
Abstract
Whole blood and serum mineral concentrations were measured in diverse bighorn sheep (Ovis canadensis) metapopulations in California, and 90% reference intervals were determined. While there were some statistical differences between median concentrations among the different metapopulations, detected values were generally in good agreement with concentrations reported for other bighorn sheep populations and with reference ranges widely accepted for domestic sheep (Ovis aries). Although median whole blood selenium and serum copper concentrations were within adequate ranges reported for domestic sheep, some metapopulations had substantial numbers of individuals whose concentrations would be considered suboptimal for domestic sheep. There are a number of factors that can influence mineral concentrations in wildlife species such as bighorn sheep and that make the establishment of reference ranges challenging. However, the establishment of mineral reference ranges is important for such species, as their health and productivity are increasingly scrutinized and actively managed.
Introduction
All animals require inorganic elements, or minerals, to carry out normal life processes. 9 The essential mineral elements can be divided into 2 groups based on the amounts needed in the diet. 12 Minerals that are needed in relatively large amounts include calcium, phosphorous, potassium, magnesium, sodium, sulfur, and chlorine and are referred to as macrominerals. Minerals that are needed in small amounts are referred to as microminerals or trace elements.4,9 Such microminerals include chromium, cobalt, copper, fluorine, iodine, iron, manganese, molybdenum, selenium, zinc, and vanadium. Macro- and microminerals can also be referred to as essential minerals because they are required to support adequate growth, reproduction, and health throughout life. Essential minerals act as structural components of body organs and tissues, serve as electrolytes in body fluids and tissues, and act as catalysts in enzyme and endocrine systems. 9 Biochemical defects, altered physiologic functions, and structural disorders occur when tissue concentrations of a mineral become deficient or excessive.9,12 Signs of mineral deficiency or toxicity specific to a given mineral depend on the degree and duration of exposure along with animal variables such as species, age, and sex. 14
One of the most common approaches to assessing antemortem mineral status of animals involves determination of their concentrations in samples such as whole blood or serum. Serum is the preferred sample for determining most essential minerals. However, many laboratories prefer whole blood samples for assessing selenium status because whole blood selenium concentrations reflect both erythrocyte and serum selenium pools. 6 Both whole blood and serum selenium concentrations are reasonably good indicators of nutritional status. 6
Unfortunately, for many wild animal species, reference intervals from apparently healthy feral animals have not been determined, which makes interpretation of detected concentrations for surveillance or disease diagnostic purposes more difficult. For increasingly managed wild animal populations, the development of species-specific reference ranges for a variety of diagnostic parameters is important for assessing population health. Bighorn sheep (Ovis canadensis) populations have declined in a number of states since 1990 (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs: are current population declines caused by malnutrition? Master of Science Thesis, University of Colorado, Boulder, CO). Some investigators have speculated that mineral deficiencies, in particular selenium deficiency, might be playing a role in population declines (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs; Dean R, Hnilicka P, Kreeger T, Delcurto T: 2002, An investigation into the selenium requirements for Rocky Mountain bighorn sheep. Biennial Symposium of the North Wild Sheep and Goat Council 13:95–99). Although reference intervals in whole blood and serum for essential minerals have been established for domestic sheep, limited information is available for bighorn sheep. Thus, laboratories that evaluate mineral concentrations in bighorn sheep from blood and serum samples often use domestic sheep reference intervals. Establishment of mineral concentration reference intervals for healthy bighorn sheep would be helpful in interpretation of mineral concentrations for this species. A number of studies have measured whole blood selenium concentrations in bighorn sheep, but there is a paucity of information concerning other essential minerals more typically measured in serum samples (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs; Coggins VL: 2006, Selenium supplementation, parasite treatment, and management of bighorn sheep at Lostine River, Oregon. Biennial Symposium of the North Wild Sheep and Goat Council 15:98–106; Dean R, et al.: 2002, An investigation into the selenium requirements for Rocky Mountain bighorn sheep).3,10,17,19 The purpose of the current study was to establish trace element reference intervals for several metapopulations of Sierra Nevada bighorn sheep (O. canadensis sierrae) and desert bighorn sheep (O. canadensis nelsoni) in California and report separate intervals for each metapopulation.
Materials and methods
Bighorn sheep populations in California (Fig. 1) have been organized into metapopulation management units (Epps CW, Bleich VC, Wehausen JD, Torres SG: 2003, Status of bighorn sheep in California. Desert Bighorn Council Transactions 47:20–35; Torres SG, Bleich VC, Wehausen JD: 1994, Status of bighorn sheep in California, 1993. Desert Bighorn Council Transactions 38:17–28). Each metapopulation is a system of local populations that are connected by individuals dispersing between them. 5 From 1997 to 2006, free-ranging female bighorn sheep in several metapopulations in California were captured as part of multiple ongoing studies. Bighorn sheep metapopulations sampled for the present study included Northern Mojave (n = 60), Central Mojave (n = 10), Southern Mojave (n = 69), San Gabriel (n = 38), Peninsular (n = 138), Central Sierra (n = 22), and Southern Sierra (n = 51) metapopulations (Fig. 1; Torres SG, et al.: 1994, Status of bighorn sheep in California, 1993). These captures provided an opportunity to collect biological samples to help evaluate the health status of the herds. Most captures occurred from September through December with occasional captures in August or January. Capture after January was generally avoided due to ewes being either in late pregnancy or having recently lambed.
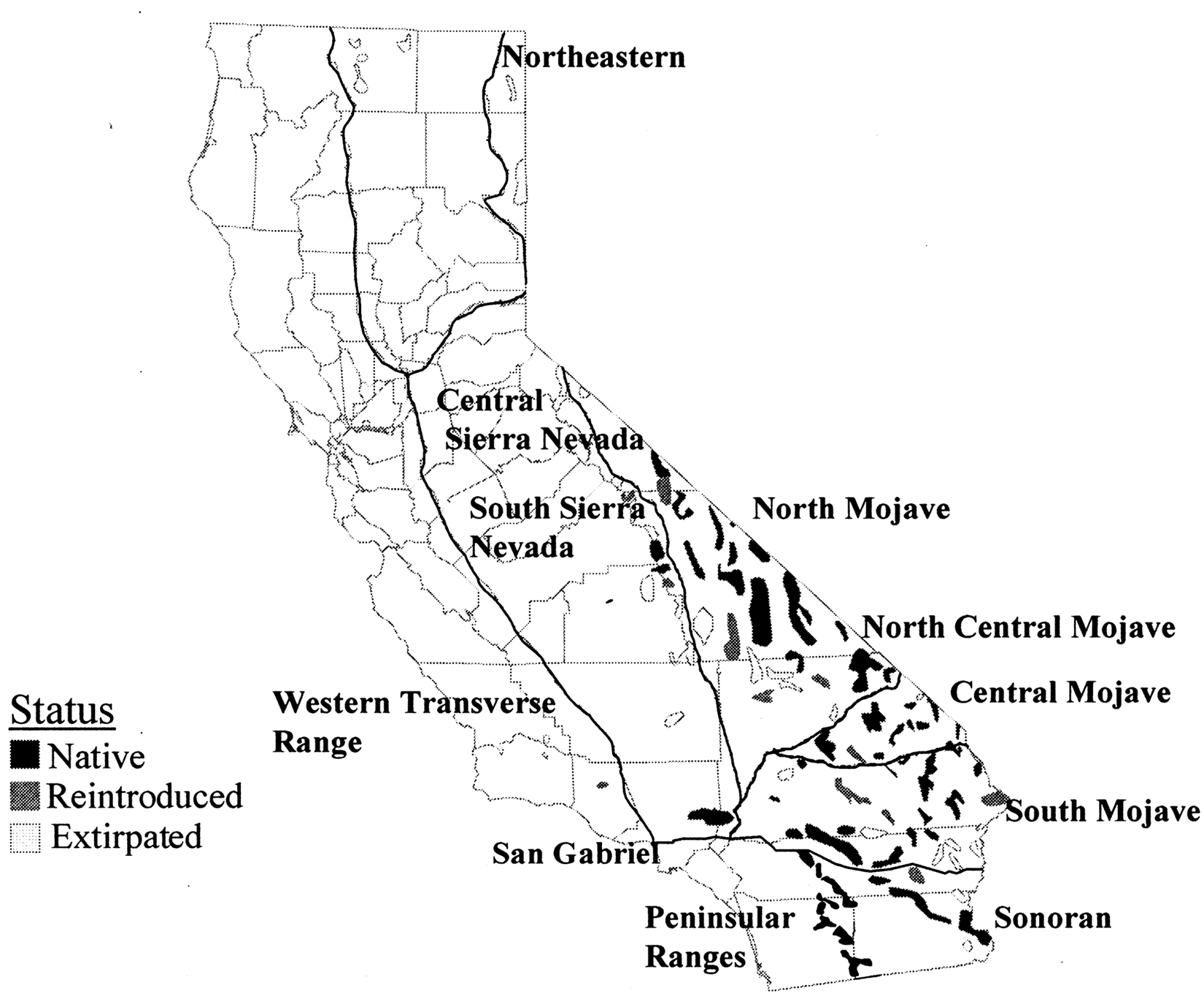
Metapopulations of bighorn sheep in California (Epps et al., 2003).
Captured sheep were examined for evidence of disease, and age was estimated based on tooth wear and horn rings.7,20,23 Animals included in the analysis appeared healthy at the time of capture based on routine physical examinations, although some individual herds within the North Mojave and Peninsular metapopulations had experienced higher than expected lamb mortality and some adult respiratory signs. In addition to determining a number of essential minerals, individuals were screened for Mycoplasma ovipneumoniae, Bovine parainfluenza virus 3, Bovine respiratory syncytial virus, Bluetongue virus, Epizootic hemorrhagic disease virus, Brucella ovis, Bovine herpesvirus 1, Orf virus, chlamydia, and Bovine viral diarrhea virus 1 and 2. Because some essential minerals (chromium, cobalt, fluorine, iodine, molybdenum, and vanadium) are not included in routine mineral screens, they were not measured in the current study. Because of missing data, not every animal was included in the analysis of each mineral, age, or sex stratification.
For purposes of the current study, blood was collected by jugular venipuncture from each bighorn sheep using an 18-gauge, 1-inch needle and 60-ml syringe. Approximately 6 ml of blood was placed in a trace element collection tube a and cooled on ice. The samples were centrifuged at 1,500– 2,000 × g for 10–15 min the day they were collected, and the serum was placed into a new trace element collection tube. Approximately 3 ml of blood was placed in a glass blood collection tube containing 4.5 mg of powdered Na2–ethylenediamine tetra-acetic acid (EDTA). b All samples were kept on ice or refrigerated until they were delivered to the California Animal Health and Food Safety Laboratory (CAHFS) at the University of California at Davis for analysis.
Selenium analysis was performed by inductively coupled plasma–atomic emission spectrometry (ICP-AES) after elemental selenium in the sample was reduced to hydrogen selenide. Briefly, whole blood was digested overnight with a mixture of sulfuric, nitric, and perchloric acids to oxidize all forms of selenium to selenate. The selenate was then reduced to selenite in hydrochloric acid at 95°C. Selenite was then quantitatively reduced to hydrogen selenide by acidic sodium borohydride, which was added into the stream of the sample as it was pumped into the nebulizer of the ICP-AES. The reporting limit for whole blood selenium was 0.010 ppm.
Other essential mineral concentrations (iron, magnesium, zinc, copper, calcium, inorganic phosphorus, sodium, and potassium) were measured in serum. One-half milliliter of serum was placed in a test tube to which 4 ml of a protein precipitating internal standard solution were added. The sample was mixed and centrifuged to produce a clear supernatant free of proteins. The sample was analyzed for the listed elements by ICP-AES. All results were quantified and reported in ppm, except sodium and potassium, which were reported in milliequivalents per liter. The reporting limits were: iron (0.04 ppm), magnesium (2 ppm), zinc (0.1 ppm), copper (0.1 ppm), calcium (4 ppm), phosphorus (2 ppm), sodium (4 mEq/l), and potassium (0.2 mEq/l).
Selenium and other essential minerals were quantitated by comparison with external calibration solutions. The quality of the analytical run was controlled through the use of Standard Reference Materials (SRMs) containing certified concentrations of the analytes and matrix matched blank samples. Method validation followed a CAHFS-approved validation protocol.
Standard descriptive statistics c were derived for the concentrations of each mineral for the overall population and for each metapopulation separately. Normality of the distribution of each mineral concentration was assessed. Values that were more than 3 standard deviations (SDs) above or below the mean were considered to be outliers and were eliminated from statistical analysis. Because element concentrations did not follow a Gaussian distribution, nonparametric statistical methods were used to compare groups. The Mann–Whitney Test was used to look for significant differences in concentrations of each element between juvenile animals (≤2 years old) and adults (>2 years old) and between males and females within each population. The Kruskal–Wallis test was used to determine whether concentrations differed by metapopulation for each mineral, and the Mann–Whitney test was used to make pairwise comparisons to determine which metapopulations had element concentrations significantly different from one another. P values less than 0.05 were considered significant for all tests. The nonparametric method was used to determine 90% reference limits for each element within each metapopulation. 21
Results
Reference intervals for mineral concentrations in whole blood and serum for 7 free-ranging bighorn sheep metapopulations are presented in Table 1. Selenium concentrations varied the most between metapopulations. Eleven out of 38 sheep from the San Gabriel metapopulation and 5 out of 52 sheep from the South Sierra metapopulation would be considered deficient based on a previously published source. 13 Using a higher limit for a minimally adequate selenium concentration of 0.12 ppm from another publication, 6 the number of individuals falling into a deficient range for the San Gabriel and South Sierra metapopulations increase to 20 each.
Reference ranges (90% nonparametric intervals) for essential mineral concentrations in whole blood (selenium) and serum of bighorn sheep (Ovis canadensis) from 7 metapopulations in California.
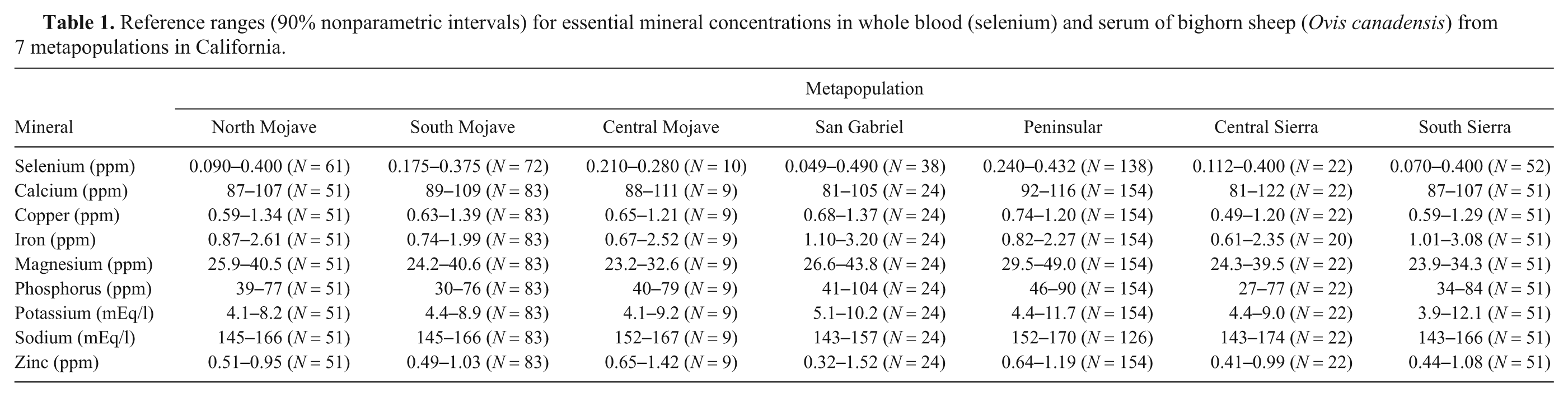
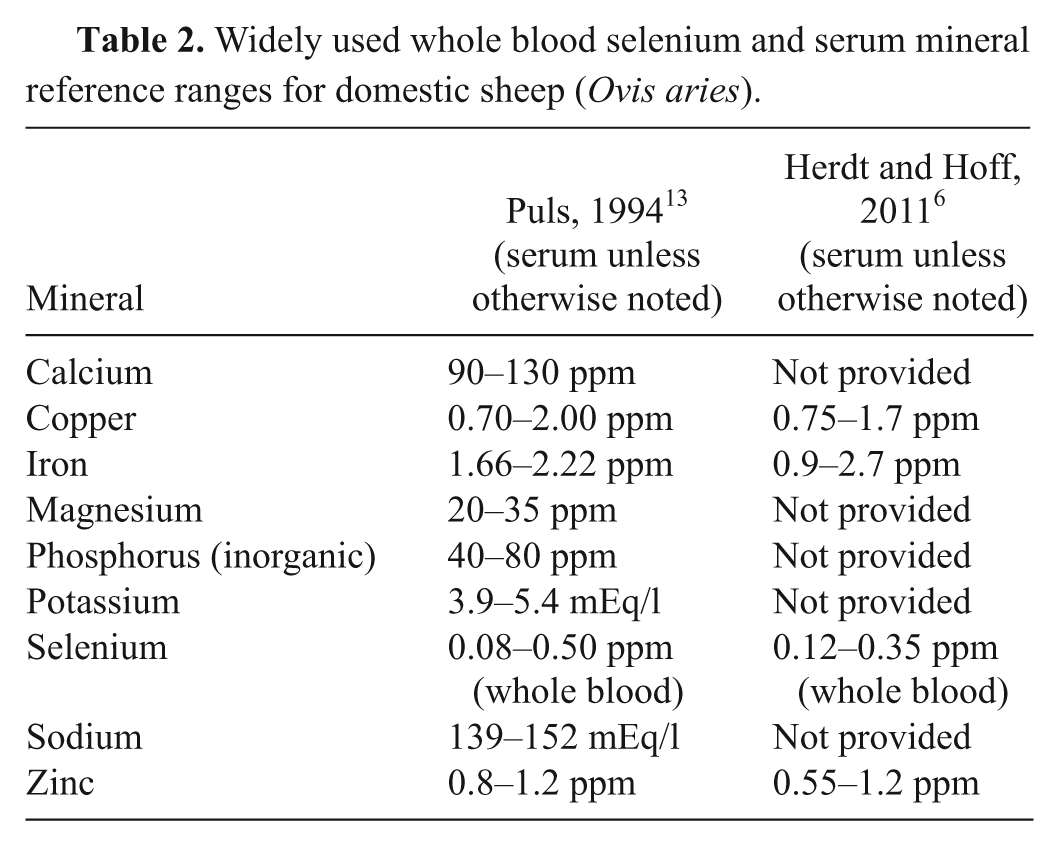
In general, serum calcium, copper, magnesium, inorganic phosphorus, potassium, and zinc concentrations were within published reference ranges for domestic sheep (Table 2), although some individuals fell within ranges that would be considered suboptimal.6,13 Specifically, serum copper concentrations below a range considered to be adequate for domestic sheep according to a widely referenced source 13 (0.70–2.0 ppm) were noted for 17.6% (9/51) of the North Mojave, 9.6% (8/83) of the South Mojave, 11% (1/9) of the Central Mojave, 4.2% (1/24) of the San Gabriel, 2.6% (4/154) of the Peninsular, 32% (7/22) of the Central Sierra, and 27% (14/51) of the Southern Sierra metapopulations. These percentages increased somewhat for most metapopulations when using a higher lower limit of adequate (0.75 ppm) provided by another source 6 (13/51 or 25.5% for North Mojave, 13/83 or 15.7% for South Mojave, 2/9 or 22.2% for Central Mojave, 7/154 or 4.5% for Peninsular, and 18/51 or 35.3% for Southern Sierra). While many individual serum iron and zinc concentrations were below adequate ranges for domestic sheep as reported in a previous published source, 13 the concentrations were within ranges typical for the CAHFS laboratory (unpublished information) and in better agreement with the lower limits of adequate for serum iron and zinc provided by others. 6
Widely used whole blood selenium and serum mineral reference ranges for domestic sheep (Ovis aries).
Discussion
Whole blood selenium and serum mineral reference ranges are available for major domestic ruminant species.6,13 Unfortunately, reference ranges are not available for comparable wild ruminant species. Such ranges can be used to help diagnose mineral deficiencies or intoxications. However, mineral concentrations that fall outside of reference ranges need to be evaluated in conjunction with other clinical and diagnostic information to reach a diagnosis because there is substantial variation among species and individuals. In general, the mineral concentrations detected for the bighorn sheep metapopulations in the current study fell within those widely used for domestic sheep. Interlaboratory variation in analytical procedures can make the general applicability of published reference ranges problematic. For example, the CAHFS laboratory tends to have serum iron and zinc concentrations that are consistently below adequate ranges reported by a widely cited source 13 for a variety of species, and CAHFS-specific reference ranges have not been determined for domestic sheep. The differences in reference ranges between those from 2 sources for whole blood selenium, serum copper, and serum zinc illustrate discrepancies between such ranges.6,13 Ranges were established either from a review of the literature 13 or were derived from a single diagnostic laboratory using a specific mineral screening methodology (inductively coupled plasma–mass spectrometry) that has become more commonly used in recent years for diagnostic purposes. 6
It is important to point out that there are reported overlaps between what would be considered adequate, marginally adequate, and deficient serum concentrations for some minerals according to one source. 13 For example, in domestic sheep, serum copper concentrations of 0.10–1.00 ppm, 0.40–1.00 ppm, and 0.70–2.00 ppm are considered deficient, marginal, and adequate, respectively.
Another potential issue when comparing mineral concentration ranges established for domestic species to similar wildlife species relates to the possibility that wildlife species have developed adaptive mechanisms that make them less susceptible to mineral aberrations, especially deficiencies. 19 Bighorn sheep evolved in granitic and glaciated environments that are characteristically low in minerals such as selenium and copper, and it has been postulated that daily dietary mineral requirements for wildlife ruminant species differ from those for domestic ruminants.2,15,24
Seasonal variations in trace element concentrations are likely to occur due to variations in mineral intake, which is an additional complication when reference ranges are derived from a single sampling period. Indeed, serial mineral data collected from a relatively small number of individual bighorn sheep at different times of the year showed substantial variation, particularly for selenium. 3 Movement from summer to winter habitats alters mineral intake due to differences in plant mineral content (Coggins VL: 2006, Selenium supplementation, parasite treatment, and management of bighorn sheep; Dean R, et al.: 2002, An investigation into the selenium requirements for Rocky Mountain bighorn sheep). Forage and fecal pellet mineral concentrations were higher in montane soils in Colorado bighorn sheep winter ranges compared to alpine soils in summer ranges (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs). The habitats used by the sampled metapopulations in the present study are diverse and range from desert scrub and desert wash communities at lower elevations to alpine meadows and mixed conifer and lodgepole pine communities at higher elevations (Holl SA, Bleich VC: 1983, San Gabriel Mountain sheep: biological and management considerations. USDA Forest Service, unpublished report on file at San Bernardino National Forest, San Bernardino, CA).1,18,22 In California, many bighorn sheep do not utilize separate summer and winter ranges (Ben Gonzales, personal communication, 2011) although several populations do migrate to different altitudes based on season.
The selenium status of wild populations of bighorn sheep has been studied the most extensively (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs; Coggins VL: 2006, Selenium supplementation, parasite treatment, and management of bighorn sheep; Dean R, et al.: 2002, An investigation into the selenium requirements for Rocky Mountain bighorn sheep).3,17,19 Selenium is a key component of the antioxidant enzyme glutathione peroxidase and, along with vitamin E, is important for preventing muscular dystrophies, ill-thrift, poor reproduction, impaired immunity, and bone marrow abnormalities in domestic sheep.14,16 Concentrations as low as 0.01 ppb (0.00001 ppm) were noted in one study, with the highest concentration reported in sampled bighorn sheep of 11.76 ppb (0.0117 ppm). 3 These values are extremely low, as a comparable reference range for domestic sheep is 80–500 ppb. 13 Extremely low concentrations were also reported for other minerals such as copper, iron, and zinc. Few analytical details were provided by the authors, and it is possible that units (µg L–1 or ppb) were reported incorrectly.
Mean ± SD whole blood selenium concentrations reported for 2 bighorn sheep populations in Alberta, Canada, were 0.025 ± 0.011 ppm (N = 51; range: 0.005–0.045 ppm) and 0.120 ± 0.030 ppm (N = 8; range: 0.070–0.160 ppm). 19 Another study reported mean ± SD whole blood selenium concentrations for 2 populations of bighorn sheep in Colorado of 0.17 ± 0.05 ppm (N = 30; range: 0.07–0.27 ppm) and 0.17 ± 0.05 ppm (N = 8; range: 0.11–0.27 ppm; Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs). A third study reported selenium population means (± SD) for bighorn sheep populations in Oregon, Washington, and Idaho from 0.09 ± 0.06 ppm (Asotin, WA; N = 13) to 0.39 ± 0.15 ppm (Lostine, OR; N = 116; Coggins VL: 2006, Selenium supplementation, parasite treatment, and management of bighorn sheep). The Lostine, Oregon, population received supplemental selenium in the form of selenized salt. Interestingly, the bighorn sheep population in Alberta that had the lowest mean whole blood selenium concentration also had what was considered to be high lamb production, younger ages to first estrus, overall high flock survival, and long life spans. 19 Selenium concentrations detected for the California metapopulations were well above the concentrations described for the Alberta herd.
Data specific to bighorn sheep are limited for other trace elements such as copper, iron, and zinc. Aside from the questionable results from a previously published study, 3 the only data available for other trace elements are those reported for Rocky Mountain bighorn sheep (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs). Mean ± SD for 2 populations of bighorn sheep for copper, iron, and zinc were 0.68 ± 0.29 ppm (range: 0.18–1.31 ppm) and 0.63 ± 0.24 ppm (range: 0.49–1.24 ppm), 1.77 ± 0.4 ppm (range: 0.98–2.37 ppm) and 1.91 ± 0.56 ppm (range: 0.6–2.43 ppm), and 0.61 ± 0.14 ppm (range: 0.4–1.03 ppm) and 0.67 ± 0.23 ppm (range: 0.43–1.21 ppm), respectively (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs). The serum mineral concentrations determined for the California metapopulations appear to be comparable. In domestic sheep, serum copper, iron, and zinc concentrations considered to be marginal or deficient by a widely used source are 0.40–1.00 ppm (marginal) and 0.10–1.00 ppm (deficient) for copper, <1.60 ppm (deficient) for iron, and 0.40–0.80 ppm (marginal) and 0.22–0.45 ppm (deficient) for zinc. 13 Copper deficiency in domestic sheep is associated with neurological deficits in lambs (enzootic ataxia or swayback), anemia, reduced immune response, increased susceptibility to infectious and parasitic diseases, poor growth rate, poor wool pigmentation and length, reduced fertility, abortions, and stillbirths.13,14 Iron deficiency is associated with anemia, and zinc deficiency is associated with weak hoof horn, enlarged joints, ankylosed hocks and knees, reduced reproductive performance, impaired immune function, skin abnormalities, wool loss, anemia, and weight loss. 13 It is important to point out that deficiencies of more than 1 mineral can have combined adverse effects on reproduction and immune function.
Differences in trace element concentrations between bighorn sheep metapopulations are consistent with the many factors that can affect regulation of mineral elements, such as forage and soil components, rumen microflora, interactions among the various trace elements, breed or genetic effects, physiological state, parasites, toxicant exposure, and immune interactions. 8 Metapopulations in close geographic proximity did, in some cases, have significantly different concentrations of trace elements (data not reported). The habitats of sheep in each metapopulation are quite diverse. Mineral content of the soil and, secondarily, vegetation is likely to differ extensively across the habitats used by these populations. For example, selenium content of forages and grain varies across California. 11 As much as 80% of forages and grain in some areas contain less than 0.1 ppm selenium on a dry matter basis, which is a concentration considered minimally sufficient to prevent deficiency in domestic sheep.11,14 For this reason, domestic sheep in such areas are supplemented with selenium. The effect of low selenium forages on bighorn sheep is not clearly understood, and it is possible that bighorn sheep can tolerate lower dietary mineral intakes than domestic sheep. There is limited evidence that mineral supplementation can affect bighorn sheep productivity. One study reported increased lamb recruitment in Colorado flocks supplemented with minerals (Carpenter LM: 2005, The role of trace nutrients in the survival of bighorn sheep lambs). In contrast, selenium supplementation of an intensively managed bighorn sheep population in Oregon did not appear to prevent low lamb survival or the occurrence of disease outbreaks (Coggins VL: 2006, Selenium supplementation, parasite treatment, and management of bighorn sheep).
Although the effects of essential mineral deficiencies on the health of domestic sheep have been well documented, this information might not be applicable to bighorn sheep. Toxic accumulation of mineral elements is likely rare in bighorn sheep as it is in domestic sheep, with the exception of copper. 8 However, marginal deficiencies that could prevent optimal health and reproduction might occur but are more difficult to detect. Establishment of reference intervals for bighorn sheep provides baseline information for monitoring the impacts of various mineral disturbances in these metapopulations, and data provided herein might be useful for evaluation and management of herd productivity. Long-term disease screening, in combination with health monitoring of bighorn sheep herds, will be useful to advancing the understanding of the population and the individual animal significance of these reported mineral concentrations.
Footnotes
a.
Vacutainer® Trace Element Serum Plus Blood Collection Tube, BD, Franklin Lakes, NJ.
b.
Monoject®, Sherwood Medical, St. Louis, MO.
c.
SPSS Base 11.0 for Windows User’s Guide, SPSS Inc., Chicago, IL.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
This work was funded through the California Department of Fish and Game.