for 5 min, 220 µL of particulate-free supernatant was transferred to a lysis block, and the extraction was completed on the instrument, i with a final elution volume of 200 µL.
PCR amplification
Genomic DNA from liver samples of aborted fetuses was examined by PCR as detailed below. At AHL, Helicobacter genus–specific PCR using primer pairs C97 (276-291 forward) and C98 (681-698 reverse) or C97 and C05 (1478-1495 reverse) were used to generate 16S rRNA amplicons of ~400 or 1,200 base pairs, respectively, as described 7 with modifications: 2.5 µL of DNA was added to a reaction mixture containing a commercial master mix j supplemented with 25 mM MgCl2, to a final concentration of 2.75 mM, and 0.5 µM of each primer to achieve a total volume of 25 µL. The following conditions were used for amplification on a thermal cycler k : initial denaturation at 95°C for 2 min, then 30 cycles of denaturation for 15 s at 95°C, annealing for 15 s at 60°C, and extension for 1 min at 72°C.
At MIT, C97 and C05 primers were used, and the PCR amplifications were performed with a thermal cycler and a commercial mastermix. h The following conditions were used for amplification: 35 cycles of denaturation at 94°C for 1 min, annealing at 58°C for 1.5 min, and elongation at 72°C for 2 min followed by an elongation step of 7 min at 72°C. 7 All PCR products were visualized by electrophoresis on a 1.5% (w/v) ultrapure agarose gel l using a commercial stain. m Campylobacter genus–specific primers C98 and C99 were used to confirm isolates as Campylobacter spp. 21 For the bacterial isolates cultured at MIT, the nearly complete 16S rRNA gene was amplified using primers F24 (9-27 forward) and F25 (1524-1541 reverse), 12 using the same PCR conditions employed for the C97 and C05 primers.
16S rRNA sequence analysis
16S rRNA sequencing was performed to verify that the amplicons were Helicobacter 16S rRNA and to determine the Helicobacter species detected. At AHL, 7 PCR-positive 1.2 kilobase (kb) amplicons from the C97 and C05 primer pair were purified. n Purified DNA was subjected to automatic dye-terminator cycle sequencing with a commercial kit and on a sequencer k or sequenced fully on both strands by a commercial provider o as previously described. 21 Alignments were performed on the trimmed sequences, p and sequences were compared directly with the NCBI GenBank nucleotide database by BLAST search (http://blast.ncbi.nlm.nih.gov/Blast.cgi). At MIT, 1.2-kb PCR products from 5 of the Helicobacter genus PCR assays were sequenced. For the 3 bacterial isolates from livers, the nearly complete 16S rRNA genes were sequenced. Sequences were entered into an aligned database of Campylobacter and Helicobacter species sequences and phylogenetically positioned within the Helicobacter species determined by constructing a neighbor-joining tree. 5
Electron microscopy
Isolates identified as Helicobacter spp. with flexispira morphology were examined by electron microscopy. Cells grown on blood agar for 48 h were gently suspended in 10 mM Tris–HCl buffer (pH 7.4) at a concentration of ~108 cells/mL. Samples were negatively stained with 1% (w/v) phosphotungstic acid (pH 6.5) for 20–30 s. Specimens were examined with a transmission electron microscope q operating at 100 kV.
Statistical analysis
Detection of Helicobacter spp. as a function of abortion submissions tested was compared between group A and B (negative controls) using the chi-square test for equality of proportions in R v2.15 (https://CRAN.R-project.org).
Results
The survey included 58 submissions of aborted material received from 48 farms surveyed from 30 July 2012 to 25 October 2012. Data from these submissions showed that the average abortion rate for the 5 farms with submissions positive for Helicobacter spp. was 10.8% (median: 7.5%, range: 3.2–25%), whereas for farms in group B, the average abortion rate for known causes of abortion was 2.8% (median 2.5%; range 1.1–5.4%; note—abortion prevalence data for group B submissions was limited to 8 farms). However, this data related to the time of submission, and it is possible that subsequently more animals aborted from these outbreaks. No indication was given as to whether the population at risk used to calculate the prevalence affected was the mob where abortions were occurring or the total number of breeding animals on the property.
Gross and histology findings
Most aborted lambs had minimal postmortem decomposition. In almost all submissions, the placentas were not available. In lambs later confirmed as Helicobacter spp. positive by PCR, 5 of 8 submissions exhibited gross liver lesions. In 4, autopsy findings included a swollen liver containing a few to moderate numbers of round, distinct pale foci often with depressed centers, creating a ring-like appearance. These foci varied in size from 1–2 to 10 mm in diameter (Fig. 1). In 1 submission, the livers from the aborted lambs were a normal color but were enlarged and appeared more autolyzed in contrast to the rest of the carcass. These livers contained a number of embedded soft spherical 20–30 mm in diameter nodules.
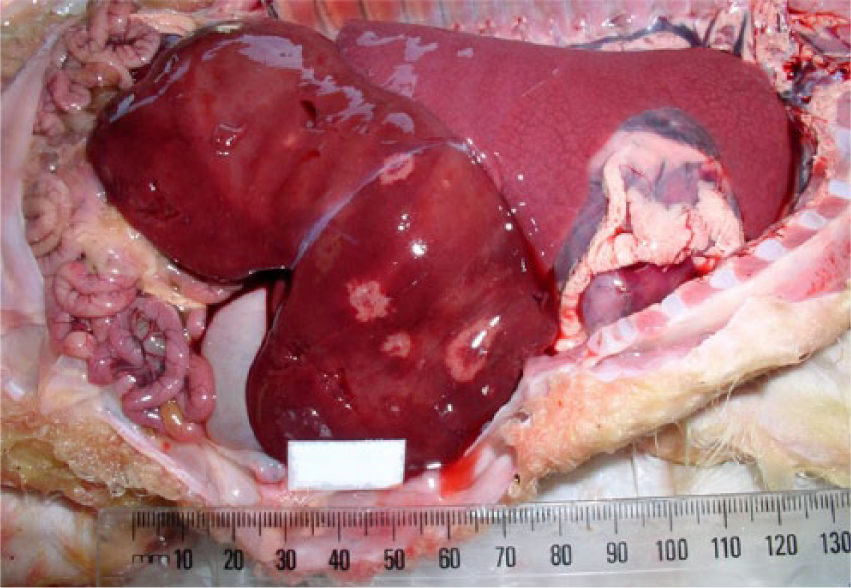
Viscera of aborted near full-term lamb from farm 23 showing minimal postmortem decomposition and aerated lungs. Liver swollen with a small number of variably sized, circular, pale foci, many with depressed centers.
The livers of most Helicobacter spp. PCR–positive lambs showed random, small focal to locally extensive areas of acute coagulative necrosis (Figs. 2, 3). In 1 Helicobacter spp. PCR–positive submission, there were no histologic changes present in the liver. In the larger lesions, the necrosis was periacinar to midzonal or affected the entire lobule. Neutrophils were in low numbers in the necrotic areas and, in some necrotic areas, the central veins were thrombosed. In some affected livers, there was quite extensive portal infiltrate of mononuclear cells often extending into the parenchyma. Silver staining showed variable numbers of slightly curved fusiform bacilli in 3 affected livers, clustered linearly in the bile canaliculi of the surviving hepatocytes bordering the necrotic areas (Fig. 4).
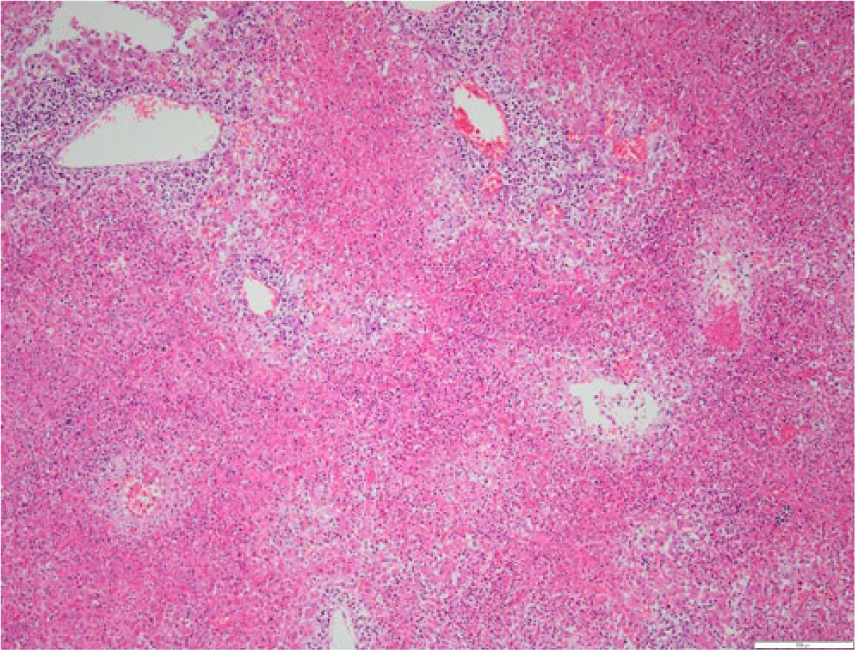
Section of liver from lamb in Figure 1. There are large confluent areas of necrosis involving whole lobules. The surrounding portal veins have an extensive mononuclear cell infiltrate. Hematoxylin and eosin. 10×. Bar = 200 μm.
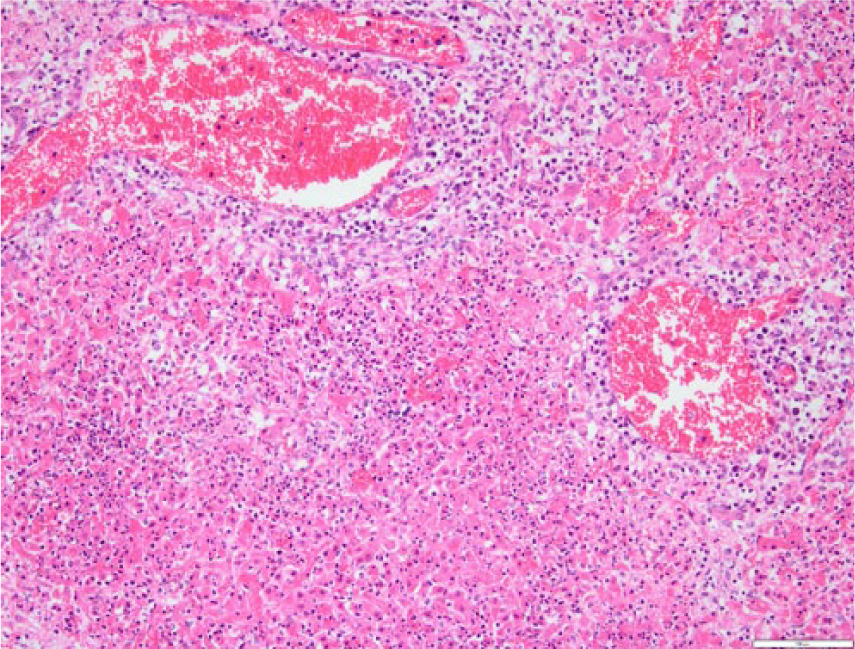
Section of liver from lamb in Figure 1. Neutrophils are infiltrating extensive areas of necrotic hepatocytes. Mononuclear cells (most likely small lymphocytes) are infiltrating in large numbers around the dilated portal veins. Hematoxylin and eosin. 20×. Bar = 100 μm.
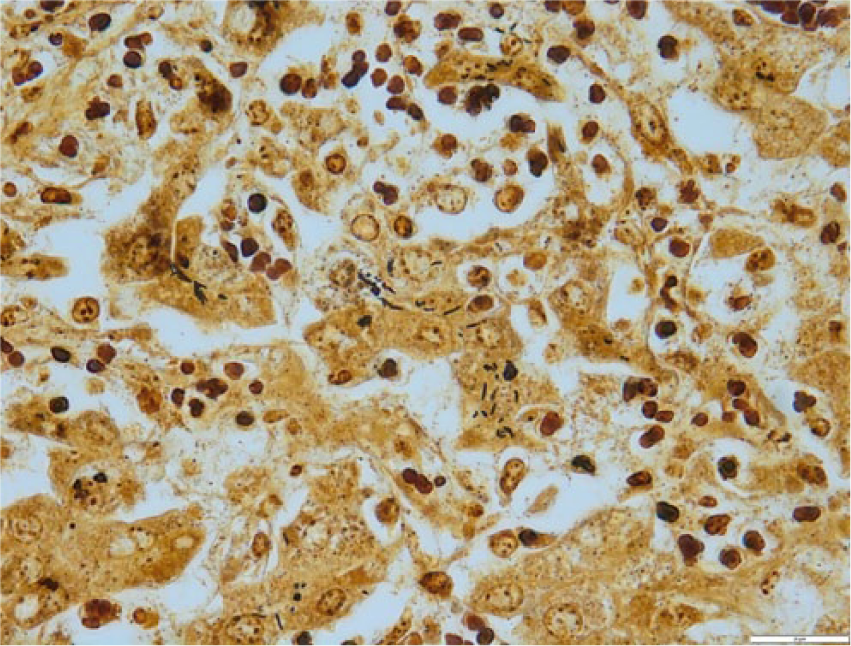
Section of liver from lamb in Figure 1. Dark-staining, faintly curved bacilli resembling Helicobacter in the cytoplasm of intact hepatocytes. The bacilli are occasionally arranged in linear clusters in canaliculi. Warthin–Starry. 100×. Bar = 20 μm.
Helicobacter culture and electron microscopy
Helicobacter spp. were isolated from the liver samples of 3 aborted fetuses (Table 1). By 16S rRNA sequence analysis, 2 isolates had 98.9% identity with H. trogontum (“Flexispira” taxon 5 genotype), and 1 isolate had 99.7% identity with H. bilis (“Flexispira” taxon 8 genotype). The position of these isolates can be seen in Figure 5, which depicts the neighbor-joining tree for the enterohepatic Helicobacter spp. The Helicobacter spp. isolates had typical “F. rappini” morphology with fusiform shape (0.5 × 4–5 μm), periplasmic fibers, and multiple bipolar sheathed flagella (Fig. 6).
Summary of Helicobacter spp. and other abortion agents detected from group A (no abortion agent identified) and group B (control group) submissions of aborted material.

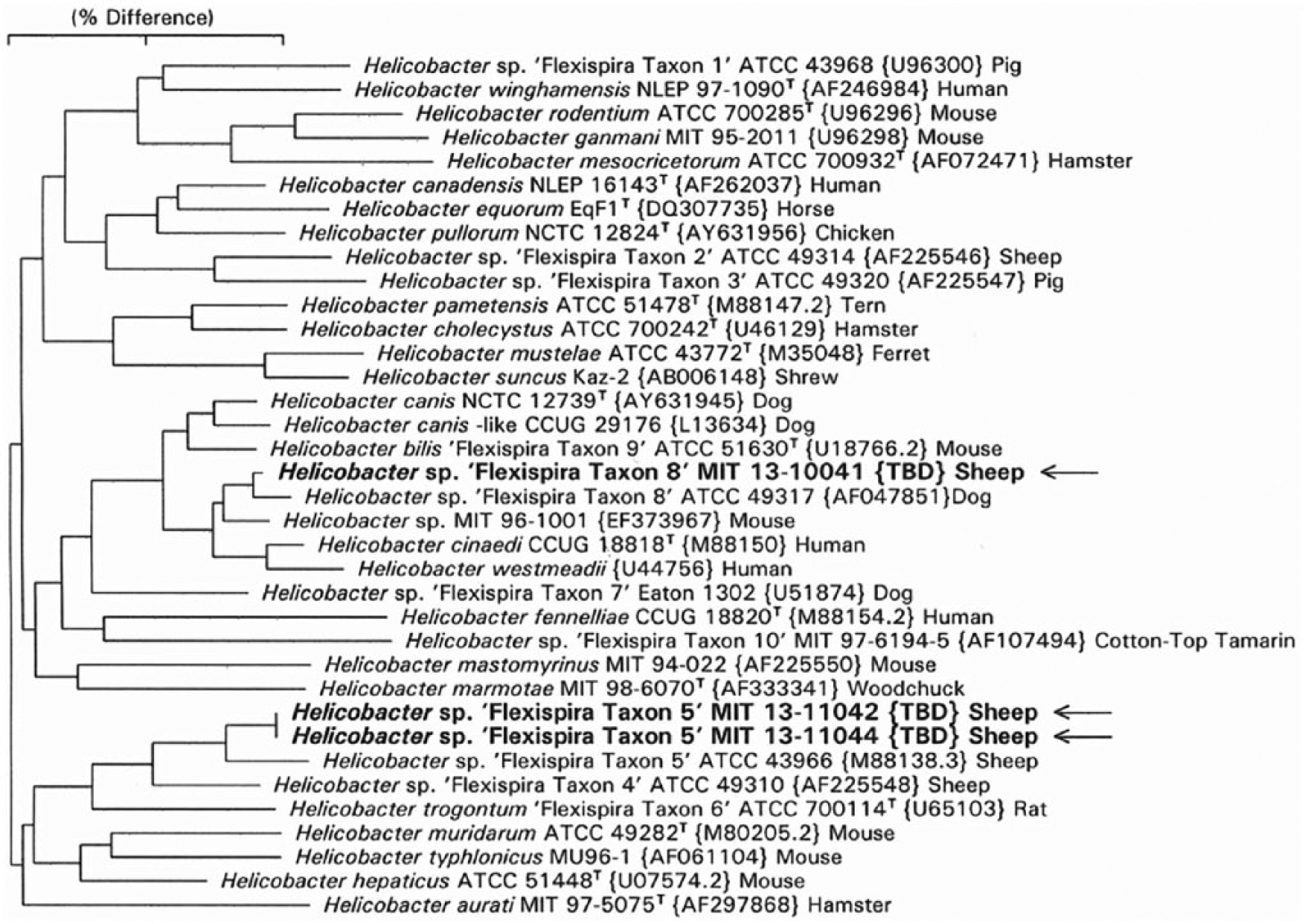
Phylogenetic tree for enterohepatic helicobacters, including those with flexispira morphology associated with abortion in sheep. The neighbor-joining tree is based on comparison of 1,466 base positions. GenBank accession numbers are given in brackets. The scale bar represents a 2% sequence divergence. “Flexispira” taxa are numbered as in Dewhirst et al. 4
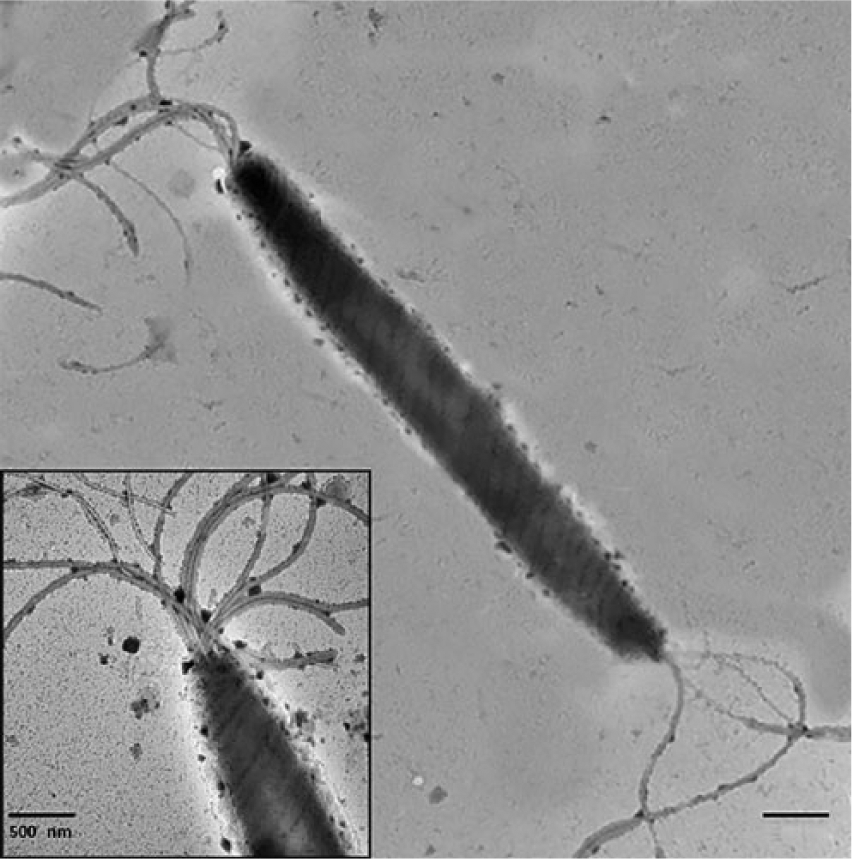
Negatively stained preparations of Helicobacter spp. with flexispira morphology. Typical cell morphology with tufts of bipolar, sheathed flagella. Bar = 0.5 nm. Inset: higher magnification of end of cell illustrating tuft of sheathed flagella. Bar = 0.5 nm.
PCR amplification and sequence analysis
Helicobacter DNA was identified in 8 of 58 (14%) fetal liver samples by the Helicobacter genus–specific PCR with 1.2-kb primers (Tables 1, 2). The 16S rRNA sequences of the PCR products reveal that 3 of the sequences had the same identity as the 3 Helicobacter spp. isolated by microaerobic culture (above). Sequences from 4 additional PCR products had 99% identity with H. trogontum (“Flexispira” taxon 5 genotype). One weak PCR-positive sample failed sequencing. This sample was negative in the Helicobacter genus–specific PCR at AHL. The position of these PCR-positive samples can be seen in Figure 5, which depicts the neighbor-joining tree for the enterohepatic Helicobacter spp. All 8 Helicobacter spp.–positive samples were negative for Campylobacter spp. by PCR.
Farm details and laboratory test results from the 8 submissions of aborted material from 5 farms in New Zealand experiencing ovine abortion (samples positive by PCR for Helicobacter spp.).*
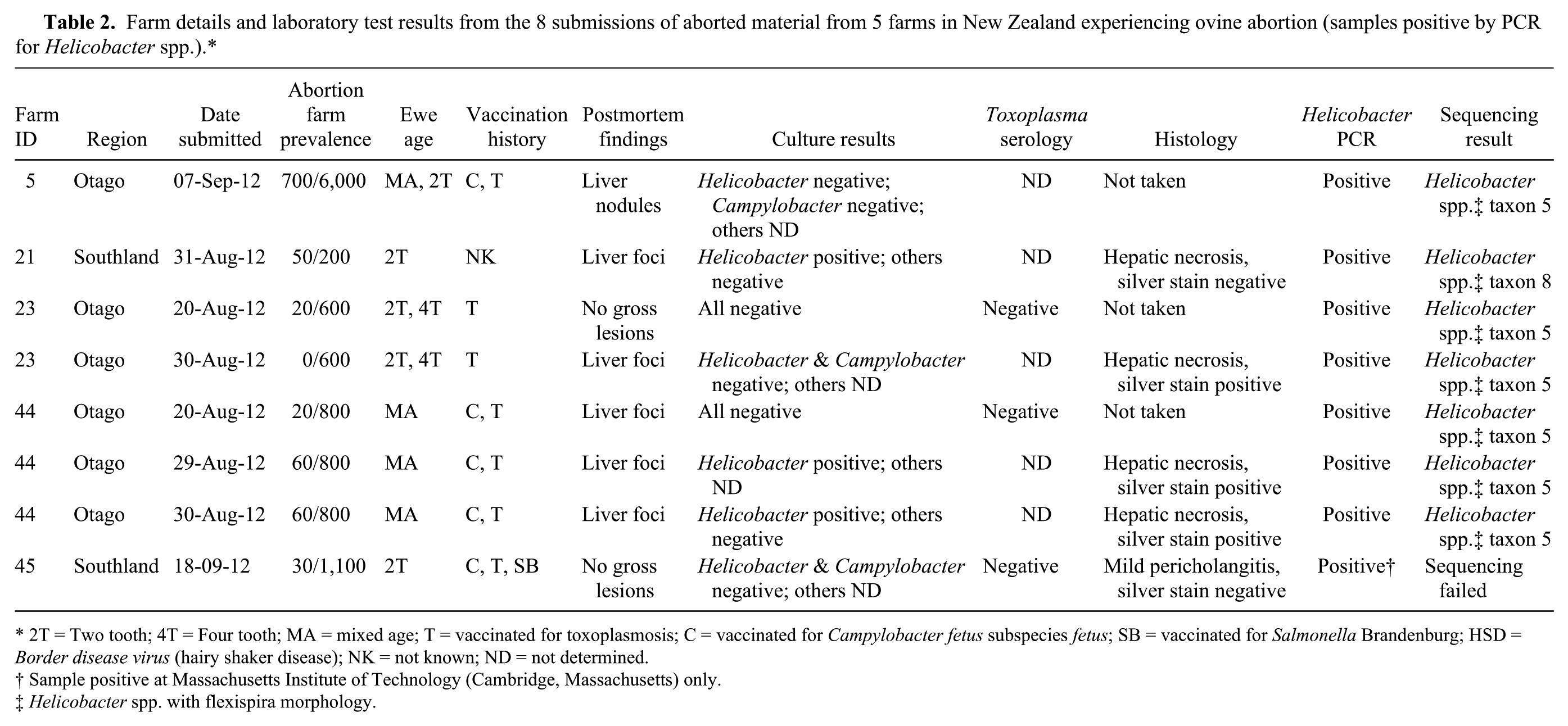
2T = Two tooth; 4T = Four tooth; MA = mixed age; T = vaccinated for toxoplasmosis; C = vaccinated for Campylobacter fetus subspecies fetus; SB = vaccinated for Salmonella Brandenburg; HSD = Border disease virus (hairy shaker disease); NK = not known; ND = not determined.
Sample positive at Massachusetts Institute of Technology (Cambridge, Massachusetts) only.
Helicobacter spp. with flexispira morphology.
From group A, aborted material was positive for Helicobacter spp. in 8 of 40 (20%) submissions received from 31 farms. From group B, the control farms, Helicobacter spp. was not detected in any of the 18 submissions from 17 farms (Tables 1, 2). There was no statistical difference between the proportions of Helicobacter spp. detected in laboratory submissions from the 2 groups (P = 0.10). A sample size of 53 submissions from each group would have been necessary to detect a difference of 18% between groups where the power is set at 90%.
The 58 submissions subjected to laboratory analysis for Helicobacter spp. represented 41% (58/142) of all submissions for ovine abortion made to 1 regional veterinary laboratory a over the 2012 lambing period. From all submissions made to the laboratory for this period, a definitive diagnosis was reached for 38% (54/142) of cases. The breakdown of confirmed diagnoses being 38 of 142 (27%) Salmonella Brandenburg, 6 of 142 (4%) Campylobacter fetus subspecies fetus, 5 of 142 (4%) Toxoplasma spp., 2 of 142 (1%) Listeria spp., 2 of 142 (1%) Border disease virus, and 1 of 142 (<1%) C. fetus subspecies jejuni. The breakdown of confirmed diagnoses for 2009–2013 is summarized in Table 3.
Breakdown of confirmed diagnoses of sheep abortion for submission to Gribbles Veterinary Pathology, Invermay, New Zealand for 2009–2013.
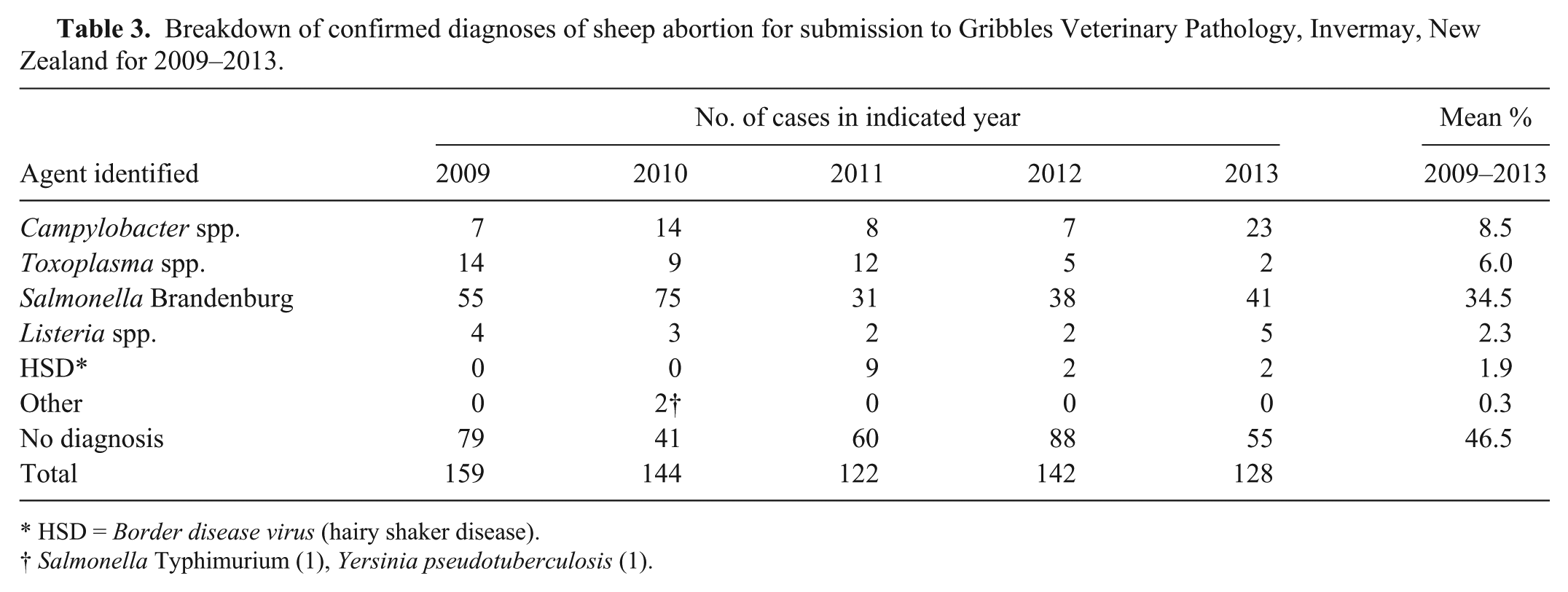
HSD = Border disease virus (hairy shaker disease).
Salmonella Typhimurium (1), Yersinia pseudotuberculosis (1).
Discussion
A Helicobacter spp. with “F. rappini” morphology was first isolated in 1985 from twin, late-term aborted sheep fetuses. 16 Thereafter, inoculation experiments in pregnant ewes and guinea pigs indicated the biological plausibility of “F. rappini” as a cause of abortion.1,15 These experimentally infected animals had similar histologic pathology to that described in the initial abortions. Since its first detection, Helicobacter spp. have occasionally been implicated as a sporadic cause of abortion in sheep from the United States, the United Kingdom, and New Zealand (Bingham P, 2010).2,9,14
The study reported herein has confirmed the presence of Helicobacter spp. on multiple farms experiencing abortion in 2012 with histology consistent with previous cases. Helicobacter spp. were detected in a relative high percentage of abortion submissions where no diagnosis had been determined (16%, 5/31). On 3 of the farms where Helicobacter spp. were detected, there were only single detections relating in each case to a single submission of abortion material. On the other 2 farms, Helicobacter spp. were identified from multiple submissions (3 of 4 submissions on one farm and from both submissions from the other). On both of these farms, lambs positive for Helicobacter spp. had been detected over a 10-day period. Helicobacter spp. were not detected in the 18 negative control submissions (group B). Based on this preliminary data, we consider that if Helicobacter spp. was a true pathogen, detection would be unlikely to occur in this group, as sheep had aborted from either environmental factors or to a specific pathogen known to cause abortion.
As additional isolates of Helicobacter spp. were identified, a 16S rRNA analysis of 36 strains of Helicobacter spp. from multiple hosts was undertaken. 3 The investigators divided these strains into 10 taxa. “F. rappini” taxa 2, 4, and 5 were isolated from aborted sheep fetuses or sheep (tissue not designated). Subsequently, using a polyphasic taxonomic approach, taxa 2, 3, and 8 were grouped with taxon 9 as H. bilis and taxa 1, 4, and 5 were grouped with taxon 6 as H. trogontum.10,11 The taxonomic classification and identity of taxa 5 and 8 strains identified in our study is consistent with our current findings of H. trogontum from aborted sheep fetuses; however, H. bilis (“Flexispira” taxon 8) was not previously isolated from sheep, but from human and dog feces, 3 as well as mouse feces and liver. 7
In 2014, H. canis (a helicobacter with curved morphology) was cultured from sheep feces obtained from a small flock with a history of exposure to cats and dogs, suggesting that helicobacters can be transmitted from dogs to sheep. 20 Given that H. bilis (“Flexispira” taxon 8) has also been isolated from dog feces, the close association of dogs and sheep could explain cross-species transmission of H. bilis.
The pathogenesis of abortion caused by Helicobacter spp. is likely to mirror the mechanisms of infection of the other enterohepatic Helicobacter species.8,9 It is probable that in the ewe, bacterial transfer via the portal system results in hepatitis, which may be chronic and active in nature, seeding an ongoing bacteremia.5,6,8 In guinea pigs inoculated intraperitoneally, Helicobacter spp. were cultured from heart blood at autopsy 11 days after inoculation, suggesting persistent bacteremia. 1 As a result of the bacteremia, suppurative placentitis and a breakdown of the placental-blood barrier allows Helicobacter spp. to enter the fetal portal system directly, or via the fetal fluids and gut. 15 Regular isolation or visualization of Helicobacter-like organisms in the stomach contents of aborted fetuses supports the latter route. 15 In the fetus, necropurulent hepatitis ensues characterized by focal areas of periportal and sinusoidal necrosis, consistent with that seen in campylobacter abortions. 14 The large, circumscribed liver lesions seen in our series of Helicobacter spp.–associated abortions are reportedly more common than similar liver lesions noted with Campylobacter spp. abortions. 14 In a trial involving 28 ewes inoculated with Helicobacter spp. intravenously, 4 ewes aborted or gave birth to weak infected lambs, indicating that other factors, possibly previous exposure or impaired immunity in the ewe, may be involved in inducing Helicobacter spp. abortion. 15
There have been relatively few reports where Helicobacter spp. with flexispira morphology have been implicated in sheep abortion.2,14,16 Of 1,784 ovine abortions and stillbirths assessed over a 10-year period by a veterinary laboratory in South Dakota, Helicobacter spp. with flexispira morphology was detected in 10 (0.56%) submissions. 14 In New Zealand, an abortion episode investigated by the MPI was provisionally diagnosed as being caused by Helicobacter spp. (Bingham P, 2010). 9 Abortions were characterized by late-term abortions from 100 days postmating, variably affecting 9–20% of ewes across various management groups. In the current series of abortions covering the 2012 lambing season, the prevalence of abortion in the flocks from which Helicobacter spp. were isolated was similar to the above case, and comparable to that reported for the other causes of abortion during the 2012 season.
There is limited data on the national incidence of abortion in sheep flocks in New Zealand. It has been estimated that abortion outbreaks of varying intra-farm prevalence occur in 1–2% of flocks annually; however, small scale outbreaks are not necessarily observed or reported. In many of the outbreaks that are investigated, a causal agent is not identified. 23 For example, a previous study 23 reported that 18–49% of abortion storms per year did not have an agent identified for the period of 1973–1996. Data for 2009–2013 showed similar results with an agent not identified in 35–61% of cases where specimens were submitted (Table 3).
The numbers of outbreaks investigated and the number of submissions made per outbreak from our survey are insufficient to provide a clear indication of relative impact of infectious causes of abortion in the region. However, assuming that Helicobacter spp. were detected at the same rate in the other abortion submissions where no cause was identified, Helicobacter spp. would have accounted for 13% (18/142) of abortion diagnoses at this laboratory (Table 3). We have limited knowledge on the epidemiology of Helicobacter spp.; therefore, it is possible that the methods we have employed may be under-representing true Helicobacter spp. prevalence. Further studies are needed to understand the full impact of H. bilis and H. trogontum as causes of ovine abortion; however, the preliminary data we present indicates that Helicobacter spp. are an important cause of sheep abortions in the southern regions of New Zealand.
It is possible that there are a number of other infectious causes of ovine abortion that have not been identified because they are not part of routine abortion diagnostic panels. Helicobacter spp. may represent a proportion of these undiagnosed cases, but there are likely to be other infectious causes that have not been elucidated. The cause of abortion is multi-factorial, and it may be challenging to clarify the involvement of some of the less common infectious causes of abortion from environmental and host factors. Advances in molecular technology may assist with identification of infectious agents previously undetected.
Previous New Zealand investigations identified silver-staining bacteria, at the edges of diseased liver areas, which were confirmed using electron microscopy as having morphology consistent with Helicobacter spp. (Bingham P, 2010). 9 The presence, in the current study, of H. bilis and H. trogontum in aborted lambs with consistent histology, and its absence in lambs from which a known abortion agent was confirmed, add weight to the earlier New Zealand findings, and provide evidence that Helicobacter spp. are unlikely to be an incidental finding in aborted fetuses. Our findings support a conclusion that Helicobacter spp. with flexispira morphology are probable causative agents of ovine abortions in New Zealand.
Conducting a cross-sectional, large-scale survey across different sheep-rearing regions of New Zealand, along with testing a larger number of negative control samples, will be necessary to understand the true impact of Helicobacter spp.–associated abortions. A further objective will be to determine whether Helicobacter spp.–induced abortions present an emerging condition and whether research into prophylaxis or treatment is warranted. Improved diagnostics implemented in New Zealand for this study, with the assistance of overseas collaborators, will be instrumental in helping achieve these objectives.
Footnotes
Acknowledgements
We thank laboratory staff and other members of the Incursion Investigation team at the Investigation and Diagnostic Centre, Ministry for Primary Industries for their contribution to this investigation. We acknowledge the expert assistance of Robert Fairley with reviewing the histology, and of Kelly Buckle with taking the photomicrographs included in this publication.
Authors’ contributions
J Gill contributed to acquisition, analysis, and interpretation of data; drafted the manuscript; and gave final approval. TG Haydon and BJ Paster contributed to acquisition, analysis, and interpretation of data. TG Rawdon, JG Fox, and RP Spence contributed to conception and design of the study; contributed to acquisition, analysis, and interpretation of data; drafted the manuscript; critically revised the manuscript; and gave final approval. AMJ McFadden, HJ Ha, Z Shen, Y Feng, J Pang, and AG Swennes contributed to analysis and interpretation of data. FE Dewhirst contributed to acquisition, analysis, and interpretation of data; critically revised the manuscript; and gave final approval. TG Rawdon, FE Dewhirst, JG Fox, RP Spence, and agreed to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
a.
Gribbles Veterinary Pathology, Invermay, New Zealand.
b.
Eiken latex test, Eiken Chemical Co. Ltd, Tokyo, Japan.
c.
Fort Richard Laboratories, Otahuhu, New Zealand.
d.
CHROMagar, Paris, France.
e.
Sifin Diagnostics GmbH, Berlin, Germany.
f.
Institute of Environmental Science and Research, Enteric and Leptospira Reference Laboratory, Wallaceville, Wellington, New Zealand.
g.
Remel Inc., Lenexa, KS.
h.
Roche Molecular Biochemicals, Indianapolis, IN.
i.
Qiagen, Limburg, The Netherlands.
j.
Kapa Biosystems, Boston, MA.
k.
Applied Biosystems Inc., Foster City, CA.
l.
Invitrogen Corp., Carlsbad, CA.
m.
Biotium Inc., Hayward, CA.
n.
Bioline Ltd, London, UK.
o.
Macrogen Corp., Cambridge, MA.
p.
Genious Pro 5.5.6, Biomatters Ltd, Auckland, New Zealand.
q.
JEM-1200EX, JEOL Ltd., Tokyo, Japan.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was internally funded by the Ministry for Primary Industries, Investigation and Diagnostic Centre, and supported by Office of The Director of the National Institutes of Health under award number T32-OD010978 (JGF) and R01-OD011141 (JGF), National Institute of Environmental Health Sciences under award P30-ES002109 (JGF), and The National Institute of Dental and Craniofacial Research under award R37DE016937 (FED). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.