
Abstract
Diagnostic analyses often employ single antibody systems but are potentially limited by epitope sequence variation. United States regulatory testing for scrapie primarily uses antibody F99/97.6.1 for immunohistochemistry (IHC) of the prion protein associated with scrapie (PrPSc). Whereas the epitope bound by F99/97.6.1 is highly conserved in sheep, a polymorphism in caprine PRNP results in a glutamine to lysine change at codon 222 and affects PrP detection. This study evaluated the performance of immunoassays (Western blot and IHC) in the presence of PRNP polymorphisms observed in U.S. goat populations. Effects of naturally occurring caprine prion protein alterations at codons 142, 143, 146, 154, or 222 were first evaluated using bacterially expressed recombinant normal cellular prion protein (rec-PrPC) and commercially available antibodies (F99/97.6.1, F89/160.1.5, L42, and SAF84). Detection of rec-PrPC using F89/160.1.5 was reduced by alterations at 142 and 143; this was also observed in brain PrPC from goats expressing these PRNP variants. Effect of allelic variation at 222 was confirmed by Western blot with F99/97.6.1. No differences were observed with L42 or SAF84. IHC of brain demonstrated reduced signal with F89/160.1.5 in animals heterozygous at 143. Decreasing F89/160.1.5 titers were used to demonstrate the impact of PrPSc immunolabeling in preclinical goats and as a surrogate for F99/97.6.1 detection in 222 variants. In the absence of epitope-relevant knowledge of individual goat PRNP, a multi-antibody approach or an antibody that binds an invariant site may provide a more robust immunoassay of PrPSc in classical scrapie, thus reducing the likelihood of false-negative results due to allelic variation.
Introduction
Scrapie is a transmissible spongiform encephalopathy of sheep and goats. The infectious agent is referred to as a prion, an essential component of which is disease-associated, misfolded isoforms of the prion protein associated with scrapie (PrPSc). 31 Immunoassays are the mainstay of current testing and are designed to detect PrPSc in a variety of tissues. Antemortem and postmortem detection of disease has primarily been through immunohistochemical (IHC) analyses of the brain and peripheral lymphoid tissues.3,15,19 PrPSc-specific enzyme-linked immunosorbent assay and observations of resistance to proteinase K digestion via conventional Western blot are also commonly used. 12 Classical scrapie is slowly progressive in sheep 35 and goats, 36 and development of antemortem tests has aided eradication efforts through detection of preclinical animals. Such tests include IHC analysis of the third-eyelid lymphoid tissue in sheep 25 and analysis of rectoanal mucosa–associated lymphoid tissue in sheep 15 and goats. 16 In the United States, both antemortem and postmortem IHC testing uses a single monoclonal antibody (mAb; F99/97.6.1) 10 that was originally described as a part of the third-eyelid test. 25
In sheep, there is a strong association between allelic variation in PRNP and incidence of classical scrapie. 17 Unlike the sheep population, in which genetic selection has been implemented in an effort to decrease the incidence of scrapie, 9 genetic selection for scrapie resistance has not yet been implemented in goats. The lack of universal acceptance of a resistant allele is in part because of the high number of single nucleotide polymorphisms in caprine PRNP and observations of reduced risk or protection by candidate alleles in goats. Polymorphisms resulting in amino acid (aa) substitutions have yielded 10 PRNP haplotypes describing aa changes at codons 127, 142, 143, 146, 154, 211, 222, and 240 in a sample of U.S. goats. 37 These haplotypes have also been observed in goats worldwide. 36 Several of these alterations have been associated with delayed incubation or potentially a reduced risk of infection with scrapie. Such alterations include DNA encoding isoleucine (I) to methionine (M) at codon 142 (I142M),4,13 histidine (H) to arginine (R) at codon 143 (H143R), 6 asparagine (N) to serine (S) or aspartic acid (D) at codon 146 (N146S or N146D), R to H at codon 154 (R154H), R to glutamine (Q) at codon 211 (R211Q), and Q to lysine (K) at codon 222 (Q222K). 22 To date, there have been limited observations of scrapie-positive goats that are N146S or N146D 30 or when at least 1 allele encodes K at codon 222.1,7,22,38
A 2012 study demonstrated that allelic variation at codon 222 of the normal cellular prion protein (PrPC) in goats decreases epitope recognition by mAb F99/97.6.1. 24 It is critical that immunoassays for scrapie consider the potential for this or other alterations to become more prevalent in the goat population worldwide if selection schemes are implemented based on observations of reduced incidence or protection against disease. For example, the third-eyelid test in sheep was originally designed to include mAb F89/160.1.5, 25 but restriction to a single epitope may have yielded false-negative findings because of the presence of an alternative allele in crossbred sheep. 25 The goal of our study was to examine the impact of caprine PRNP allelic variation on detection of the scrapie agent by immunoassay. A combination of Western blot analyses for PrPC and IHC analyses for PrPSc using commercially available antibodies F99/97.6.1 (F99), F89/160.1.5 (F89), L42, and SAF84 were used to assess whether a multi-mAb approach or single antibody with an invariant epitope site may provide a more robust assessment of scrapie in goats than the current single antibody system because there would be a reduction in the possibility of false-negative results owing to allelic variation in regions of antibody epitope binding. To date, tissues from naturally infected goats carrying at least 1 allele encoding K222 are unavailable. Goats with at least 1 allele encoding R143 may also be moderately protected 6 but may have decreased epitope recognition by F89. Thus, goats with naturally acquired scrapie that were heterozygous at codon 143 were used as a surrogate to demonstrate the limitations of assays when using an antibody for which epitope variation within an animal population exists.
Materials and methods
Cell lines containing caprine PRNP harboring select polymorphisms
Archived DNA from goats previously determined to be homozygous for allelic variants at codons 142, 143, 146, 154, and 222 was initially isolated from peripheral blood collected into anticoagulant (ethylenediamine tetra-acetic acid) using either of 2 commercial kits.a,b The PRNP open reading frame was cloned into the pIRESpuro3 vector
c
for other studies in mammalian cells. This served as the DNA template for amplification and ligation into the pCR4-TOPO vector.
b
Cloned PRNP was excised by restriction digest with EcoRI and XhoI
d
and ligated into the pET30 vector
e
for expression with a His tag. The presence of single nucleotide polymorphisms of interest was confirmed by DNA sequence analysis as previously described
37
prior to transformation of BL21(DE3) Escherichia coli.
e
Kanamycin-resistant colonies were selected for growth, and expression of recombinant PrPC (rec-PrPC) was induced by addition of 1 mM isopropyl-β-
Animals and tissue samples
All animals included in our study were maintained, and procedures performed, in accordance with use approved by the Washington State University Institutional Animal Care and Use Committee (WSU-IACUC; ASAF 3811, ASAF 3815, ASAF 3819, and ASAF 4107). Animals were either naturally exposed to classical scrapie (n = 4), experimentally exposed to scrapie (n = 3), had limited exposure to scrapie (n = 5), or were never exposed to scrapie (n = 2). Naturally exposed animals were referred by U.S. Department of Agriculture (USDA) Animal and Plant Health Inspection Service from herds in which caprine scrapie had been confirmed. Experimentally exposed animals were inoculated orally within 1 week of birth at a USDA animal research unit (Pullman, Washington) with placenta from a goat that was infected with scrapie or brain and placenta from a goat confirmed to have scrapie. The limited exposure group included goats brought to the USDA research unit as breeding stock for studies of genetics-associated scrapie resistance or offspring born to them that were not included in inoculation studies. Finally, the 2 goats that were never exposed to scrapie were sourced from a scrapie-free herd maintained at Washington State University (Pullman, WA). The goats included in our study were of a variety of breeds (Table 1) and grouped for analyses based on PRNP haplotypes. At cull, all animals were euthanized and tissues collected according to WSU-IACUC–approved procedures. Tissues were trimmed of excess fat and either frozen at −20°C or fixed in 10% neutral buffered formalin.
Study population.
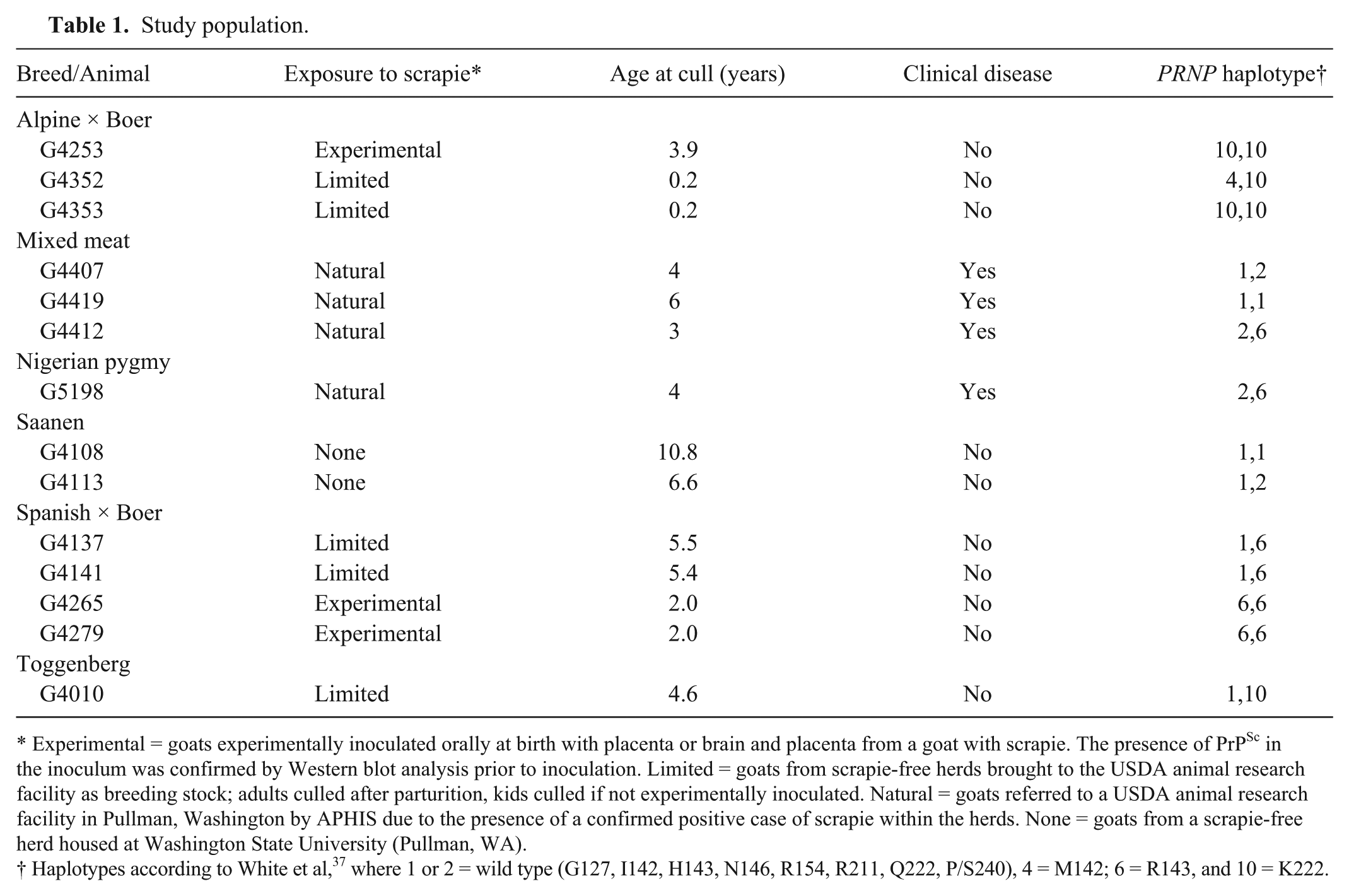
Experimental = goats experimentally inoculated orally at birth with placenta or brain and placenta from a goat with scrapie. The presence of PrPSc in the inoculum was confirmed by Western blot analysis prior to inoculation. Limited = goats from scrapie-free herds brought to the USDA animal research facility as breeding stock; adults culled after parturition, kids culled if not experimentally inoculated. Natural = goats referred to a USDA animal research facility in Pullman, Washington by APHIS due to the presence of a confirmed positive case of scrapie within the herds. None = goats from a scrapie-free herd housed at Washington State University (Pullman, WA).
Haplotypes according to White et al, 37 where 1 or 2 = wild type (G127, I142, H143, N146, R154, R211, Q222, P/S240), 4 = M142; 6 = R143, and 10 = K222.
Western blotting
Recombinant PrPC was assessed by bicinchoninic acid (BCA) protein assay h prior to loading equivalent protein concentrations for Western blot analyses. Tissue samples were prepared as 10% homogenates (w/v) by homogenizing 150 mg of brainstem rostral to the obex (wet weight equivalent) in direct lysis buffer (0.5% deoxycholic acid, 0.5% NP-40, 10 mM Tris–HCl; pH 7.5) with a tabletop homogenizer i on setting 5 for a total of 90 sec (2 × 45 sec). Total protein of homogenates was also estimated using BCA protein assay. Prior to electrophoresis, lithium dodecyl sulfate (LDS) sample buffer b and sample reducing reagent containing dithiothreitol b were added to all samples followed by boiling for 10 min. Samples (25 µg total protein for tissue homogenates; 40 ng or 60 ng for rec-PrPC) were loaded and electrophoresed through 12% Bis–Tris gels b and proteins transferred to polyvinylidene fluoride membrane as previously described. 28
Following protein transfer, membranes were blocked with blocker h containing 0.05% Tween 20 for 1 hr at room temperature. Membranes were incubated with mAbs overnight at 4°C with either F99/97.6.1 (3.5 µg/mL), F89/160.1.5 (3.5 µg/mL), L42 (0.02 µg/mL j ), or SAF84 (0.2 µg/mL k ) followed by washing and incubation for 40 min at room temperature with either horseradish peroxidase (HRP)-conjugated goat anti-mouse immunoglobulin G1 (IgG1; 1:6,000 l ) or HRP-conjugated goat anti-mouse IgG (H+L; 1:6,000 m ). Membranes were washed, incubated with enhanced chemiluminescent substrate, n and the chemiluminescent signal was captured on film. o Following detection of rec-PrPC, membranes were stripped h per the manufacturer’s instructions, and detection was performed with anti-His tag (0.02 µg/mL p ) and one of the secondary antibodies as described above. Equivalent loading of caprine brain homogenates was assessed in stripped membranes using anti–β-actin (0.1 µg/mL q ) and HRP-conjugated goat anti-rabbit IgG (H+L; 1:5,000 m ).
Immunohistochemistry
Postmortem brain tissue was processed and analyzed by automated immunohistochemical staining (IHC) as previously described29,33 with some modification. Briefly, the meninges of the brainstem were removed and a cross-section harvested from the obex region and fixed in 10% neutral buffered formalin for at least 24 hr before routine automated tissue processing and paraffin embedding. A minimum of 20 sequential thin (3 µm) sections of each paraffin-embedded obex hindbrain sample were prepared. Sections were collected out of a warm water bath onto glass slides, which were then air dried overnight and baked at 56°C for 30 min. Baked slides were immersed in 95% formic acid for 3 min at room temperature, washed 3 times with Trizma buffer (pH 7.0), o and loaded onto an automated slide staining system r for deparaffinization, antigen retrieval, s and IHC labeling. Serial sections were incubated with either F99/97.6.1 (17 µg/mL), or F89/160.1.5 (38 µg/mL) and reagents from a FastRed kit r for the detection of PrPSc. An irrelevant mAb of the same isotype was used at a concentration equivalent to the highest concentration of anti-PrP antibody to serve as a negative control. In contrast to the IHC protocol described above (in-house protocol), formic acid treatment was increased to 5 min, a manual deparaffinization step was used, the antigen retrieval step was modified, and an anti-mouse IgG and IgM detection system r was used to more closely align with the protocol used by the USDA National Veterinary Services Laboratory (SOP-PPE-0046). The modified method is hereafter referred to as the “NVSL-like protocol.” In addition to F99/97.6.1 and F89/160.1.5, mAbs L42 and SAF84 were tested using the conditions of the NVSL-like protocol.
Determination of PrPSc accumulation profiles
Obex sections were digitally photographed using a fluorescence microscope t equipped with an LED illuminating system, u digital camera, t and connected to a desktop computer running digital image processing software. t The entire surface area of each section was digitally acquired using a commercial software module t and a 1.25× objective. Image files were further processed in tiff format using the ImageJ-based open-source processing package Fiji (version 1.49a; http://pacific.mpi-cbg.de/). 32
Chromogen (FastRed) color was digitally extracted from background-corrected images (a posteriori method [Landini, G. How to correct background illumination in brightfield microscopy. Available from: http://imagejdocu.tudor.lu/doku.php?id=howto:working:how_to_correct_background_illumination_in_brightfield_microscopy&s[]=landini]) using the “Colour Deconvolution” plugin (Landini, G. Colour deconvolution. Available from: http://imagejdocu.tudor.lu/doku.php?id=plugin:color:colour_deconvolution:start) and user-defined color vector settings (hematoxylin stain vector: [r1] = 8.399071, [g1] = 4.038505, [b1] = 0.6539797; FastRed chromogen vector: [r2] = 13.301524, [g2] = 48.587498, [b2] = 19.41882; null vector: [r3] = 0.0, [g3] = 0.0, [b3] = 0.0). The suitability of these vector settings was confirmed by the lack of significant residual color in the null vector.
Each FastRed intensity image was subjected to threshold conversion by the Phansalkar method (Phansalkar N, et al. Adaptive local thresholding for detection of nuclei in diversity stained cytology images. In: Communications and Signal Processing (ICCSP), 2011 International Conference, 2011: 218–220. Calicut, Kerala, India; “Auto Local Threshold” plugin) using user-defined plugin settings for each IHC protocol (in-house: window radius = 6 pixels, k = 0.045, and r = 0.5; NVSL-like: window radius = 3 pixels, k = 0.01, and r = 0.5). Each section was subsequently divided into unit size-subregions such that the surface area of each obex section was represented by ~2,400 to 2,700 subregions, each measuring 50,000 sq μm and for which the local accumulation of PrPSc was expressed as the percent exceeding threshold (Fiji, “Measure” tool, http://pacific.mpi-cbg.de/). Local accumulation data were imported into SAS, v and the empirical distribution function for each section was generated using the NPAR1WAY procedure and “EDF” option, which expresses the cumulative proportion of subregions (y-axis, 0.0–1.0) having a given or less than percent threshold immunolabeling (x-axis, 0–100%).
Results
Allele effects on Western blot analyses of PrPC
To examine the impact of allelic variation on Western blot analyses, caprine PRNP haplotypes were expressed as rec-PrPC, including wild-type (I142, H143, N146, R154, Q222) and sequence variants singly coding M142, R143, S146, H154, or K222. These haplotypes were probed by Western blot for epitope effects on antibodies developed for testing use in the United States (F99 and F89).5,10,25,27,33 Antibody F99 targets a conserved epitope in the C-terminus of PrP at aa 220–225 (QYQRES) 25 whereas the epitope for F89 is aa 142–145 (IHFG). 26 Of the caprine haplotypes tested, F99 failed to detect only the PrP variant K222 even though the quantity of this His-tagged rec-PrPC was equivalent or greater than the other recombinant variants in the blot (Fig. 1A). Antibody F89 successfully detected rec-PrPC when expressed as wild type, S146, H154, or K222, whereas the presence of M142 or R143 abolished the signal. Detection with 2 additional antibodies, L42 (detecting aa 148–153, YEDRYY 23 ) or SAF84 (detecting aa 166–172, YRPVDQY; deduced for goat based on the sequence recognized in bovine prion protein 18 ) was not altered by any of the allelic variants when compared with wild type.
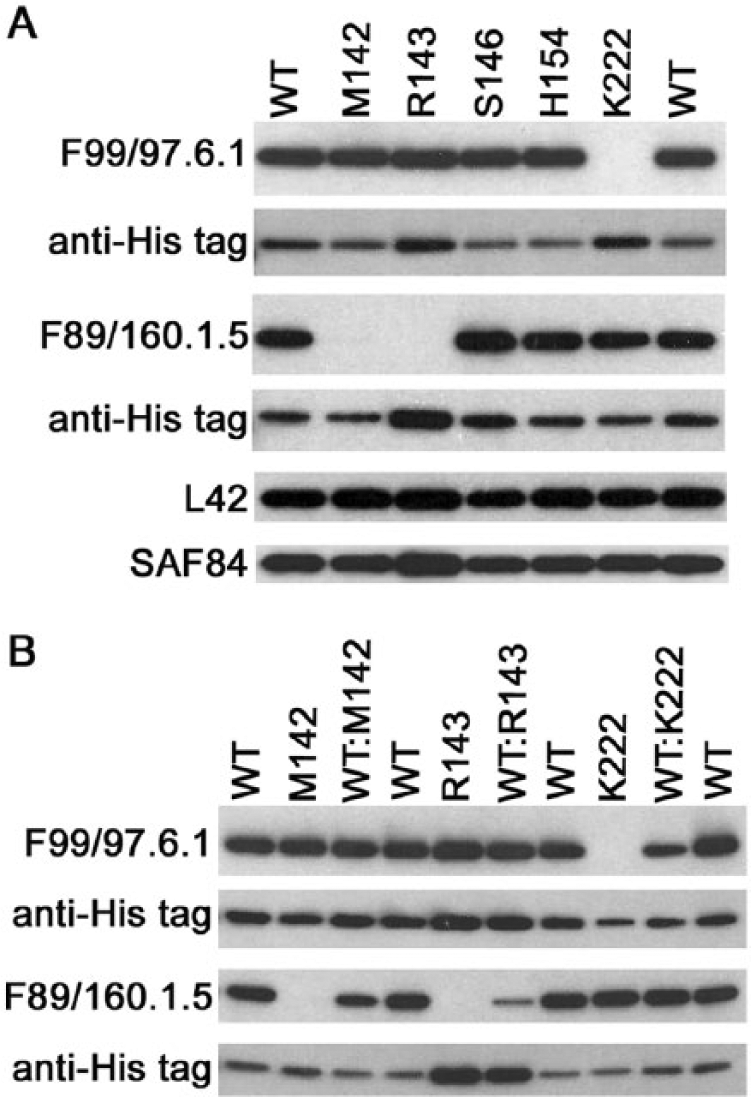
Epitope-associated sensitivity in recombinant PrPC (rec-PrPC) representing wild-type PRNP (WT; I142, H143, N146, R154, Q222) and individual alternative caprine alleles.
The series of Western blots used to examine the influence of individual alleles on detection are representative of what would be expected in goats that are homozygous for PRNP. Next, individual alleles were mixed at a ratio of 1:1 (wild-type to allelic variant) prior to Western blot analyses as a demonstration of what may be observed in PRNP heterozygous goats (Fig. 1B). Total protein was loaded at a modestly lower level compared with analysis of individual alleles to ensure that the signal was not saturated and that differences resulting from the presence of multiple alleles could be visualized. The signal generated by detection with F99 was reduced when wild type and K222 PrPC were mixed. The presence of M142 or R143 with wild-type PrPC resulted in a reduced signal when detection was performed with F89. The impact of R143 is even more pronounced when protein loading was confirmed through detection of the His tag, despite efforts to ensure equivalent sample loading across the blot. These results were anticipated based on previous observations 24 and known epitope-binding regions for both F99 and F89.
Not only were decreases observed with rec-PrPC, but the impact of aa changes was also observed in normal caprine brain tissue homogenates. Western blot analyses were performed using caprine brain expressing wild-type PRNP, heterozygous HR143, or homozygous RR143 (Fig. 2) or wild-type, heterozygous QK222, or homozygous KK222 (Fig. 3; n = 2 per group). Normal cellular prion protein was readily detected with F99, L42, and SAF84 in wild-type animals and in goats expressing PRNP variants at codon 143 (Fig. 2), whereas the signal for F89 was dramatically reduced in HR143 animals and almost completely abrogated when both alleles encoded R143 (Fig. 2). In addition, F89 did not perform as well as the other antibodies on wild-type PrPC (film exposure time of 30 min vs. 3 min for all other antibodies). The reduced signal observed with F89 did not result from variation in protein loading as was demonstrated by a strong signal when detection was performed with anti–β-actin following membrane stripping. The overall signal with F89 was also reduced when analyzing PrPC from goats with QK222 or KK222 requiring a longer film exposure (20 min), but PrPC was detected in all samples similar to what was observed with L42 and SAF84 (Fig. 3). This is in contrast to detection with F99, which was greatly reduced in animals with the QK222 alleles, and complete abrogation was observed in goats with KK222. Detection with anti–β-actin confirmed that the reduction with F99 was not due to variation in protein loading.
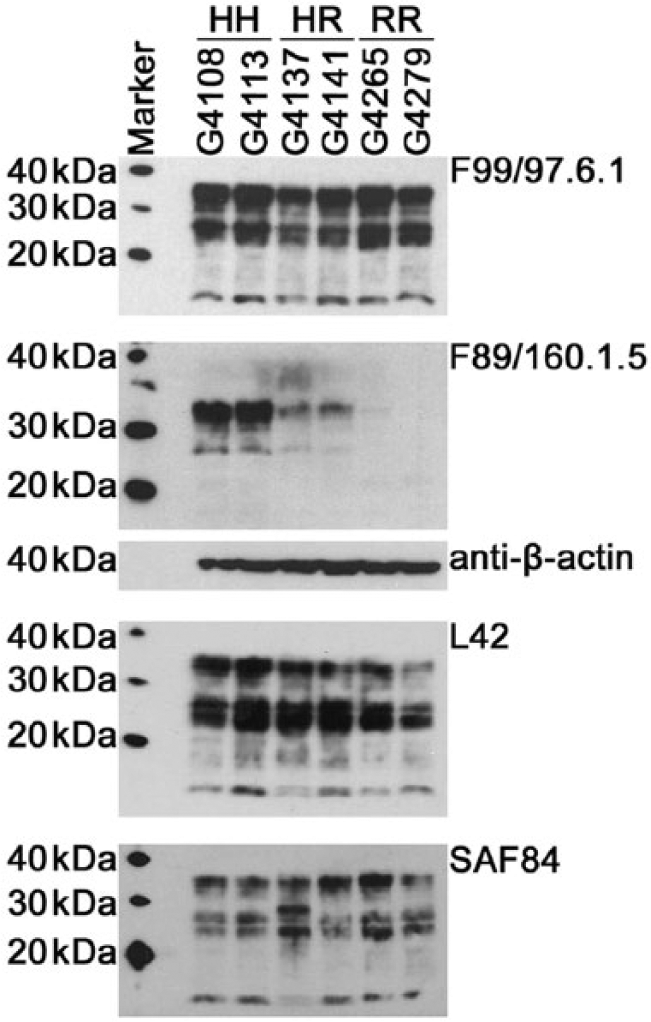
Western blot analyses of PrPC from goats homozygous or heterozygous at codon 143. Homogenates (10%) were prepared from brainstem immediately anterior to the obex, and 25 µg of total protein was loaded in each lane. Detection was performed with monoclonal antibodies F99/97.6.1, F89/160.1.5, L42, or SAF84. Membranes were stripped following detection with anti-PrP antibody and re-probed with β-actin (representative image shown for membrane initially used for detection with F89/16.1.5).
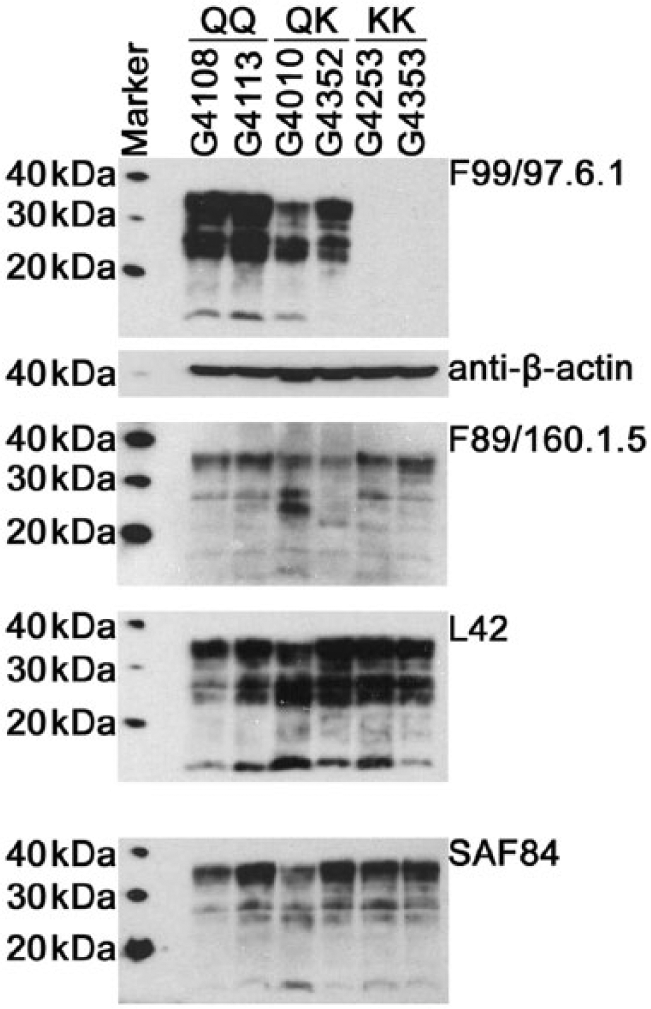
Western blot analyses of PrPC from goats homozygous or heterozygous at codon 222. Homogenates (10%) were prepared from brainstem immediately anterior to the obex, and 25 µg of total protein was loaded in each lane. Detection was performed with monoclonal antibodies F99/97.6.1, F89/160.1.5, L42, or SAF84. Membranes were stripped following detection with anti-PrP antibody and re-probed with β-actin (representative image shown for membrane initially used for detection with F99/97.6.1).
Allele effects on IHC analyses of PrPSc
Immunohistochemical analysis is a mainstay of scrapie testing in the United States. 5 Prior to determining the impact of allele effects on detection of PrPSc by IHC with in-house single antibody scrapie IHC protocols for F99 and F89, antibody concentrations were titrated and formic acid treatments (0 vs. 3 vs. 5 min on slide) were compared (data not shown). Efforts were made to optimize for strong labeling of positive aggregates in the dorsal motor nucleus of the vagus nerve and clean labeling, which lacked pink hues. The in-house protocol was then applied to formalin-fixed, paraffin-embedded thin sections of obex hindbrain collected from goats displaying clinical signs of scrapie at cull. Obex hindbrain samples representing similar stages of clinical disease were available from 2 wild-type goats (G4407 and G4419) and 1 goat heterozygous at codon 143 (HR143; G4412). The F99 immunolabeling of PrPSc in the obex was similar between wild-type and the HR143 goats (Fig. 4, left column). In comparison with F99, F89 immunolabeling appeared considerably less intense in the obex of a HR143 goat (Fig. 4, middle column) even though F89 immunolabeling appeared equivalent to robust F99 immunolabeling in the obex of a wild-type goat (Fig. 4, top row). Minimal background labeling (pink hues) was observed with an irrelevant antibody isotype control (Fig. 4, right column).
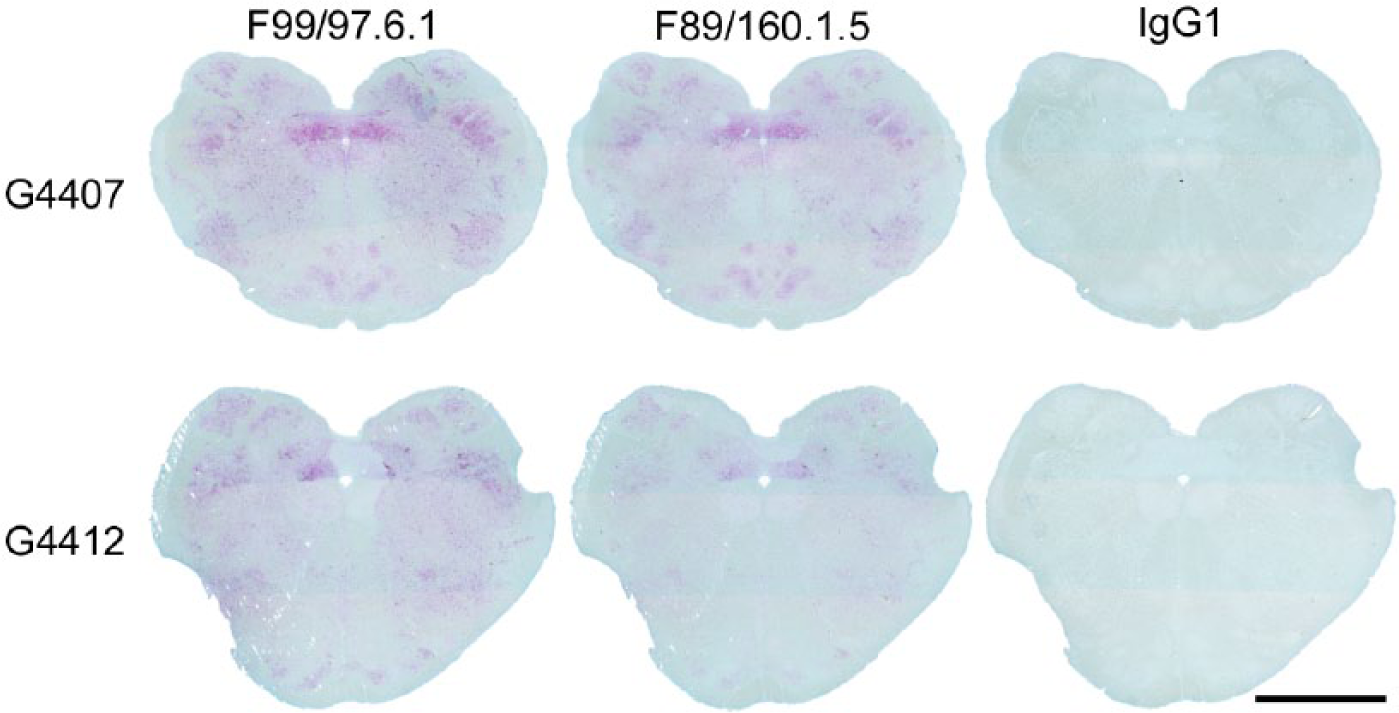
The comparative effects of PRNP HR143 using the in-house protocol with F99/97.6.1 and F89/160.1.5 on obex hindbrain of goats with clinical scrapie at time of cull. Top row: serial sections from a representative PRNP wild-type goat (G4407). Bottom row: serial sections from a PRNP HR143 goat (G4412). Immunolabeling was performed with a commercial kit r and in-house preparations of F99 at 17 µg/mL, F89 at 38 µg/mL, and an irrelevant antibody (isotype control) at 38 µg/mL. Bar = 5 mm.
To ensure this difference was observed under near equivalent effects of primary antibody, a 4-point doubling dilution titration of F89 and F99 on serial sections was performed (Fig. 5). In PRNP wild-type goats, the spatial patterns (insets) and accumulation profiles (graphs) of PrPSc threshold immunolabeling by F89 and F99 appeared similarly dependent on primary antibody concentration (Fig. 5, left and right columns). The spatial pattern of PrPSc detected by F99 in the obex of the HR143 goat (middle column) was also similar to that observed in the wild-type goats. The accumulation profiles of PrPSc as detected by F99 at 34 and 17 μg/mL were slightly less intense (shifted left) in the HR143 goat as compared to that detected in either wild-type goat. In contrast, very little of the PrPSc detected by F99 in the obex of the HR143 goat was detected by even the highest concentration of F89 (Fig. 5, top, middle column).
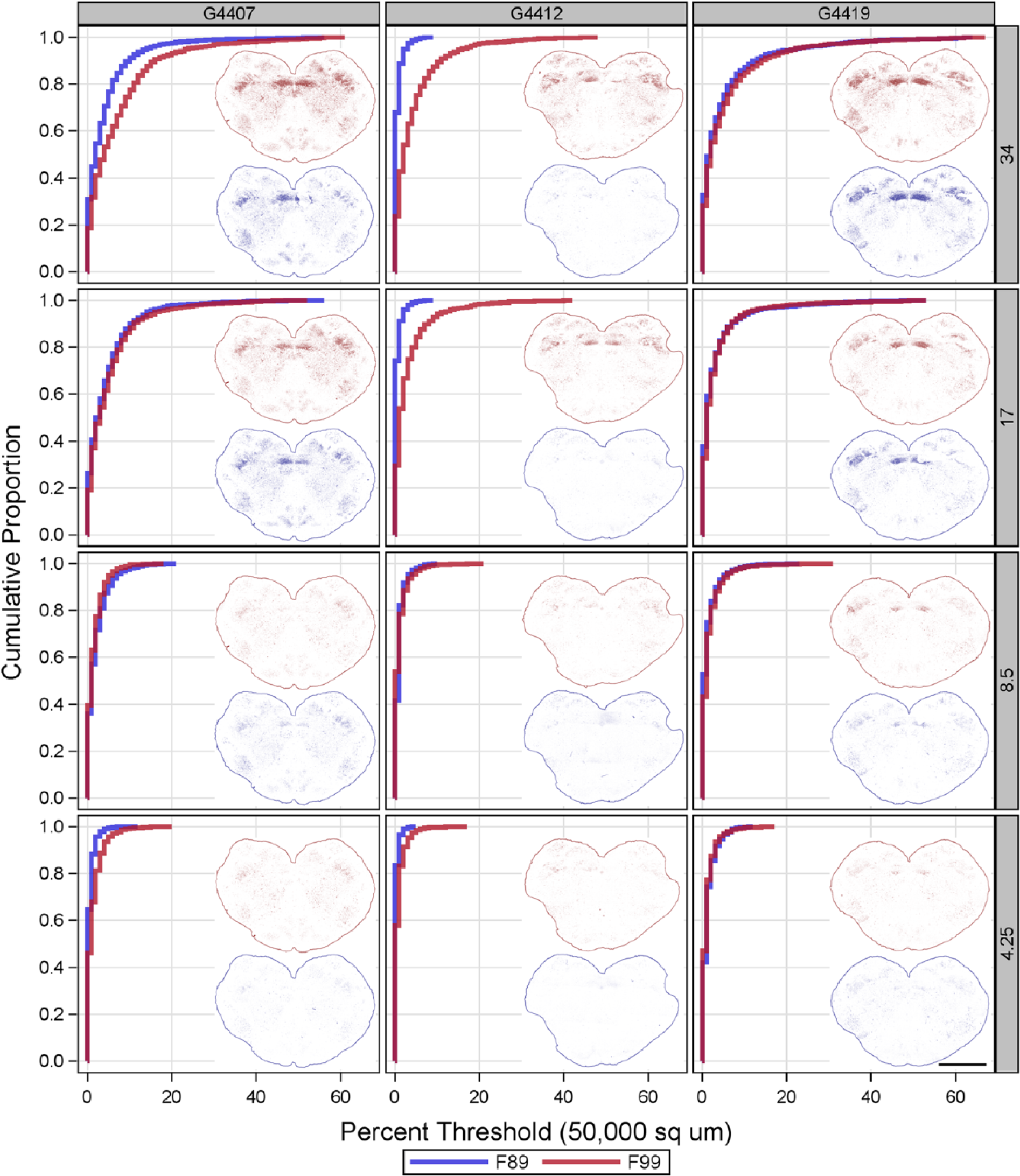
The comparative effects of PRNP HR143 polymorphism on titrated F89/160.1.5 (F89) and F99/97.6.1 (F99) immunolabeling using the in-house protocol on the obex hindbrain of goats with clinical scrapie. Each column of graphs and inset images show the paired results for a given concentration (µg/mL; listed on the right) of F89 (blue) and F99 (red). Inset images show the spatial pattern of threshold immunolabeling (outlines added to indicate tissue margins), and graphs depict the corresponding empirical distribution functions. The concentration-dependence and accumulation profile of PrPSc as detected by F89 and F99 were very similar in the obex of wild-type goats. Threshold immunolabeling in the obex from the HR143 goat was minimal at all concentrations of F89, despite the detection by F99 of PrPSc accumulation similar to that in the wild-type goats. Bar = 5 mm.
The effects of PRNP genotype on F99 and F89 immunolabeling were also tested using a kit and protocol approved for use by U.S. regulatory laboratories. Since a commercial preparation of F89 is not currently available with this kit, the effects of genotype on immunolabeling using the NVSL-like protocol were evaluated using in-house supplies of F99 and F89. Using tissue from a wild-type goat, titration of F99 and F89 under the NVSL-like protocol was first determined by serial dilution (2-fold dilutions from 136 to 4.25 μg/mL) similar to that described above. Significant chromogen was produced diffusely over the slide only when F99 or F89 was used at 136 μg/mL (data not shown). Immunolabeling produced by F99 at 68 μg/mL appeared equivalent to that produced by the F99 included in the manufacturer’s kit (Fig. 6, A1 vs. A2), but immunolabeling by F89 (Fig. 6A3) was significantly weaker at all antibody concentrations tested. Next, the effects of PRNP genotype on PrPSc detection using the NVSL-like protocol were evaluated at 68 μg/mL of F99 and F89. The impact of PRNP genotype on PrPSc detection under the NVSL-like protocol was evaluated by comparing spatial pattern and accumulation profiles of threshold immunolabeling. For this, the remaining serial sections from the 2 wild-type goats and one HR143 goat were used. A serial slide set of obex from an additional clinical field case, a HR143 Nigerian pygmy goat (G5198) with naturally acquired scrapie, was also included. As shown in Figure 6B, PrPSc threshold immunolabeling by F99 (red) appeared similarly robust in the 2 wild-type goats (upper row) and slightly less (G4412) or moderately less (G5198) in HR143 goats (lower row). Under the NVSL-like protocol, threshold immunolabeling by F89 in comparison to F99 (blue vs. red) was relatively poor in wild-type goats (upper row) and nearly absent in HR143 goats (lower row). In wild-type goats, however, subregions with considerable amounts of F89 immunolabeling (40–60% of the subregion greater than threshold) were still readily apparent (e.g., the dorsal motor nucleus of the vagus nerve).
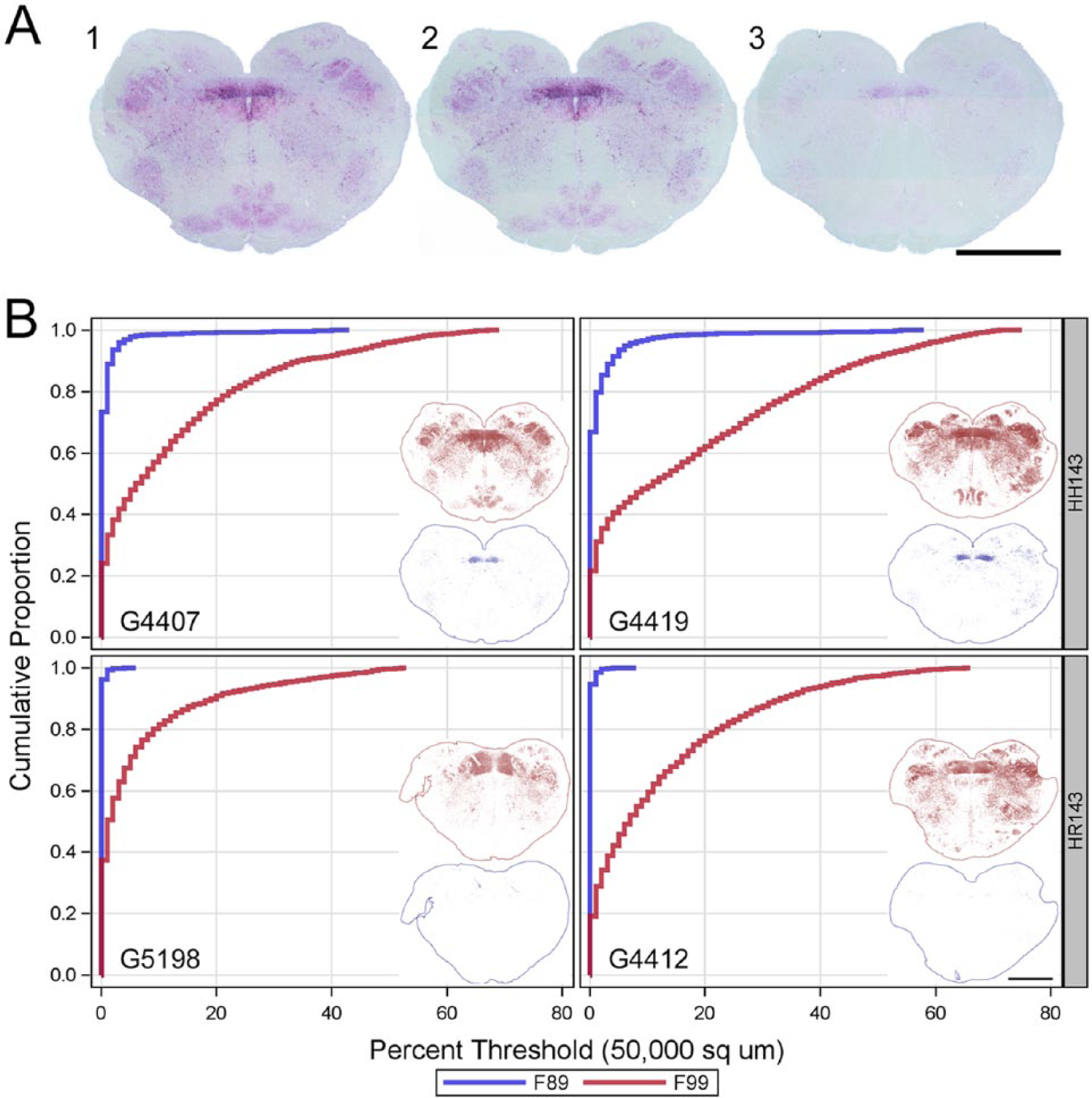
The comparative effects of PRNP HR143 using the NVSL-like protocol on the obex hindbrain of goats with clinical scrapie.
Because of the reduced reactivity of F89 using the NVSL-like protocol, it was of interest to see if antibodies other than F99 performed better under these conditions. Western blot analyses had demonstrated that the other allelic variants tested did not impact detection of rec-PrPC with L42 or SAF84 (Fig. 1A). Thus, a limited doubling dilution titration experiment was performed (data not shown), and the concentrations that yielded uniform, strong labeling were applied to obex hindbrain sections from wild-type and HR143 goats (Fig. 7). Unlike our observations with F89, L42 performed similarly to what was observed with F99. Immunolabeling with SAF84 was moderate compared to F99 and L42 under these conditions but improved relative to F89. Although the concentrations used yielded uniform, strong labeling of the obex, a diffuse background of red chromogen was observed in obex hindbrain of a scrapie-naive, wild-type goat (G4413). Thus, other antibodies may work within the NVSL-like protocol, but optimization and validation of any additions or substitutions will be needed before modifying the current diagnostic protocol.
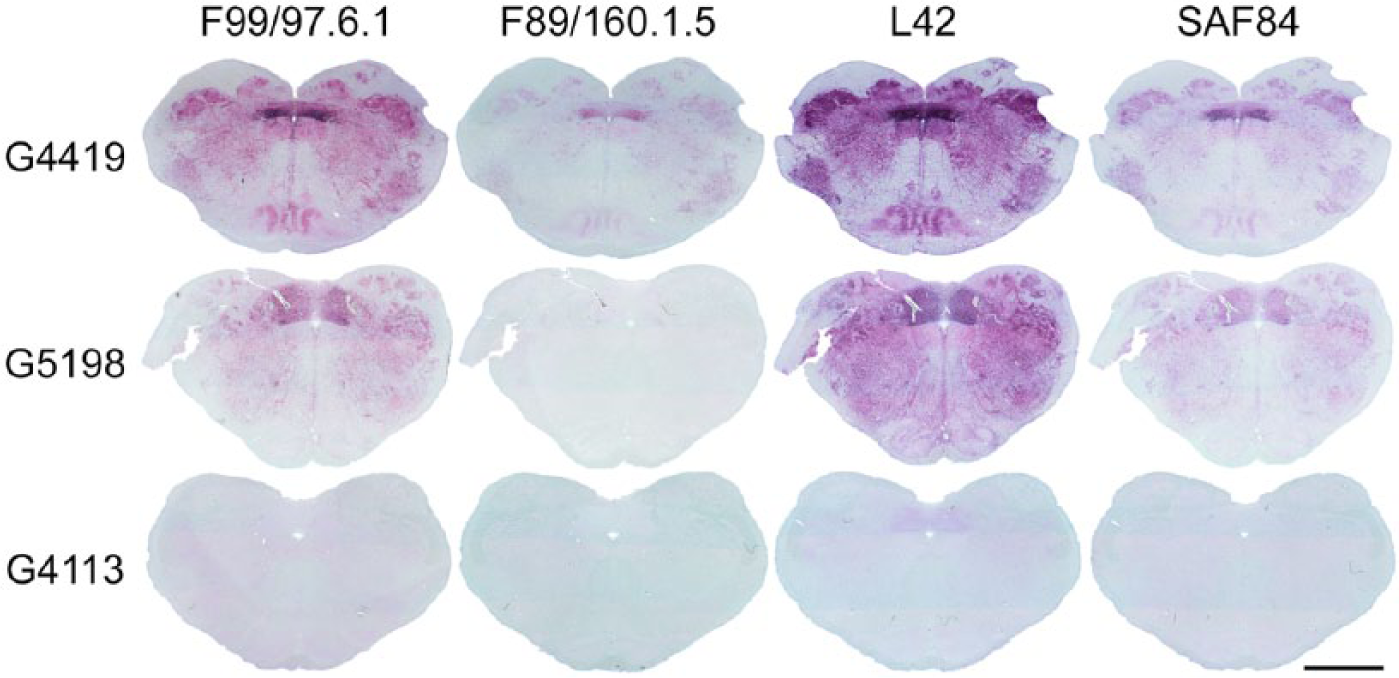
Comparison of PrPSc immunolabeling by different antibodies under the NVSL-like protocol. Shown are the results for serial thin sections of obex hindbrain from 2 adult goats with clinical scrapie but having different PRNP genotypes (wild type [G4419] and HR143 [G5198]) and from a scrapie-naive, wild-type goat. Under these conditions, strong immunolabeling was produced for thin sections from both clinical goats when using monoclonal antibodies F99/97.1.6 (68 μg/mL) and L42 (0.25 μg/mL). In comparison, immunolabeling produced by SAF84 (1 μg/mL) was only moderate under these conditions but did not appear to be affected by PRNP genotype at codon 143. In contrast, immunolabeling produced by F89/160.1.5 (68 µg/mL) was poor under these conditions and further reduced in the HR143 goat. A diffuse background of light intensity red chromogen was evident in the obex hindbrain of a scrapie-naive, wild-type goat when immunolabeling was performed with F99/97.1.6 and L42, an indication that use of lower concentrations of these antibodies might be more optimal for this IHC protocol. Bar = 5 mm.
Discussion
In the early 2000s, the National Scrapie Eradication program was initiated by the Animal and Plant Health Inspection Service of the USDA with the goal of eradicating scrapie from the United States and subsequently meeting criteria set forth by the World Organization for Animal Health (OIE) to gain disease-free status. This program uses a combination of regulatory scrapie slaughter surveillance, nonslaughter surveillance, and a scrapie-free flock certification program as described in the USDA National Scrapie Surveillance Plan (2010, http://www.aphis.usda.gov/animal_health/animal_diseases/scrapie/downloads/national_scrapie_surv_plan.pdf) to identify infected animals. Through monitoring of sheep and goats and genetic selection in sheep, the identification of new cases continues to decrease (http://www.aphis.usda.gov/animal_health/animal_diseases/scrapie/downloads/annual_report.pdf). The majority of cases have been observed in sheep flocks, but it is equally important to continue monitoring goat herds as a reservoir for transmission to sheep. 21 The antibody used for antemortem and postmortem IHC testing in the United States, F99, robustly detects PrPSc in sheep and most goats. A 2012 study demonstrated decreased detection of PrPC with F99 by Western blot in goats with at least 1 allele encoding K222 and the complete absence of signal in K222 homozygotes. 24 In our study, we assessed the performance of several antibodies for the detection of PrPC by Western blotting and PrPSc by IHC when the caprine prion protein was expressed as wild type or several allelic variants that are present in the United States. 37 In addition to F99, our study focused primarily on F89, because this antibody was included in the original third-eyelid testing scheme for sheep 25 ; the aa epitope it recognizes includes sites of allelic variation in caprine PRNP that may be associated with delayed incubation of disease or reduced risk of infection,4,6,8,13,14 and naturally infected goats with this allelic variation have been identified in the United States. Individually, detection of PrPC with F89 or F99 is decreased by the presence of at least 1 allelic variant in the antibody epitope-binding region with complete abrogation being observed when both alleles are encoded by sequence variants (Figs. 1–3). Furthermore, this study demonstrated that presence of an allelic variant can alter detection of PrPSc by IHC (Figs. 4–7). These observations have the potential to influence scrapie eradication efforts in the United States.
Genetic selection for relatively resistant PRNP alleles in sheep has been a major factor in decreasing the prevalence of scrapie. 9 The wild-type haplotype for caprine PRNP (G127, I142, H143, N146, R154, R211, Q222, P/S240) is present at a frequency of 75% when estimated across the 10 most common breeds found in the United States, 37 and goats homozygous for this haplotype are susceptible to scrapie. Polymorphisms exist at various frequencies in caprine PRNP, several of which have been examined as possible candidates for genetic selection. Animals expressing at least 1 allele encoding M142, R143, S146, or D146 are susceptible to disease but have demonstrated delays in incubation or reduced risk of infection relative to wild type.4,6,8,13,14,30 Studies have begun to focus on the K222 PRNP allele, which was estimated in 2008 to occur at a frequency of 0.6% in the United States. 37 Few natural cases of scrapie have been reported in goats heterozygous at codon 222,7,8,11 whereas orally inoculated animals remained negative for at least 5 years.22,38 Observations following intracranial challenge of goats with PRNP QK222 have shown animals to be negative after ~4.5 years post-challenge 1 but positive in a different study. 22 Additional work using transgenic mice homozygous for K222 has shown a resistance to scrapie from goats but susceptibility to bovine spongiform encephalopathy (BSE) and goat-passaged BSE. 2 Although the K222 allele does not confer complete resistance to scrapie, this allelic variant has been suggested as a plausible target for genetic selection in goats in Europe.1,2,20,22 If the United States also adopts this strategy, the current diagnostic testing protocols with F99 will need to be re-evaluated. The results of our study (Figs. 1, 3), and others, 24 clearly demonstrate that the presence of a sequence variant at 222 reduces sensitivity of Western blot detection of PrPC by F99. This difference is also anticipated to be observed when analyzing PrPSc by Western blot, IHC, or other immunoassays that use F99, and could lead to an increased number of false-negative results. Whereas samples from scrapie-positive animals harboring the K222 allele were not available for our study, scrapie-positive animals with the R143 allele were used as a surrogate to demonstrate the limitations of a single antibody approach in the presence of an alternative allele. Like F99 detection in animals with at least one K222 allele, Western blot sensitivity for PrPC was reduced for F89 when at least 1 allele encoded R143 (Figs. 1, 2). With a limited sample population, similar observations were made when assessing PrPSc by IHC in wild-type versus HR143 goats (Figs. 4–7). The animals included in the IHC analyses were displaying signs of clinical disease at the time of sample collection, but in a preclinical animal, the impact on immunoassay detection could be even more profound. In a scenario with K222 present as the alternative allele and F99 the antibody for diagnostic detection, it is anticipated that results would be similar to what was observed with F89 and HR143 goats. Thus, if the suggestion is to begin selective breeding for K222, it will be critical to evaluate the robustness of current tests used in the United States in the identification and monitoring of scrapie in goats.
Whereas testing in the United States currently uses F99, 10 and some assays were originally developed to include F89, 25 a variety of other anti-PrP antibodies are available and used in studies of scrapie. Two such antibodies were included in our study. As anticipated, detection was similar across all allelic variants tested when L42 or SAF84 was used in Western blotting (Figs. 1–3). Polymorphisms within the SAF84 epitope (aa 167–172) do not appear to be highly prevalent in U.S. goats, 37 but an allelic variant at codon 168 that impacts Western blot detection with this antibody has been identified in herds in Greece. 11 Detection with L42 (Fig. 1) was not affected by alterations occurring near (S146) or adjacent (H154) to the antibody epitope region (aa 148-153). In addition, both SAF84 and L42 appear to perform well in the current NVSL-like protocol for IHC (Fig. 7). Several other anti-PrP antibodies that are commonly used in prion research were not included in our study. For example, epitopes for P4 and 8G8 do not include highly polymorphic sites found in caprine PRNP but they do include sites of differential proteinase K cleavage and can be used in studies of strain discrimination.12,34 Although this is an important aspect of understanding the etiology of disease, it may be more beneficial to reserve P4 and 8G8 for strain discrimination or further characterization while using antibodies that detect more central (e.g., F89, L42, SAF84) or C-terminal (e.g., F99) epitopes for general scrapie testing.
The United States has made great strides toward the goal of scrapie eradication through the use of genetic selection in sheep and surveillance sampling of sheep and goats. When genotype information is lacking, observations in our study support the notion that variation of current protocols, possibly including a transition to a different single antibody (such as shown in Fig. 7) or a multi-antibody scheme as was originally used for the detection of classical scrapie in U.S. sheep, may bring more confidence to results. A potential need for adaptation will continue to grow if genetic selection of genotypes demonstrating a reduced risk of disease, such as K222 in goats, is implemented as a component of disease eradication efforts.
Footnotes
Acknowledgements
S. Madsen-Bouterse and D. Schneider contributed equally to this study. We are grateful for the assistance of the USDA-ARS-ADRU animal care staff, and E. Karel and WSU-ARU for care of the animals, L. Fuller for animal management and assistance with IHC, and D. Lesiak for assistance with PRNP genotyping. We are particularly grateful to USDA-APHIS-NVSL for helping us obtain thin sections from a natural case of clinical scrapie in a HR143 goat doe (G5198).
Authors’ note
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
a.
Gentra PureGene blood kit, Qiagen Inc., Valencia, CA.
b.
Invitrogen iPrep PureLink gDNA blood kit, NuPAGE buffer NuPAGE reagent NuPAGE gels; Life Technologies, Carlsbad, CA.
c.
Clontech Laboratories Inc., Mountain View, CA.
d.
New England Biolabs Inc., Ipswich, MA.
e.
BugBuster Plus Lysonase, EMD Millipore Corp., Billerica, MA.
f.
Bioline USA Inc., Taunton, MA.
g.
MCLAB, South San Francisco, CA.
h.
Blocker Casein in PBS, Restore Plus Western blot stripping buffer; Thermo Fisher Scientific/Pierce, Rockford, IL.
i.
FastPrep FP120, Thermo Electron Co., Waltham, MA.
j.
R-Biopharm AG, Darmstadt, Germany.
k.
Cayman Chemical Co., Ann Arbor, MI.
l.
Southern Biotech, Birmingham, AL.
m.
KPL Inc., Gaithersburg, MD.
n.
Amersham ECL, GE Healthcare Life Sciences, Piscataway, NJ.
o.
Kodak BioMax, Trizma Pre-set crystals, pH 7.0; Sigma-Aldrich, St. Louis, MO.
p.
GenScript USA Inc., Piscataway, NJ.
q.
Abcam Inc., Cambridge, MA.
r.
Discovery XT, DISCOVERY Red Map kit, DISCOVERY UltraMap anti-Ms Alk Phos kit; Ventana Medical Systems Inc., Tucson, AZ.
s.
Target retrieval solution, Dako North America Inc., Carpinteria, CA.
t.
Axio Imager.M1 microscope, AxioCam MRc5 digital camera, AxioVision v. 4.8.1.0, AxioVision 4 Module Panorama; Carl Ziess Microscopy, Thornwood, NY.
u.
X-cite Series 120, Excelitas Technologies Corp., Waltham, MA.
v.
SAS v. 9.3, SAS Institute Inc., Cary, NC.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the USDA Agricultural Research Service under CRIS 2090-32000-030-00.