
Abstract
Between 2006 and 2008, 4 captive Asian horned frogs (Megophrys montana) were diagnosed with ocular and neurologic disease associated with rhabditid nematodiasis. Mortality, either spontaneous or by humane euthanasia, was high (3/4, 75%). Gross and histologic findings included varying degrees of ulcerative keratitis, histiocytic uveitis and retinitis, meningoencephalomyelitis, and epidermal chromatophore (iridophore) hyperplasia with intralesional nematodes. Entry into the host was presumed to be by direct invasion of the skin and the cornea with migration through the optic nerve to the brain and spinal cord. One frog was diagnosed with rhabditid nematodiasis antemortem, and clinical signs and lesions in the frog did not progress after unilateral enucleation and anthelminthic treatment were completed. Gross and tissue morphology of the nematodes were consistent with the order Rhabditida. DNA was extracted separately from 2 individual nematodes that were isolated from frozen and ethanol-preserved eye and brain tissue. These DNA templates were used for polymerase chain reaction amplification and sequencing of nuclear 28S large subunit (LSU) and internal transcribed spacer (ITS) ribosomal DNA regions. Comparison of the LSU and ITS sequences to those deposited in GenBank revealed an exact match for Caenorhabditis elegans.
Infection with nematodes in the order Rhabditida, and specifically, the genera Rhabdias and Strongyloides are commonly observed in captive and free-ranging amphibians. 2,6,15 In low numbers, these rhabditid nematodes cause little impact and are restricted to the lung and intestine, respectively. In high numbers, the nematodes can cause disease due to aberrant migration, protein-losing enteropathy, or secondary bacterial infections and sepsis. These “superinfections” are often seen in confinement where amplification of the direct life cycle can result in heavy environmental loads of infective larvae and repeated infections of susceptible hosts. 15 The result can be devastating, causing significant morbidity and mortality in captive amphibians. 6,14,15 For example, a Strongyloides sp. infection resulted in death of a captive Cope's gray treefrog (Hyla chrysoscelis) due to enteritis. 12 Experimental infection with high numbers of Rhabdias sphaerocephala larvae resulted in the death of juvenile giant toads (Bufo marinus) due to cutaneous and visceral larval migrans. 20 And lastly, superinfection with rhabditid nematodes, Rhabdias sp. and Strongyloides sp., has been recognized as a threat to amphibian conservation efforts, both to wild-caught amphibians in captive assurance colonies and amphibians in captive breeding programs. 14
Ocular nematodiasis in vertebrates can occur as primary ocular parasitism or as an extension of systemic migration. 8,18 The classic example of ocular and neural rhabditoid nematodiasis in veterinary medicine is Halicephalobus gingivalis in horses, both as an extension of visceral larval migrans and potentially as the primary site of infection. 9,16,17 Thelazia spp. are primary parasites of the conjunctiva and lacrimal ducts in mammals. Some Habronema spp. and Onchocerca cervicalis cause disease in the conjunctiva and sclera of horses, dogs, and other mammals. Dipetalonema reconditum, Toxocara canis, Angiostrongylus vasorum, Dirofilaria immitis, and Baylisascaris procyonis can extend to the conjunctiva, to the eyelid, or into the eye opportunistically, as an extension of larval migration. 19 An unidentified spirurid nematode caused conjunctivitis and dacryoadenitis in 2 tiger salamanders (Ambystoma tigrinum). 7 A single case of blindness due to unidentified nematode larvae was briefly referenced in a review of the amphibian eye. 8 The only helminth parasite that is specifically intraocular is the larval stage (metacercaria) of the fluke Diplostomum spathaceum, which principally colonizes the lenses of fish. 19 The current report documents ocular and neural rhabditid nematodiasis in 4 captive Asian horned frogs (Megophrys montana).
Between 2006 and 2008, 4 Asian horned frogs were presented to the Department of Veterinary Services at the San Diego Zoo (San Diego, CA) for corneal opacity or corneal ulceration of 2–12 months duration. In 2 of the frogs, the lesions failed to respond to topical or systemic antibiotics, corticosteroids, and/or nonsteroidal anti-inflammatory drugs, and they rapidly progressed to perforation. Frog no. 1 developed progressive ataxia and paresis and was euthanized. Frog no. 2 was treated successfully with surgical enucleation of the affected eye and anthelminthic treatment. Frog no. 3 was humanely euthanized for poor response to therapy. Frog no. 4 was found dead with bilateral corneal opacities. The frogs had been housed alone or in pairs in indoor exhibits that contained sphagnum moss substrate, potted plants, and a shallow water dish. The frogs were fed ad libitum, 3 times a week, on a diet consisting of approximately 85% domestic crickets and 15% larvae of black soldier flies (phoenix worms), wax moths (wax worms), or beetles (king worms).

Asian horned frog (Megophrys montana), external appearance. Multiple, often coalescing, slightly raised yellow plaques (arrowhead) are on the dorsal and appendicular skin. The normal coloration of Asian-horned frogs is brown with darker brown markings.
The frogs were submitted to the Wildlife Disease Laboratories at the San Diego Zoo for necropsy. The surgically enucleated eye was submitted for biopsy evaluation. On gross necropsy, the frogs were thin based on a lack of significant coelomic fat stores and had focal to multifocal and coalescing raised yellow plaques on the skin of the head, appendages, and dorsum (Fig. 1). One or both corneas of affected frogs were diffusely opaque, and on 1 or both corneas there were discrete, yellow-grey, dull, raised plaques, rimmed by hemorrhage (Fig. 2). The plaques extended through the entire thickness of the cornea. Similar findings were present in the enucleated eye. A complete set of tissue samples were collected in 10% buffered formalin, routinely processed, and embedded in paraffin blocks.
From these blocks, 4- to 6-μm thick sections were cut and stained with hematoxylin and eosin. Triplicate samples of eye, brain, and skin were kept in formalin and 95% ethanol, and frozen at −80°C for further ancillary diagnostics as necessary. Samples of moss substrate from 2 frog enclosures were examined by light microscopy in a wet mount. Additional samples of used and unused heat-sterilized substrate were preserved in 95% ethanol.
Histopathologic findings were largely limited to the eyes, brain, spinal cord, and skin, and they confirmed the severity of disease. Ocular changes were characterized by chronic corneal ulceration with histiocytic to granulomatous keratitis, corneal edema and fibrosis, histiocytic to granulocytic uveitis and chorioretinitis, hyphema, and intralesional nematodes (Fig. 3). Frog nos. 3 and 4 also had cataractous changes in their lenses and retinal atrophy, which together were suggestive of glaucoma. Frog nos. 1 and 4 had acute meningoencephalomyelitis and parenchymal necrosis with intralesional nematodes (Fig. 4). Examination of the yellow plaques seen on all frogs revealed mild to severe multifocal epidermal iridophore hyperplasia (Fig. 5). Rare nematodes within the pancreas, liver, kidney, and skin were surrounded by mononuclear infiltrates and encapsulated by fibrous connective tissue with frequent mineralization. Mild membranoproliferative glomerulonephritis, chronic interstitial nephritis, melanomacrophage hyperplasia, and adrenocorticocyte vacuolation were common incidental findings.
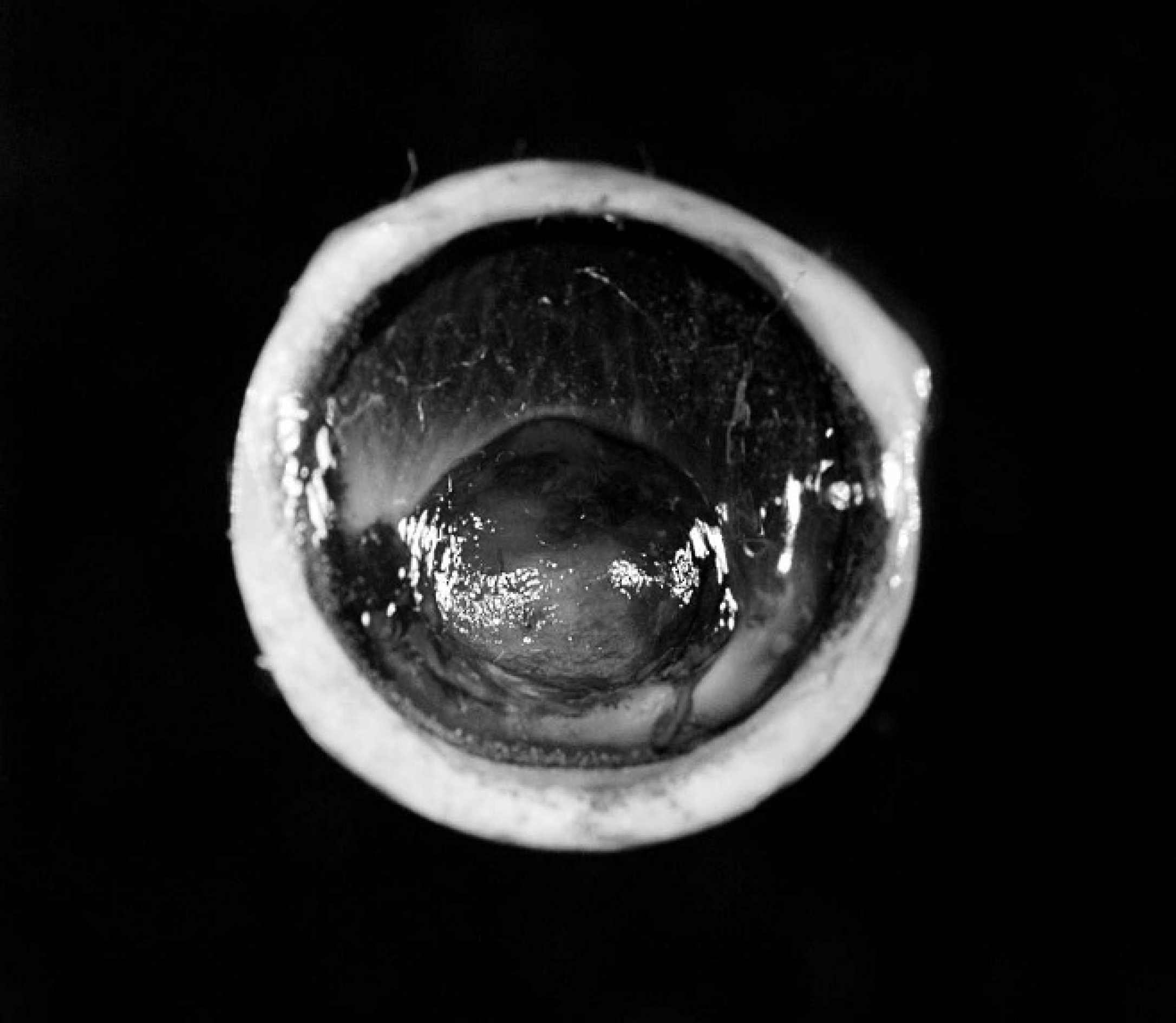
Asian horned frog (Megophrys montana), enucleated eye. Paracentral corneal ulcer covered by a mat of fibrin and hemorrhage and surrounded by corneal edema and hyperpigmentation.
The nematodes were small (25 mm in diameter) with a thin cuticle, platymyarian to inapparent musculature, and a small to medium-sized intestinal tract lined by uninucleated cells (Fig. 6 inset). Both larvae and adult nematodes were present (Fig. 6). In occasional longitudinal sections, a tubular stoma, isthmus, and basal esophageal bulb, characteristic of many nematodes in the order Rhabditida, were identifiable. 5 The rare encapsulated nematodes were similar in size but were degenerate, and their internal anatomy could not be compared to the nematodes from the eye and brain. Varieties of nematodes, including rhabditids of similar overall morphology to those infecting the frogs, were identified in a wet mount of used substrate from a frog enclosure when examined by light microscopy. Nematodes were not identified in a wet mount of unused substrate. The frozen tissue was submitted to the Department of Nematology at the University of California, Davis, for nematode identification. Based on the gross and histopathologic findings, other ancillary diagnostic tests were not pursued.
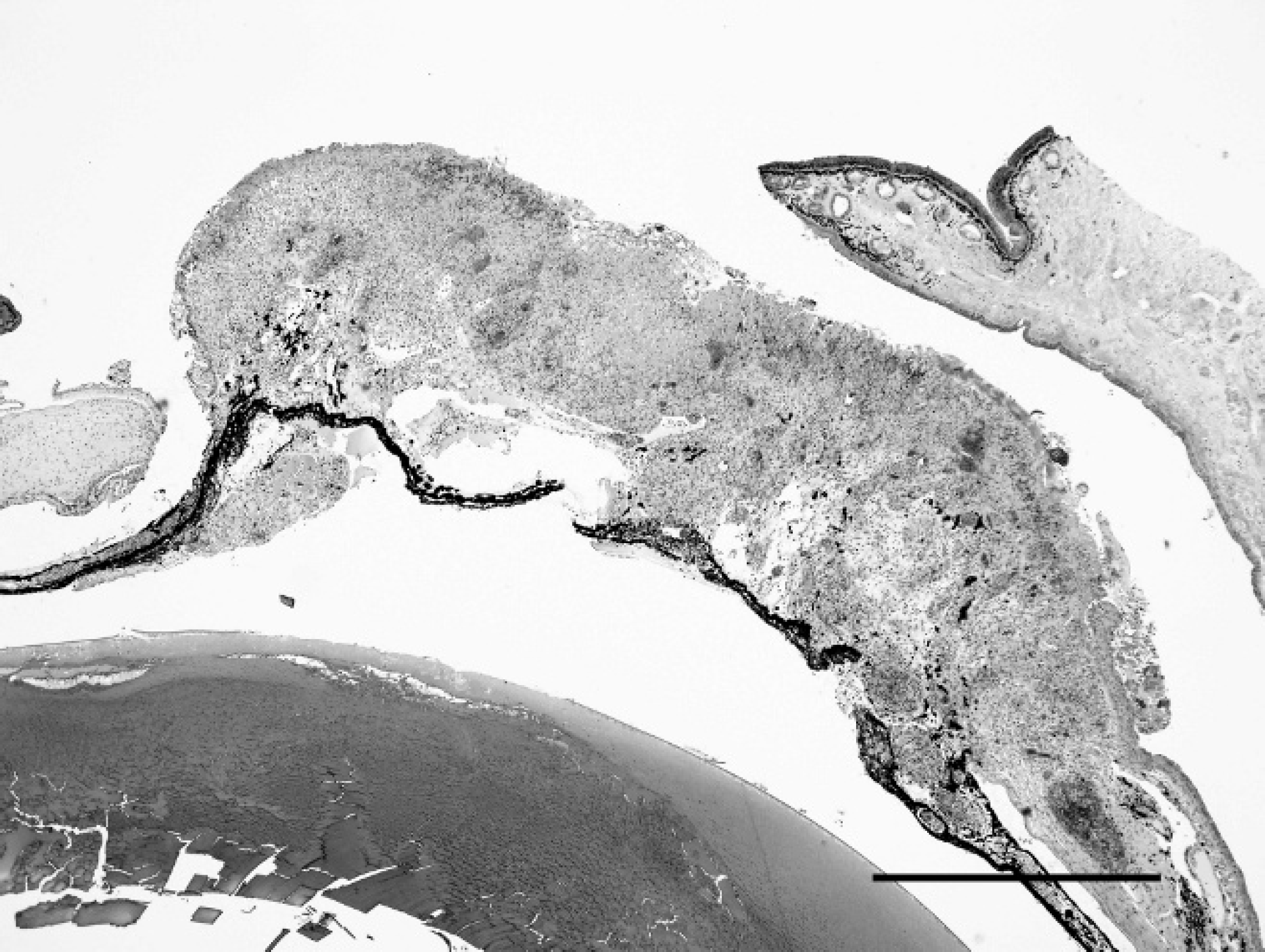
Asian horned frog (Megophrys montana), enucleated eye. Segmental expansion of the cornea with ulceration, focal perforation, and entrapment of the iris. Hematoxylin and eosin. Bar 5 1 μm.
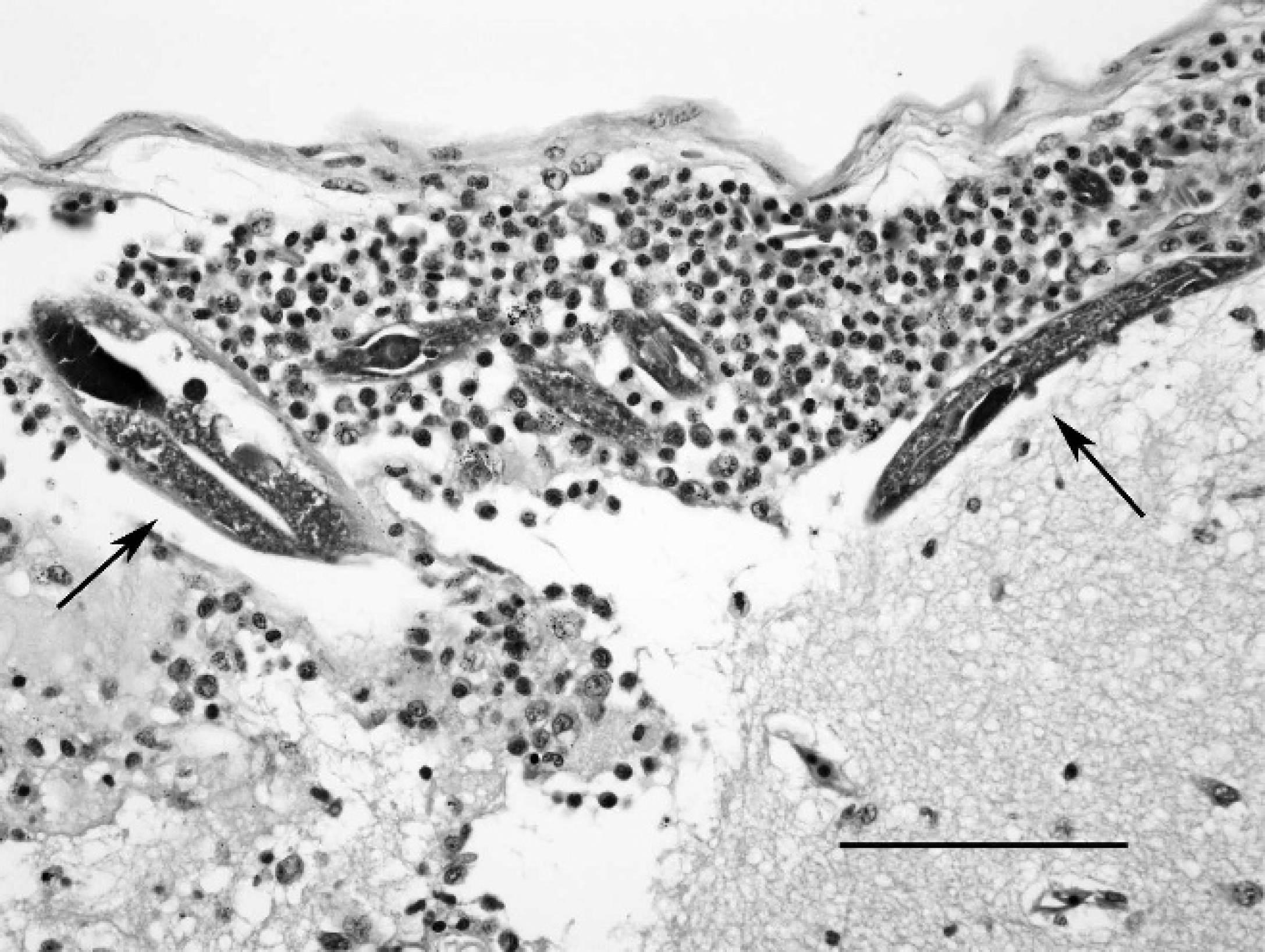
Asian horned frog (Megophrys montana), cerebral cortex and meninges. Multiple small nematodes (arrows) admixed with histiocytes, few granulocytes, and fibrin in the subarachnoid space. Hematoxylin and eosin. Bar = 100 μm.
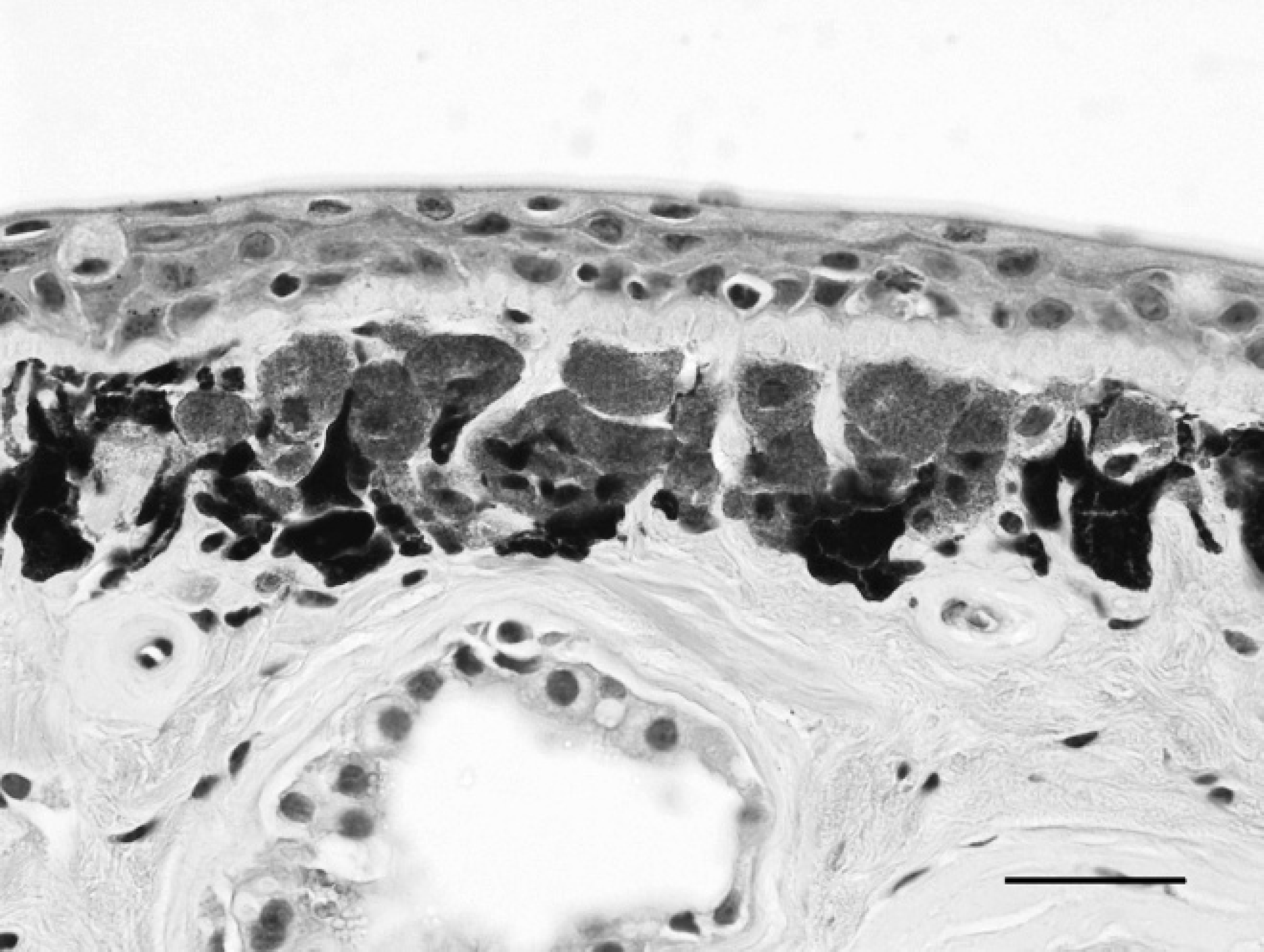
Asian horned frog (Megophrys montana), dorsal skin. Marked chromatophore (iridophore) hyperplasia in the superficial dermis. Hematoxylin and eosin. Bar = 20 μm.
Two individual nematodes were dissected from frozen and ethanol-preserved eye and brain tissue from one frog and digested for DNA extraction using the sodium hydroxide method. 4 Nucleic acids from the individual nematodes (1.5 μl of extract) were used separately for polymerase chain reaction (PCR) amplification. A region of the 28S large subunit (LSU) nuclear ribosomal DNA (rDNA) gene, including the D2/D3 divergent domains, was amplified using primer no. 500 (5′ -ACTTTGAAGAGA-GAGTTCAAGAG) and primer no. 501 (5′-TCG-GAAGGAACCAGCTACTA). Similarly, a region including the nuclear ribosomal internal transcribed spacers (ITS1, ITS2) and 5.8S gene was amplified using primer no. 93 (5′-TTGAACCGGGTAAAAGTCG) and primer no. 94 (5′-TTAGTTTCTTTTCCTCCGCT), which anneal within the small subunit (SSU; no. 93) and LSU (no. 94) rDNA. The PCR amplification reactions (25 μl) consisted of 0.5 μmol of each primer, 200 μmol deoxynucleoside triphosphates, and 3 mmol MgCl2. Proofreading polymerase (0.5 units) a was used for amplification. The PCR cycling parameters included denaturation at 94°C for 3 min, followed by 35 cycles of 94°C for 30 sec, 54°C for 30 sec, and 72°C for 1 min, with a postamplification extension at 72°C for 7 min. The PCR products were separated by agarose gel electrophoresis and visualized using ethidium bromide to confirm product size and yield. The PCR products were prepared for direct sequencing using enzymatic treatment with exonuclease I and shrimp alkaline phosphatase (PCR product presequencing kit b ). Sequencing reactions were performed using dye-terminator sequencing chemistry, c and reaction products were separated and detected using a capillary DNA sequencer. c Sequences were obtained using the PCR primers and internal primers as required, yielding 546 base pair (bp) of double-stranded sequence for the LSU region, and 1,005 bp for the ITS1/5.8S/ITS2 region. CodonCode Aligner (version 1.5.1) d and Phred base calling e were used for assembly of contigs. Sequences produced during the present study have been deposited in GenBank (accession nos. FJ589007-FJ589008 [ITS rDNA] and FJ589009-FJ589010 [LSU rDNA]).
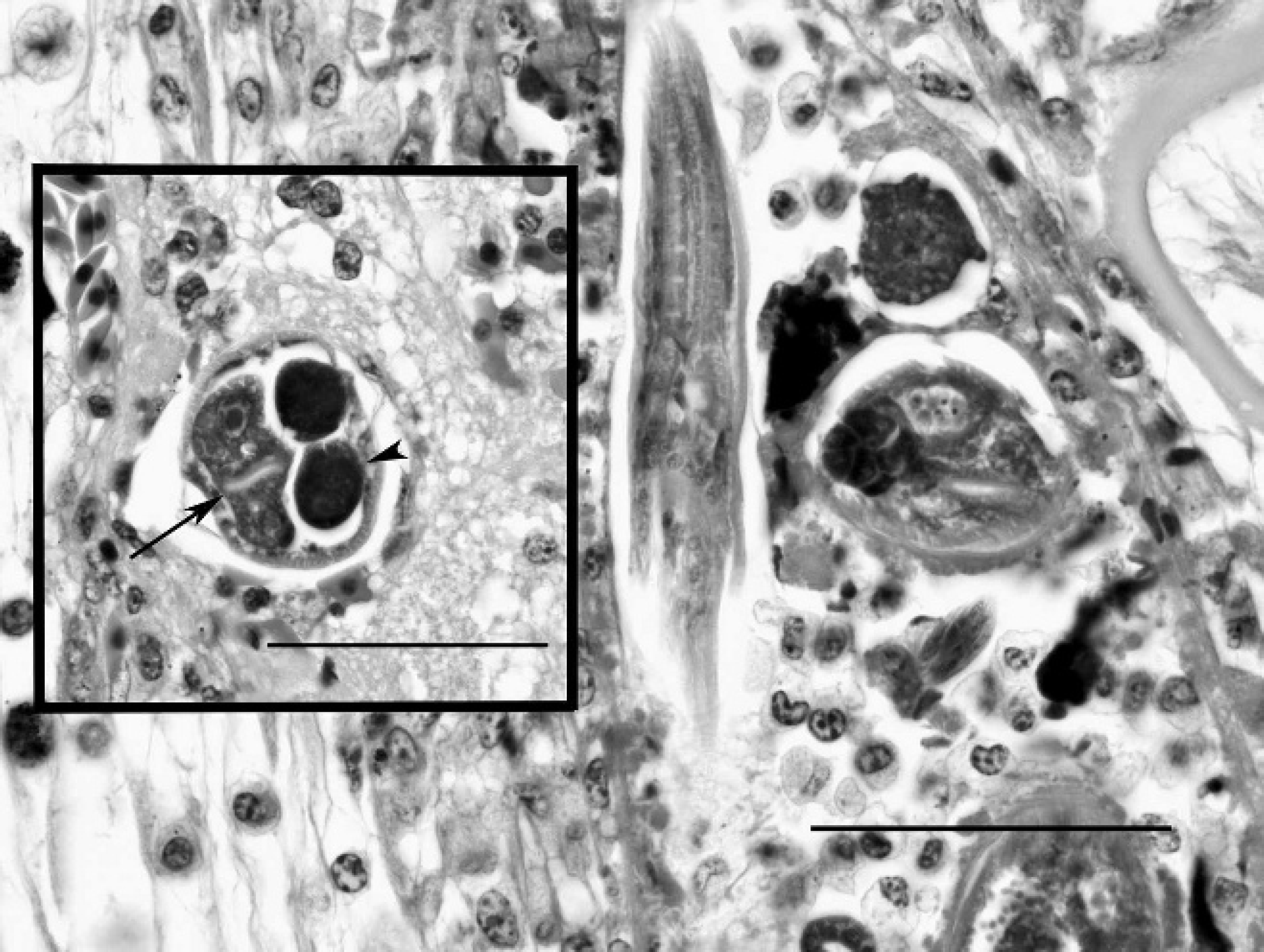
Asian horned frog (Megophrys montana), cornea. Larvae and adult nematodes surrounded by histiocytes and granulocytes in the posterior cornea. Both transverse and longitudinal profiles demonstrate characteristic features of the nematodes, including the tubular stoma and isthmus. Hematoxylin and eosin. Bar = 50 μm. Inset: Asian horned frog, spinal cord. Adult female nematode with low platymyarian musculature, an intestinal tract (arrow) lined by large uninucleated cells, and a single genital tract (arrowhead), containing large ova, that is dorsally retroflexed at the caudal pole. Hematoxylin and eosin. Bar = 50 μm.
The LSU and ITS ribosomal sequences from the 2 nematodes isolated from one of the affected frogs had absolute similarity (100%) to the Gen Bank sequences for Caenorhabditis elegans. 3 These rDNA regions have shown divergence among sequenced Caenorhabditis species, and the LSU rDNA sequence, among other rDNA sequences, has been used previously to create a phylogenetic tree demonstrating relationships among 10 Caenorhabditis species. 11 Based on this phylogenetic tree and the LSU region sequenced for the frog Caenorhabditis, the nematodes isolated from the affected frogs were distinct from the most closely related Caenorhabditis species (C. briggsae, C. remanei, and Caenorhabditis sp. CB5161) by 22–25 differences in pairwise comparisons. It remains possible that the nematodes are not C. elegans, but are a very closely related and undescribed Caenorhabditis species. However, for this to be true, the species would have to be so recently divergent from C. elegans as to have identical rDNA sequences. Thus, the sequence data strongly suggests that C. elegans was responsible for the infection of these frogs.
Caenorhabditis elegans is a rhabditid nematode used widely in biological research as a model organism. The complete sequence of its genome is available, and much is known about the cellular biology, neurobiology, and development of this nematode; however, data on its natural history and normal habitat are currently speculative. Though C. elegans and other Caenorhabditis species can be found in nutrient-rich organic matter, such as garden compost, none of these congeners are true soil nematodes, that is, they normally do not reproduce and establish populations in soil. 1 In contrast to popular belief, C. elegans is not prevalent in the environment but has recently been recovered with regularity from terrestrial snails (Helix aspersa) in California, in what is likely a commensal relationship defined either by phoresy (e.g., transport to new habitat) or necromeny (the animal associate to be used as a food source upon death). 1,10
Attributes such as hermaphroditic reproductive capability and high reproductive rate also make C. elegans a good candidate for tissue colonization of a living animal. Some Caenorhabditis species, though not C. elegans, have been isolated from live vertebrates, but such reports have been singular events with the exception of C. bovis, which colonizes the ears of zebu cattle (Bos primigenius indicus) with otitis externa. In the case of C. bovis, colonization of the cattle ear seems to represent a case of exploitation of a bacteria-rich habitat rather than parasitism of the host tissues in the strict sense. 10 Other instances include C. clavopapillata, C. genitalis, and C. avicola, which were isolated from laboratory animals (mammals), an ill woman in Europe, and the intestine of a bird (Rhyacornis fuliginosus), respectively. Lastly, an unidentified Caenorhabditis sp. was reported in the liver of an African crocodile that died in captivity. 10
To date, there are no published reports of C. elegans associated with disease, either as a primary or opportunistic pathogen. Based on the histologic findings and progression of clinical signs in these frogs, the nematodes appear to have penetrated the cornea and subsequently migrated to the brain and incited an inflammatory response. The rare encapsulated and degenerate nematodes identified in the liver, kidney, and pancreas could not be compared with the nematodes in the eye and brain. Therefore, the nematodiasis in the frogs of this report was thought to be generally restricted to the eye and brain and unassociated with significant systemic migration. The foci of epidermal iridophore hyperplasia were interpreted as reactive, possibly to a chronic irritant, but whether this was related to invading nematodes is unknown. Discoloration or a change in epidermal pigmentation has been reported in African clawed frogs (Xenopus laevis) with cutaneous Pseudocapillaroides xenopi infection. 13,15 Dracunculus sp. have been associated with cutaneous disease in Indonesian White's tree frog (Pelodryas caerulea), and an unidentified nematode was found in skin scrapings from a captive Cayenne caecilian (Typhlonectes compressicauda). 15
There are 2 potential pathogeneses for the ocular and neural nematodiasis in these frogs. Either C. elegans has tissue-invasive capabilities that have not been recognized previously or the tissue invasion is opportunistic, secondary to damage caused by some other injurious agent or subsequent to general host immunosuppression. No other amphibians in any other exhibit were affected, suggesting that there was no common environmental factor (injurious agent) to predispose the frogs to cutaneous/ocular invasion by nematodes. General host immunosuppression is possible, but no other opportunistic infections were observed in any of the affected frogs. In addition, there was no histologic evidence of lymphoid depletion or lack of an immune response. In the surviving frog, it can only be concluded that the concurrent presence of nematodes with inflammation and resolution of disease after treatment support a causal relationship.
In summary, the present report identifies a novel association between the free-living nematode, C. elegans, and ophthalmic and neural disease in 4 captive Asian horned frogs. Nematodiasis should be considered on a differential list for corneal opacity, ulceration, and neurologic signs, particularly in captive amphibians. Surgical removal of affected tissue and anthelminthic therapy was successful in preventing progression of clinically identifiable disease in 1 frog. The current report demonstrates that Rhabdias spp. and Strongyloides spp. are not the only rhabditid nematodes that can colonize amphibians. Therefore, further identification of tissue nematodes should be pursued, particularly if the distribution of nematodes in tissue is not consistent with the recognized organs (lung and intestine) affected by the more common rhabditids. Freezing (−80°C) or preservation in 95% ethanol are preferred methods of sample storage for molecular diagnostics; however, neither is optimal for gross morphologic preservation of nematodes. Thus, a combination of preservation methods (e.g., formalin or 70% ethanol for gross morphology and the aforementioned methods for molecular diagnostics) is required for both morphologic and molecular characterization of nematodes. In addition, the Baermann technique can be used to extract living nematodes from unfixed infected tissue. Reduction of free-living nematode populations may be a key component in preventing infections in captive amphibians, and this can be accomplished by increasing the frequency of substrate changes and heat-sterilizing substrates and other surfaces in the environment.
Acknowledgements. The authors thank the clinical, pathology, and husbandry staff at the San Diego Zoo for technical assistance, in particular, John Kinkaid, Yvonne Cates, April Gorow, and Dr. Linda Lowenstine.