
Abstract
A captive, juvenile, female northern map turtle (Graptemys geographica) was found dead following a brief period of weakness and nasal discharge. Postmortem examination identified pneumonia with necrosis and numerous epithelial, intranuclear viral inclusion bodies, consistent with herpesviral pneumonia. Similar intranuclear inclusions were also associated with foci of hepatocellular and splenic necrosis. Polymerase chain reaction (PCR) screening of fresh, frozen liver for the herpesviral DNA-dependent DNA polymerase gene yielded an amplicon with 99.2% similarity to recently described emydid herpesvirus 1 (EmyHV-1). Molecular screening of turtles housed in enclosures that shared a common circulation system with the affected map turtle identified 4 asymptomatic, EmyHV-1 PCR-positive painted turtles (Chrysemys picta) and 1 asymptomatic northern map turtle. Herpesvirus transmission between painted and map turtles has been previously suggested, and our report provides the molecular characterization of a herpesvirus in asymptomatic painted turtles that can cause fatal herpesvirus-associated disease in northern map turtles.
Herpesviruses are ubiquitous pathogens of the animal kingdom, and have been documented in a diverse range of hosts, including mammals, birds, reptiles, amphibians, fish, and invertebrates. 4 Infection is frequently subclinical or mild in infected natural host species, which is proposed to be the result of coevolution of the virus and host. Severe or fatal infections are most commonly seen in immunosuppressed natural hosts or when a herpesvirus of one species infects a closely related species, as has been documented in multiple instances.13,16,18
Several clinically significant herpesviruses have been identified in chelonians (turtles and tortoises). In marine turtles, manifestations of disease include epidermal necrosis, pneumonia, stomatitis, and fibropapilloma formation.7,9,20 In tortoises, herpesviruses have been associated with oral and glossal inflammation.10,15,17 In freshwater turtles, fatal herpesvirus-like disease has been described histologically in Pacific pond turtles (Actinemys marmorata), painted turtles (Chrysemys picta), and map turtles (Graptemys spp.).3,5,11 Reports that combine molecular viral characterization with clinical findings in chelonians are limited. Such reports include a description of terrapene herpesvirus 1 infection with or without concurrent ranaviral infection in captive eastern box turtles (Terrapene carolina carolina) 19 and emydid herpesvirus 1 (EmyHV-1) in an eastern river cooter (Pseudemys concinna concinna). 12
In March of 2012, a captive, juvenile, female northern map turtle (Graptemys geographica) from a zoological institution was noted to be weak with a frothy nasal discharge. This individual was 1 of a group of 8 recently acquired northern map turtles being housed under quarantine conditions. Radiographs revealed patchy irregular to diffusely increased radiopacifi-cation in both lung fields. Despite supportive treatment with parenteral isotonic fluids and a broad-spectrum antibiotic (ceftazidime) for suspected bacterial pneumonia, it was found deceased later that day and was submitted for postmortem evaluation. Grossly, the lungs were diffusely dark red, wet, and heavy. The cranial portions of the lung were thickened, wet, and gelatinous and drained a moderate amount of watery fluid on section. Trace amounts of similar fluid were present in the trachea. The liver was diffusely tan, and the edges of all lobes were slightly rounded. No other gross abnormalities were noted. A sample of fresh liver was collected and archived frozen (–80°C). A sample of lung submitted for aerobic and anaerobic bacterial culture yielded the growth of moderate numbers of Flavobacterium spp. and many Chryseobacterium indologenes. A complete set of tissues was collected, fixed, and stored in 10% neutral buffered formalin at room temperature for 8 days, and processed routinely for histologic examination. Paraffin sections were cut at 5 μm and stained with hematoxylin and eosin.
Significant lesions were present in the lung, liver, and spleen. Primary changes in the lungs consisted of pulmonary epithelial necrosis with large numbers of pulmonary and bronchial, intraepithelial, intranuclear, pale amphophilic to eosinophilic, 2–4 μm in diameter, Cowdry type A viral inclusions (Fig. 1A). Heterophils and lymphocytes were present in the pulmonary interstitium and bronchial submucosa, and there was diffuse, acute congestion of the pulmonary vasculature with multifocal hemorrhage and the accumulation of proteinaceous fluid within airways. In the liver, foci of acute hepatocellular necrosis were associated with eosinophilic intranuclear viral inclusions similar to those seen in the lung (Fig. 1B). In the spleen, fibrinoid vascular necrosis was present with rare intraendothelial, intranuclear viral inclusions.
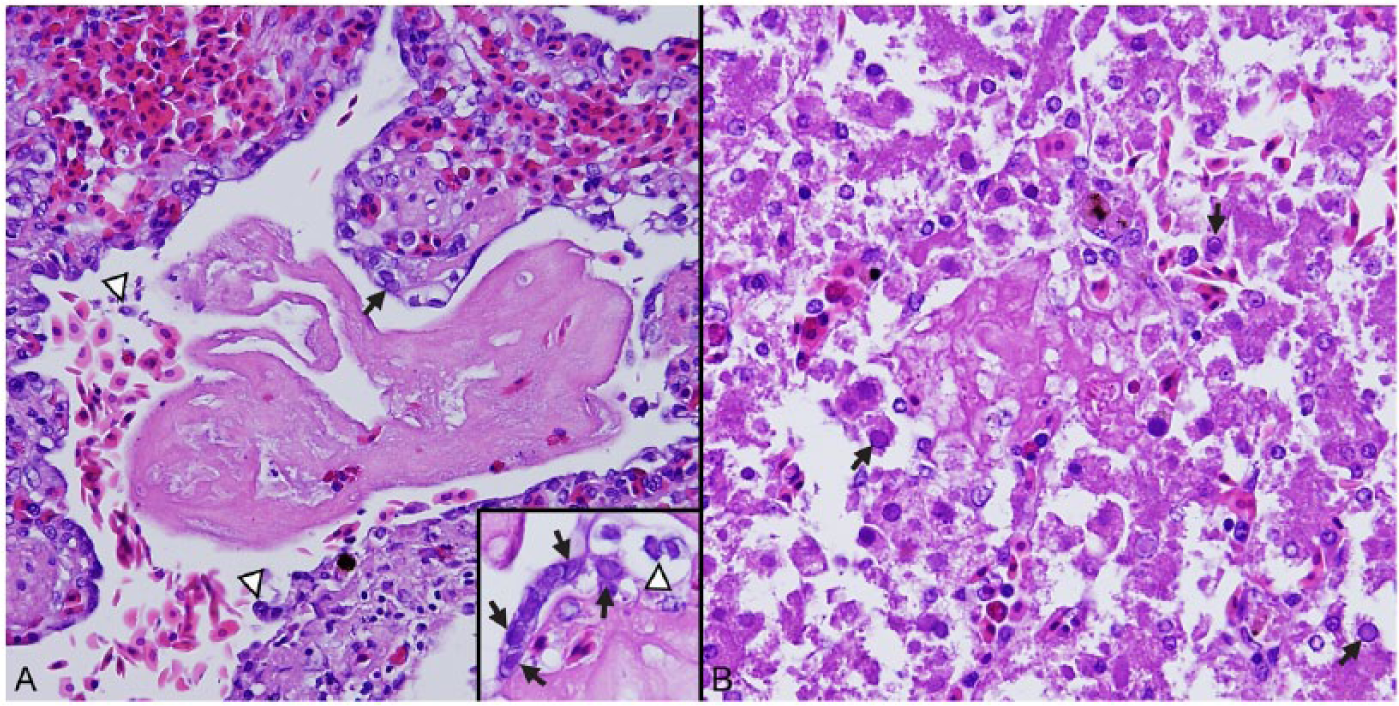
Lesions in a northern map turtle with emydid herpesvirus 1–associated disease.
Nucleic acids were extracted and purified from the sample of fresh, frozen liver using a commercial kit a according to the manufacturer’s instructions. Qualitative nested polymerase chain reaction (PCR) amplification of a short fragment of the DNA-dependent DNA polymerase was performed using previously reported methods. 21 Reactions contained the following: 12.5 μL of a commercial master mix, b 25 pmol each of degenerate primers DFA, ILK, and KG1 (round 1) or TGV and IYG (round 2), 2 μL of either the nucleic acid extract (round 1) or the round 1 reaction product (round 2), and molecular-grade water to bring the total reaction volume to 25 μL. Reaction conditions were as follows: 95°C for 12 min; 45 cycles at 95°C for 20 sec, 46°C for 60 sec, and 72°C for 60 sec; and 72°C for 10 min. A commercially prepared plasmid containing primer binding sites was used as a positive control. An ~220–base pair (bp) amplicon was visualized with gel electrophoresis and stained. c To obtain a larger frag-ment of the DNA-dependent DNA polymerase, 2 altered second-round PCR reactions were performed using primers DFA and IYG or TGV and KG1, resulting in amplicons of ~500 bp and 430 bp, respectively. The samples were treated with a commercial enzymatic cleanup reagent, d and submitted for commercial sequencing. e Sequences were edited, trimmed, and assembled using commercial bioinformatics software. f The resulting 689-bp fragment was uploaded to GenBank (accession no. KM357866). The short, first-round polymerase fragment corresponds to nucleotides 299–480 of the Genbank accession.
Basic local alignment search tool nucleotide (BLASTn) analysis of the herpesviral polymerase sequence from the map turtle sample revealed 99.2% sequence identity to EmyHV-1 (KF478668), 77% to terrapene herpesvirus 1 (KJ004665), and 73% to tortoise herpesvirus (AB047545). Given the high sequence identity with EmyHV-1, the molecular characterization of the herpesvirus in affected liver is consistent with a diagnosis of EmyHV-1–associated pneumonia and hepatic and splenic necrosis in this turtle.
Because EmyHV-1 has been reported in the absence of lesions in the eastern river cooter, 12 and transmission of a disease-associated herpesvirus between map turtles and painted turtles has been previously suggested, 11 we were interested in investigating whether EmyHV-1 might be present subclinically in turtles that shared a water system with the dead map turtle. A single, combined choanal and cloacal swab was therefore collected from a total of 27 turtles, including 7 additional northern map turtles, 11 red-eared sliders (Trachemys scripta elegans), 2 spotted turtles (Clemmys guttata), and 7 painted turtles, and tested for herpesvirus. Swabs were extracted using a commercial kita per the manufacturer’s recommendations, and qualitative PCR amplification of the DNA polymerase gene was performed as described above.
Of the additional animals tested, 5 turtles, including a single northern map turtle and 4 painted turtles, were herpesvirus PCR positive. The amplified short, 181-bp polymerase fragments were 99.4–100% identical to each other and to the sample from the deceased map turtle. Modified second-round PCR reactions amplifying a larger, 689-bp fragment of the DNA polymerase gene were also performed as described above for 2 of the 4 painted turtles and the map turtle. Amplicon sequences were 99.9–100% identical to each other and to the deceased map turtle sample. A single, silent nucleotide mutation (T→G; position 431 of accession KM357866) was present in 1 painted turtle and the asymptomatic map turtle.
The deduced amino acid sequence of the partial DNA-dependent DNA polymerase nucleotide sequence was aligned with 59 representatives of the family Herpesviridae using commercial software. f Bayesian phylogenetic analysis was performed with a fixed Poisson rate matrix, gamma distributed rate variation, 4 heated chains, unconstrained branch lengths, and a subsampling frequency of 200 8 ; the first 25% of 1,100,000 chains were discarded as burn-in. The analysis placed both the 689-bp map turtle fragment (boldface, Fig. 2) and the 407-bp EmyHV-1 eastern river cooter fragment (KF478668) within the single, monophyletic Alphaherpesvirinae clade containing Chelonid herpesvirus 5, the type species of the genus Scutavirus, as well as other herpesviruses of freshwater and terrestrial tortoises and turtles (underlined, Fig. 2) and marine turtles (italicized, Fig. 2). 1 These results are consistent with previous phylogenetic studies of EmyHV-1 and other turtle herpesviruses.2,12,19,20

Midpoint-rooted Bayesian phylogenetic tree of predicted amino acid sequences of herpesviral DNA-dependent DNA polymerases. Bayesian posterior probabilities are shown at branch points. The sequence of the virus amplified from the reported map turtle is in boldface. Herpesviruses of freshwater and terrestrial turtles and tortoises are underlined; herpesviruses of marine turtles are italicized. Brackets demarcate the Betaherpesvirinae (β), Gammaherpesvirinae (γ), and Alphaherpesvirinae (α) subfamilies as well as the Alphaherpesvirinae genera Iltovirus, Varicellovirus, Simplexvirus, and Mardivirus. The type virus of the genus Scutavirus, Chelonid herpesvirus 5, is also indicated. GenBank accession numbers are shown in parentheses following the virus name. An asterisk (*) highlights viruses previously published in reference but not present in GenBank. 19
In summary, our study characterizes a fatal herpesvirus infection that caused necrotizing pneumonia and hepatic and splenic necrosis in a northern map turtle. Intralesional, intranuclear, Cowdry type A inclusions were a prominent feature in the pulmonary and hepatic lesions. Aerobic bacterial culture of sections of affected lung yielded growth of both Flavobacterium spp. and C. indologenes. While a concurrent, secondary bacterial infection cannot be excluded, the lesions were not characteristic for an acute, primary bacterial pneumonia; furthermore, as both organisms can be found in water, they may represent postmortem contaminants.
The PCR and sequence analysis identified EmyHV-1, an alphaherpesvirus of turtles, in affected tissue of the dead map turtle and in clinically unaffected painted turtles and a northern map turtle that shared a common circulation system with the dead turtle. These results confirm that EmyHV-1 can cause significant disease in northern map turtles, and suggests that asymptomatic, EmyHV-1–positive painted turtles, and possibly other emydid turtles, such as eastern river cooters, are a possible reservoir for infection. As such, care should be taken managing these species in captivity to limit cross-exposure. Of the 8 map turtles maintained in the system with the EmyHV-1 PCR-positive painted turtles, 2 were EmyHV-1 PCR positive but only a single animal developed clinical disease. Therefore, additional factors such as stress or concurrent infection may be important contributors to the development of herpesviral-associated disease. Excretion and aqueous transmission of herpesviruses is well characterized for some fish pathogens, including Ictalurid herpesvirus 1 and Cyprinid herpesvirus 3,6,14 and we suspect that horizontal transmission through the shared water system or possibly fomites occurred in the current case.
Footnotes
Acknowledgements
We thank the Wildlife Conservation Society’s curatorial and clinical medical staffs for their exceptional care, and Alfred Ngbokoli and Daniel Friedman for slide preparation.
a.
QIAamp DNA mini kit, Qiagen Inc., Valencia, CA.
b.
AmpliTaq Gold 360 master mix, Applied Biosystems, Foster City, CA.
c.
SYBR Safe DNA Gel Stain, Invitrogen Corp., Carlsbad, CA.
d.
ExoSAP-IT, Affymetrix Inc., Cleveland, OH.
e.
Genewiz Inc., South Plainfield, NJ.
f.
Geneious v. 6.1.7, Biomatters Ltd., Auckland, New Zealand.
Declaration of conflicting interests
The author(s) declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.