
Abstract
Brain tissue from 12 aborted bovine fetuses submitted to the Wisconsin Veterinary Diagnostic Laboratory revealed histologic lesions that consisted of glial nodules and variable degrees of mononuclear inflammation, microhemorrhage, neuronal necrosis, and cerebral cortical cavitation. A diagnosis of Bovine herpesvirus 1 (BHV-1) abortion had been made in all of these cases through multiple testing modalities. Brain tissue from 8 of the 12 fetuses was immunohistochemically stained with a monoclonal antibody specific to BHV-1, and, in 5 fetuses, there was positive intralesional staining of neurons, glial cells, and endothelial cells. This preliminary data suggested that herpesviral infection of brain tissue led to the described neurologic lesions. BHV-1 was then amplified from brain tissue in all 12 of the fetuses and was confirmed by partial sequencing of the thymidine kinase and glycoprotein C genes. To the authors' knowledge, neurologic lesions have not previously been described in BHV-1–infected fetuses, nor has BHV-1 previously been identified in bovine fetal brain tissue. The neurologic histopathology attributed to BHV-1 infection in these cases overlaps with the neurologic lesions produced by Neospora caninum, a common etiologic agent of bovine abortion. Therefore, when bovine fetal neurologic lesions are found, both etiologies should be considered and then distinguished by using additional diagnostic tools.
Introduction
Bovine herpesvirus 1 (BHV-1; family Herpesviridae, subfamily Alphaherpesvirinae, genus Varicellovirus), the etiologic agent of infectious bovine rhinotracheitis (IBR), establishes latency in sensory neurons, predominantly in the trigeminal ganglia, and in germinal centers in the pharyngeal tonsil. 19 Periodic recrudescence and shedding can occur in a manner similar to human herpes simplex and other mammalian herpes-viruses. Abortions because of BHV-1 occur 2 weeks to 2 months after the rhinotracheitis form of the disease or after vaccination of pregnant cows with modified live BHV-1 vaccines. 14 In natural outbreaks, up to 60% of a herd may abort, most commonly in the third trimester of gestation. Fetal expulsion typically occurs 24–36 hr after death, and there is often advanced postmortem autolysis. 11 Autolysis can limit identification of the classic histopathologic lesions, which include necrosis of the liver, lymph nodes, spleen, kidney, and placenta, and the presence of intranuclear inclusion bodies in these tissues.
Bovine herpesvirus 1 was initially subcategorized as BHV-1.1, BHV-1.2, and BHV-1.3, with BHV-1.3 referred to as the neurovirulent strain. 3 BHV-1.3 is now classified as BHV-5, bringing into question some previous reports of BHV-1 encephalitis. 2,10 Although BHV-5 is thought to be primarily responsible for fatal meningoencephalitis in calves, BHV-1 is occasionally neuroinvasive and can also cause encephalitis in calves and adult cattle. 12,16,17 Even though BHV-5 replication occurs both in neurons of the central nervous system (CNS) and in respiratory cells of the lung, trachea, and nasal mucosa, 12 the primary clinical manifestation is neurologic and the lesion produced is severe meningoencephalitis. Natural BHV-5 infections are reported from birth to 10 months of age. 12 Bovine herpesvirus 5 establishes latency in the trigeminal ganglia, as do both the vaccine and the natural strains of BHV-1. The CNS and the respiratory tract also appear to be sites for latency for BHV-5. 12
Histopathologic lesions in 8 Angus and 4 Holstein late-term BHV-aborted fetuses, submitted to the Wisconsin Veterinary Diagnostic Laboratory (University of Wisconsin, Madison, WI) over the course of 3 years, were reviewed. The diagnosis in all of these cases was based on classic gross and histopathologic lesions, including severe multifocal hepatocellular necrosis, and BHV-1–positive indirect fluorescent antibody staining of multiple fetal tissues. Encephalitis was observed in all cases. Because encephalitis has not been previously associated with BHV-1 abortion, brain tissue was then subjected to confirmatory diagnostics, including immunohistochemical (IHC) staining, virus isolation (VI), and polymerase chain reaction (PCR), followed by sequencing to identify the herpesvirus subtype. Finally, the possibility of infection with Neospora caninum, a common pathogen that causes similar fetal brain lesions (Fig. 1C, 1D) 15 and has been identified serologically in coinfections with BHV-1, 18 was investigated in all cases.
Summary of diagnostic tests performed to detect and/or confirm the presence of Bovine herpesvirus 1 (BHV-1).*

IHC = Immunohistochemistry performed on brain sections; FA = fluorescent antibody–positive tissue (L = lung; K = kidney; Li = liver; B = Brain; Pl = placenta); VI = virus isolation; H = Holstein breed; ND = specified test was not run; A = Angus breed.
All BHV-1 polymerase chain reaction (PCR) assays were positive on brain tissue; specific cycle threshold (Ct) level is given.
Materials and methods
Histopathologic and IHC specimen preparation
Tissues from necropsied fetuses were fixed in 10% buffered neutral formalin. Multiple tissues, including one or more sections of cerebral cortex and a section of brainstem, were paraffin embedded, sectioned at 5–8 μm, and stained with hematoxylin and eosin. Brain tissue blocks from 8 cases identified on the basis of lesions were then sectioned and mounted on charged glass slides. a The slides were incubated overnight with BHV-1 monoclonal antibody F-2 directed at BHV-1 glycoprotein C. b The slides were then stained with an enhanced alkaline phosphatase red detection kit c and hematoxylin counterstain. Positive and negative control tissues were tested concurrently.
Immunofluorescent antibody staining and Neospora serology
Fresh fetal tissues (brain, liver, lung, and kidney) were obtained at necropsy and were embedded in tissue-freezing media at −20°C and were sectioned. Cryosections were incubated at 37°C with a polyclonal antisera directed to BHV-1 and were conjugated to fluorescein isothiocyanate (FITC). b The slides were washed in phosphate buffered saline (PBS) and then mounted with buffered glycerol. The sections were examined with epifluorescence microscopy for apple-green fluorescence. For Neospora, a competitive enzyme-linked immunosorbent assay was performed on the first 9 samples (Table 1) by using fetal thoracic fluid, according to the manufacturer's protocol. b
Indirect fluorescent antibody
An indirect fluorescent antibody technique was performed on the last 3 samples (Table 1). Chamber slides with Neospora caninum tachyzoites b were incubated with serially diluted sera for 30 min at 37°C, followed by washes with PBS. Bovine antiglobulin conjugated to FITC was applied, and slides were incubated for 30 min at 37°C, followed by 3 washes with PBS. Slides were mounted with buffered glycerol and were observed for peripheral fluorescence.
Virus isolation
One-gram portions of tissues collected at necropsy (Table 1) were minced to approximately 3–5-mm pieces. Samples were diluted 1:10 in Hanks' Balanced Salt Solution d and were processed on a stomacher for 60 sec. Supernatant was transferred to primary bovine testicular cells and primary bovine turbinate cells. Cultures were incubated for 7 days and observed for cytopathic effect. Cultures were frozen, and supernatant was passed to bovine turbinate cells seeded in a chamber slide, which were observed for cytopathic effect for 2–3 additional days. Cells were stained by fluorescent antibody technique for the presence of BHV-1.
Nucleic acid extraction
Nucleic acid was extracted from 25 mg of fresh tissue samples by using the Qiagen DNeasy tissue kit, e by following the protocol recommended by the manufacturer. Nucleic acid was eluted in 50 μl of the supplied elution buffer. All paraffin-embedded tissues were pretreated by using a Chelex-100 f method, 9 followed by extraction with the Qiagen DNeasy tissue kit.
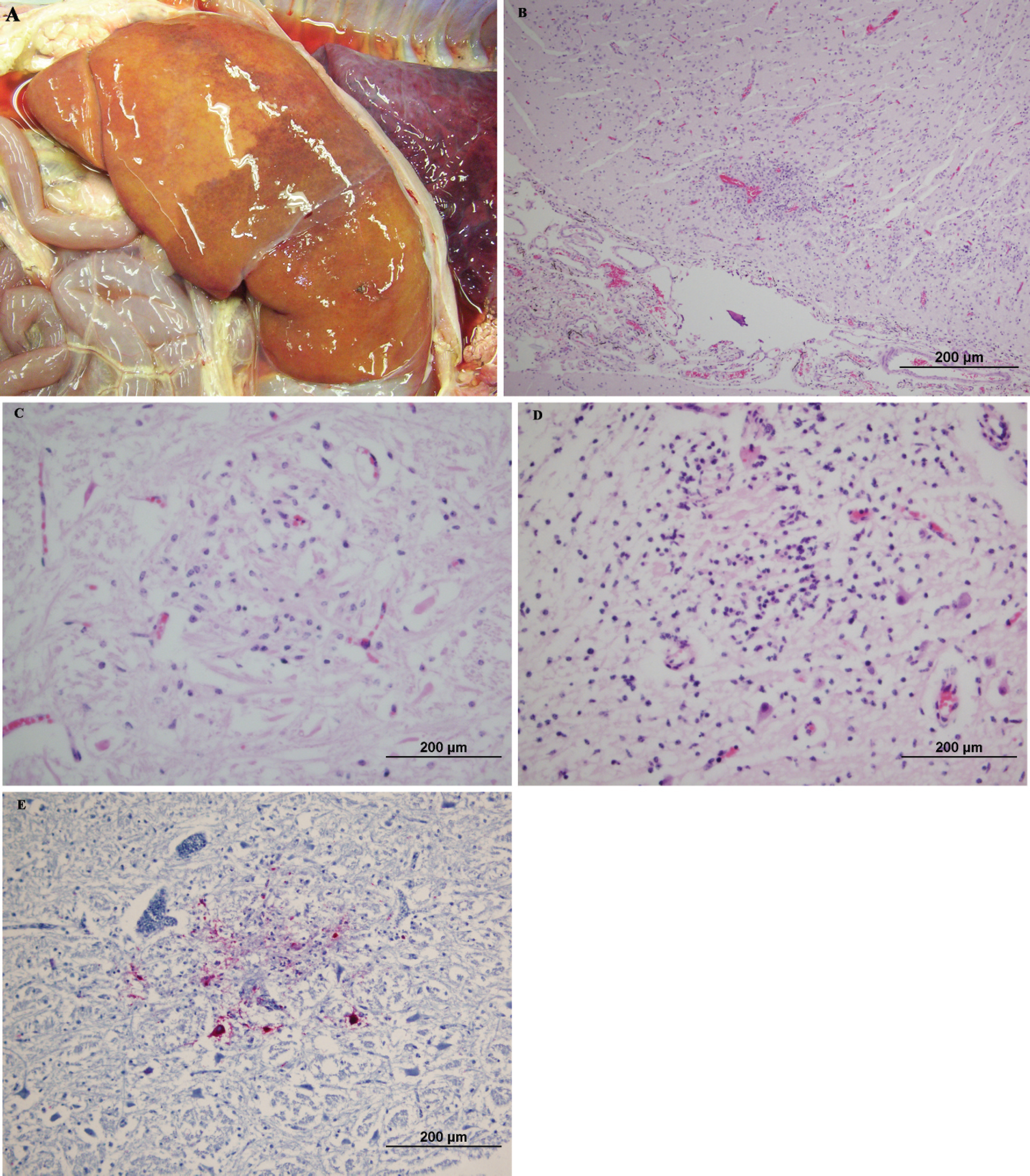
Aborted bovine fetal tissues.
Polymerase chain reaction
Polymerase chain reaction was performed on paraffin-embedded blocks for detection and confirmation of the presence of Neospora caninum and BHV-1. A validated protocol was used for brain tissue. Because of the problem of overfixing, unembedded fixed tissues were not used.
Bovine herpesvirus 1 real-time PCR was performed by using primers designed to amplify the thymidine kinase gene of BHV-1. 13 The forward and reverse primers and probe sequences (GenBank accession number D00438) are as follows: TK615F (5′-GGCACTGTGACCCTCGTGTT-3′), TK700R (5′-TTGATCTCGCGGAGGCAGTA-3′), and TK652 Probe (5′-FAM-CCGCGTGCCTCTGCTACCCCTTC-BHQ-1-3′). g The PCR included 400 nM of each primer, 200 nM of dual labeled fluorogenic probe, 12.50 μl of TaqMan Universal PCR Master Mix, h 6.00 μl of nuclease-free water, and 5 μl of template. The parameters for the real-time assay performed on either a 7000 or a 7500 ABI PCR instrument h are 1 cycle of 50°C for 2 min, 95°C for 15 min, and 40 cycles of 95°C for 15 sec, followed by 60°C for 1 min, 72°C for 1 min, and a final extension of 10 min at 72°C. Any results with cycle threshold values that exceeded 35 were retested to rule out contamination. A dilution of BHV-1 Colorado strain 036-BDV i was used as a positive control, and master mix with water instead of template was used as a negative control.
Neospora caninum real-time PCR was modified from the literature 8 to include a compatible dual fluorescent-labeled probe. The probe sequence was derived from GenBank accession number X00525. The published primer sequences and the newly designed probe are as follows: Nc5 Forward (5′-ACTGGAGGCACGCTGAACAC-3′); NC5 Reverse (5′-AACAATGCTTCGCAAGAGGAA-3′); Probe (5′-FAM-TGTCGTAAATCGGAGTTGCTTCTATGTGGC-Iowa Black FQ-3′). j The cycling parameters are identical to those for BHV-1.
Sequencing
To substantiate that the isolates were BHV-1, larger segments of the glycoprotein C and thymidine kinase genes were amplified and sequenced. Gene segments chosen were previously identified as having regions that differentiated BHV-1 from BHV-5. 4–6,13 Previously published primer sequences were chosen for the thymidine kinase gene: forward (TK1, 5′-TGGTACGGACGCCTTAAGTGG-3′) and reverse (TK2, 5′-GTTGATCTCGCGGAGGCAGTA-3′). 13 The following primer sequences were chosen for the glycoprotein C gene: BHV-1 gC 1F (5′-CATGGGCCCGCTGGGGCGA-3′) and reverse (BHV-1 gC 477R, 5′-CAGTGCACGTACAGCGGCT-3′). The master mix included 5.00 μl of Buffer A, 200 nM of each primer, 0.50 μl of AccuPrime GC-rich DNA polymerase, k 5.00 μl of template, and nuclease-free water up to a volume of 25 μl. Cycling parameters for the assay were as follows: a denaturation step of 95°C for 10 min, 40 cycles of 95°C for 1 min, 60°C for 1 min, 72°C for 1 min, and a final extension at 72°C for 10 min.
Polymerase chain reaction products were purified by using Qiagen QIAquick PCR purification kit, e by following the recommended protocol. The nucleic acid concentration was determined by spectrophotometry, and discrete bands were confirmed on agarose gels before submission for sequencing at the DNA Sequence Laboratory of the Genome Center of Wisconsin (University of Wisconsin, Madison, WI). All postsequence analysis of data was performed by using Vector NTI and AlignX software. k
Results
Pathology
Multifocal hepatocellular necrosis was the most severe gross and histopathologic lesion identified and was present in all of the 12 fetuses reviewed (Fig. 1A). In addition, present in all of the cases was mild-to-severe multifocal encephalitis (Fig. 1B). A postmortem change precluded a detailed description of the anatomic distribution of brain lesions, but lesions were identified in the cerebral cortex in 5 of the 12 cases and in the brainstem in 9 of the 12 cases. Glial nodules were a histologic feature present in all of the lesions. Additional findings included mononuclear inflammation with cavitation in the cerebral cortex, neuronal necrosis, and microhemorrhage. Intranuclear inclusion bodies were not found in any of the cases. All cases lacked the fetal cardiac lesions commonly associated with Neospora caninum infection. 15 There was positive IHC staining of intra-lesional perikaryon, neuronal processes, glia, and endothelial cells when a monoclonal BHV-1 antibody was used on brain sections from 5 of the 8 cases examined with this technique (Fig. 1E). Renal tissue was consistently fluorescent antibody–positive for BHV-1, but fluorescent antibody staining of brain cryosections was negative for BHV-1. Results are summarized in Table 1.
Serology, virology, and molecular diagnostics
Serologic testing for Neospora caninum was reviewed in all of the cases for which fetal thoracic fluid was available and was found to be negative. Bovine herpesvirus 1 was isolated from the kidneys of 2 cases and the lungs of one of the 5 cases that were cultured. Confirmation of BHV-1 infection of the brain tissue was established with PCR and sequence analysis. Detection of BHV-1 was consistently achieved by use of real-time PCR in brain tissue from all fetuses. Sequence analysis of all of the described isolates showed complete homology with BHV-1, approximately 70% homology with BHV-5 in the TK gene, and 37% homology in the glycoprotein C gene (Fig. 2). Neospora caninum PCR was performed on brain tissue from all of the fetuses and was found to be negative. Results are summarized in Table 1.

Thymidine kinase (
Discussion
It was initially unclear whether the described fetal brain lesions were an unrecognized or underappreciated manifestation of infection by BHV-1, an atypical manifestation of BHV-1, or the result of a different strain of BHV. Because of the known neurotropism and histologic description of encephalitis in BHV-5–infected calves, this strain was considered a possible etiologic agent. Fluorescent antibody and IHC results were inconsistent in the detection of BHV-1 in the brain, presumably because of the viral load, viral viability, or tissue integrity; nevertheless, in all cases, PCR with sequence analysis showed complete homology with BHV-1 and limited homology with BHV-5. The BHV-1 PCR was designed to rule out BHV-5 by selection of primer and probe sequences that define an amplicon more closely homologous to BHV-1. 13 Whereas BHV-1 and BHV-5 share 85% homology throughout most of the genome, there are regions where the 2 strains are more diverse. Among these are regions in the thymidine kinase, glycoprotein C, and glycoprotein E genes. 1,4–7,13,14 To distinguish BHV-1 from BHV-5, larger regions of the thymidine kinase gene and the amino terminus of glycoprotein C, which contain distinct species differences, were sequenced. Targeted primer regions were highly conserved between BHV-1 and BHV-5, and internal sequences allowed differentiation between the 2 species because of multiple mutations and deletions in BHV-5. The sequence results support an identification of BHV-1.
The encephalitis consistently present upon review of clinically typical BHV-1 abortions, and the described molecular investigations suggest that the observed neurologic infection and tissue damage that result in encephalitis is an underappreciated manifestation of BHV-1 fetal infection. The reason for the apparent inability of BHV-1 to cause significant neurologic lesions postpartum is unknown. Clearly, there are many host factors specific to the manifestation of intrauterine viral infections that do not exist postpartum, and, in some cases, viral factors, such as gene expression, vary with developmental stages of the fetus. 20 The severity of the lesions in other organ systems in the fetuses reviewed precluded the possibility of a live birth. It is unclear if mild in utero BHV-1 infections with live births and latent or active infections occur.
Because Neospora caninum causes fetal nonsuppurative encephalitis, the authors addressed the possibility of coinfections with BHV-1 by reviewing dam and fetal Neospora caninum serology, by looking for other histologic lesions associated with Neospora caninum infection, and by performing PCR for Neospora on extracted DNA from described lesions in paraffin-embedded brain tissue. All results indicated the absence of Neospora caninum infection in these cases. Other etiologic agents commonly associated with bovine abortion include Bovine viral diarrhea virus and Leptospira, which were also ruled out in all cases with negative fluorescent antibody, VI, and PCR results.
The brain has a limited range of morphologic responses to injury such that multiple infectious causes of abortion can result in similar lesions. Use of a multiple-agent real-time PCR assay for bovine abortion cases can rapidly resolve the ambiguity nonspecific lesions generate and will distinguish between BHV-1 and Neospora caninum fetal encephalitis.
Acknowledgements
The authors thank Beth Gray for her excellent histopathology technical support and Chris Schatz for editorial assistance.
Footnotes
a.
Fisher Scientific, Chicago, IL.
b.
VMRD, Pullman, WA.
c.
Ventana, Tucson, AZ.
d.
Sigma-Aldrich, St. Louis, MO.
e.
Qiagen Inc., Valencia, CA.
f.
Bio-Rad Laboratories, Hercules, CA.
g.
BioSearch, Novato, CA.
h.
Applied Biosystems, Foster City, CA.
i.
Center for Veterinary Biologics, Ames, IA.
j.
Integrated DNA Technologies (IDT), Coralville, IA.
k.
Invitrogen Corp., Carlsbad, CA.