
Abstract
Five diagnostic techniques performed on skin biopsies (shoulder region) and/or serum were compared for detection of bovine viral diarrhea virus infection in 224 calves 0–3 months of age, 23 calves older than 3 months but younger than 7 months, and 11 cattle older than 7 months. The diagnostic methods used were immunohistochemistry (IHC), 2 commercial antigen ELISAs, 1 commercial antibody ELISA, and realtime RT-PCR. Results of 249 out of 258 skin and serum samples were identical and correlated within the 3 antigen detection methods and the real-time RT-PCR used. Twenty-six of these 249 samples were BVDV-positive with all antigen detection methods and the real-time RT-PCR. Nine out of 258 samples yielding discordant results were additionally examined by RT-PCR, RT-PCR Reamplification (ReA), and antigen ELISA I on serum and by immunohistochemistry on formalin fixed and paraffin-embedded skin biopsies. Virus isolation and genotyping was performed as well on these discordant samples. In 3 cases, transiently infected animals were identified. Two samples positive by real-time RT-PCR were interpreted as false positive and were ascribed to cross-contamination. The antigen ELISA II failed to detect 2 BVDV-positive calves due to the presence of maternal antibodies; the cause of 2 false-positive cases in this ELISA remained undetermined. Only persistently infected animals were identified in skin samples by IHC or antigen ELISA I. The 3 antigen detection methods and the real-time RT-PCR used in parallel had a high correlation rate (96.5%) and similar sensitivity and specificity values.
Introduction
Bovine viral diarrhea virus (BVDV) is a pestivirus of worldwide occurrence that can cause severe economical losses due to decreased fertility, abortions, diarrhea, respiratory symptoms, and the dreaded state of persistent infection (PI) in calves. Persistently infected animals are virus shedders and therefore should be eliminated from a herd. Fetuses infected with the noncytopathic (ncp) BVDV between days 30 and 125 of gestation develop immune tolerance and will be born persistently infected (virus-positive and antibody-negative). Fetuses infected later in gestation mount an immune response and will be born antibody-positive and virus-negative. 9,12,23
A newborn calf becomes seropositive by suckling colostrum from an antibody-positive dam. Colostral antibodies can mask the presence of viral antigen in the blood or in the peripheral blood leukocytes (PBL) of young PI animals less than 3 months of age or even older. 4,12,15,16,23 Nevertheless, virtually all older PI animals are virus-positive and antibody-negative when they are infected only with 1 genotype and subtype of BVDV. 12,16 The main limitation of virus isolation from sera from PI animals below 3 months of age is that maternal antibodies interfere with growth of BVDV in cell cultures. 15,23 BVDV antibodies are indicators of an immune response rather than an indicator of a protective immunity. Following a homologous challenge neutralizing antibodies are protective and prevent disease. 5
Various methods are available for diagnosing acute or persistent BVDV infections. 6,15,16 These include: immunohistochemistry (IHC), several antigen- and antibody ELISAs, RT-PCR assays, and virus isolation. 4,6,7,8,14,15,16,20,21,23 The objective of this study was to compare the usefulness of 5 diagnostic methods for the identification of PI calves, namely IHC, 2 commercial antigen ELISAs, 1 commercial antibody ELISA, and real-time RT-PCR, with special attention to the possible interference of maternal antibodies with the diagnostic methods applied.
Materials and methods
Animals and sample collection
Snap-frozen (liquid nitrogen, −120°C) skin biopsies (6 mm in diameter) from the shoulder region and serum samples collected from 224 calves up to 3 months of age, 23 calves older than 3 months but younger than 7 months, and 11 cattle older than 7 months, sampled by veterinarians, were screened for BVDV by IHC, antigen ELISA I, a antigen ELISA II, b and antibody ELISA c as well as realtime RT-PCR. The most common breed was Brown Swiss.
Nine samples yielding discordant results were additionally examined with conventional RT-PCR, RT-PCR reamplification (ReA), antigen ELISA I on serum, and IHC on formalin-fixed paraffin-embedded skin biopsies; virus isolation and genotyping were also performed. The 9 animals from which these samples had been collected were not to be resampled. IHC staining was performed using half of each frozen skin biopsy that had been immersed in 5 ml 4% formalin for 18 hr. The serum samples were used for RT-PCR, RT-PCR-ReA, antigen ELISA I, and virus isolation.
Imiminohistoehemistry
Snap-frozen skin biopsies were treated following a previously described method. 21 Six-mm-thick sections of snap frozen skin biopsies were mounted on positive charged glass slides and fixed in cold acetone (- 20°C) for 10 min. The slides were dried at room temperature (room temperature (RT); 22°C). Treatment with 3% hydrogen peroxide (H2O2) containing 0.2% sodium azide (NaN3) was subsequently applied in order to quench the endogenous peroxidase (10 min at RT). Incubation with 1 of the following 2 antibodies was undertaken for 1 hr at 37°C: Ca3/34-C42 d (dilution 1:100; specificity: E2/gp55 and Erns/ gp48; specific for BVDV) and C16 e (dilution 1:100; specificity: NS2-3/nsp125/80; specific for pestiviruses). The secondary antibody, a peroxidase labeled anti-mouse antibody, f was applied for 30 min at RT, and the chromogen AEC g was used. The slides were thoroughly rinsed with phosphate buffer solution (PBS, pH 8.0) at the end of each step. Positive and negative controls were included.
Formalin-fixed, paraffin-embedded skin tissue sections were deparaffinized and rehydrated, rinsed in tap water for 5 min, and counterstained for 2 min in hemalum. Endogenous peroxidase was inactivated by treatment with 3% H2O2 containing 0.2% NaN3 in water for 10 min at RT. Afterwards the slides were digested with 0.01% pronase h for 10 min at RT. The slides were incubated for 1 hr at 37°C with the following 2 antibodies: C42 i (dilution 1:400; specificity: gp48) and 15c5 j (dilution 1:10,000; specificity: gp48). 10 Afterwards the ChemMate method k was applied according to the manufacturer's instructions. AEC was used as chromogen.
Antigen ELISAs
Antigen ELISA I is licensed for BVDV antigen detection in serum, plasma, whole blood, and skin biopsy samples. Microtiter plates are coated with specific monoclonal antibodies (Erns/gp44-48) to detect BVD-antigens. For testing of skin biopsy samples, a 2–3-mm piece was excised from the frozen shoulder skin biopsies and processed for antigen detection according to the manufacturer's instructions. 6 With these ELISA 258 skin biopsies and 255 serum samples were tested. Antigen ELISA II is an antigen capture ELISA that detects the Erns (E0, gp48) glycoprotein of the BVDV in serum and plasma. Microtiter plates are coated with antibodies specific for the Erns glycoprotein. Serum collected from the calves (255 out of 258 samples) was used in this ELISA as described by the manufacturer. 2
Antibody ELISA
A commercial antibody ELISA was used for detection of serum antibodies against BVDV following the manufacturer's protocol. 2 Microtiter plates used in this ELISA are coated with inactivated BVD-antigen, strain NADL.
Real-time RT-PCR
RNA isolation from the serum samples was performed using the QIAamp Viral RNA Mini Kit l according to the manufacturer's instruction (vacuum protocol), with the following modifications. Only 30 μl of the serum sample and 110 μl DEPC water instead of 140 μl sample volume was used. The RNA was eluted in 60 μl RNA storage solution. m The real-time RT-PCR was applied as previously described, using primers and probes from the 5′ UTR region. 8 Briefly, we used the TaqMan One-Step RT-PCR Master Mix Reagents Kit, n added the primes and probes as well as 5 μl of the isolated RNA in a final volume of 25 μl, and applied the following program in a ABI Prism 7700 Sequence Detection System o :1 × 30 min 48 °C (RT-step), 1 × 10 min 95°C (activating AmpliTaq Gold), and 45 × 15 sec. 95°C; 1 min 60°C (cycling).
RT-PCR
One-step RT-PCR was performed using a OneStep RT-PCR Kit p according to the manufacturer's instructions, using previously described primers. 8,19 Briefly, to the kit reagents the forward and reverse primers (end concentration 0.6 μM) and 5 μl of the isolated RNA to a final volume of 25 μl was added and the following program in a Biometra T3 Thermocycler q was applied: 1 × 30 min 50°C (RT-step), 1 × 15 min 95°C (activating HotStarTaq DNA polymerase), and 40 × 30 sec 95°C; 1 min 60°C; 1min 72°C (cycling). 5 ml of the PCR product was analyzed on a 1% agarose gel.
PCR Reamplification
0.5 μl of the PCR product was reused as template for a new PCR reaction using the same primers as in the RT-PCR. The HotSarTaq PCR Kit r was used in an end volume of 25 μl according to the manufacturer's instructions. The following program was applied in a Biometra T3 Thermocycler: 1 × 15 min 95°C (activating HotStarTaq DNA polymerase) and 40 × 30 sec. 95°C; 1 min 60°C; 1 min 72°C (cycling). 5 μl of the PCR product was analyzed on a 1% agarose gel.
Results from various tests performed on 249-calf samples with concordant skin biopsy results.

Virus isolation
Virus isolation was accomplished in cultured goat synovial membrane cells as previously described. 20 Serum was diluted 1/10 in medium, and cells were inoculated for 1hr at 37°C. Four parts of fresh medium were added, and the cells were incubated for 5 to 7 days. If the cells were positive in the immunoperoxidase staining, no further passage was accomplished; otherwise, a second passage was performed.
Genotyping
The RT-PCR products were electrophoresed on a 1% agarose gel using standard procedures. Appropriate bands were cut out and purified using a Gel Extraction Kit. s
Purified DNA was cloned using the TOPO TA Cloning Kit t according to the manufacturer's protocol. Minipreps were prepared with the Wizard Plus Minipreps DNA Purification System, u and DNA was sequenced v and analyzed w according to the manufacturer's instructions. Sequences were analyzed using the Wisconsin Package Version 10.1. x
Statistical analysis
Correlations were calculated using StatView 5.1. y Sensitivity, specificity with the IHC as relative gold standard, and unweighted kappa were calculated using Internet-based Clinical Research Calculators (http://faculty.vassar.edu/lowry/vsclin.html).
Results
Animals that were positive only in the real-time RT-PCR or in the RT-PCR ReA but negative in all other tests were regarded as transiently (acutely) infected. Animals that were positive in the IHC, the 2 antigen-ELISAs, and real-time RT-PCR were considered to be persistently infected. Based on these criteria, the results obtained with the 3 antigen detection methods and real-time RT-PCR were concordant in 249 of the 258 animals (Table 1). Twenty-six of these 249 animals were persistently infected, and 4 out of the 26 were BVDV antibody-positive. The 4 antibody-positive animals were 1 week, 3 weeks, approximately 2 months, and 3 months old, respectively. The remaining 223 cases were BVDV-negative with the 3 antigen detection methods and real-time RT-PCR; of these, 175 were antibody-positive and 48 were antibody-negative.
Nine skin biopsies out of 258 had discordant results with antigen-ELISA II or real-time RT-PCR on serum. They were additionally examined with conventional RT-PCR, RT-PCR ReA, and antigen ELISA I on serum and by IHC on formalin fixed paraffin-embedded skin biopsies and again with realtime RT-PCR. In addition, virus isolation (n = 7) and genotyping (n = 6) were performed. The IHC and real-time RT-PCR in these discordant samples were repeated twice and the serum samples in the antigen ELISA I 3 times (Table 2). Three of the 9 animals were classified as transiently infected because IHC on snap-frozen and formalin-fixed skin biopsies as well as the antigen ELISA I from skin biopsies failed to identify them. In 1 transiently infected animal the antigen ELISA I showed a borderline result (case number 4: 0.3 optical density units), which was interpreted as negative. Genotyping revealed subtype 1b in 2 cases, 1h and 1e in 1 case each.
In 2 further cases only the real-time RT-PCR was initially positive. The real-time RT-PCR was repeated twice and negative results were obtained. Furthermore, negative results were obtained with conventional RT-PCR, PCR ReA, and virus isolation as well as the antigen ELISA I from serum. The initial positive real-time RT-PCR result was therefore interpreted as a false positive, possibly resulting from cross-contamination.
Two persistently infected calves were not detected with antigen ELISA II, whereas all the other antigen detection tests and real-time RT-PCR were positive. These 2 calves also had antibodies against BVDV. Only low values for the antigen detection were obtained in the antigen ELISA I in case number 8 because of the small amount of serum available for testing. Case number 1 and 2 were classified as false-positive in the antigen ELISA II because all other antigen detecting tests and RT-PCR were negative (Table 2).
Using IHC as the relative gold standard, the antigen ELISA I had a sensitivity and specificity of 100%, the antigen ELISA II had a sensitivity of 93.5% and a specificity of 99%, and the real-time RT-PCR had a sensitivity of 100% and a specificity of 99%. The unweighted kappa values for IHC versus the antigen ELISA I was 1, for the IHC versus antigen ELISA II 0.92, and for the IHC versus realtime RT-PCR 0.96, respectively.
Results of various tests on 9 calves with discordant skin biopsy test results.
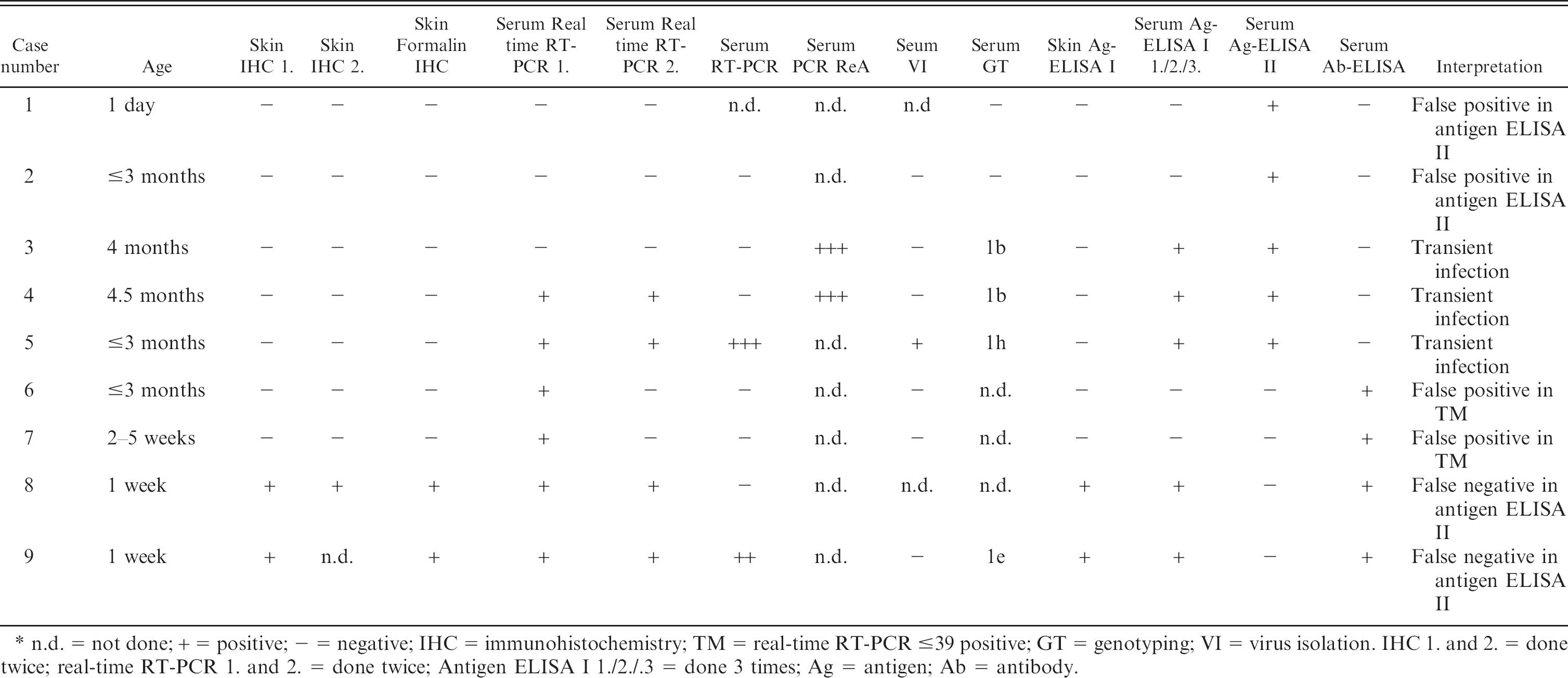
n.d. = not done; + = positive; — = negative; IHC = immunohistochemistry; TM = real-time RT-PCR s39 positive; GT = genotyping; VI = virus isolation. IHC 1. and 2. = done twice; real-time RT-PCR 1. and 2. = done twice; Antigen ELISA I 1./2./.3 = done 3 times; Ag = antigen; Ab = antibody.
Discussion
In the present study, results of 249 skin and 246 serum samples out of 258 animals examined were in agreement and correlated among the 3 antigen detection methods (IHC, Ag-ELISA I, and Ag-ELISA II) and real-time RT-PCR (correlation rate: 96.5%). The results were discordant in 9 animals (3.5%). Transiently infected animals were identified in 3 cases by Ag-ELISA I and II by real-time RT-PCR or the RT-PCR ReA in serum samples but not in skin biopsies. Persistently infected animals were identified in skin samples by IHC, antigen ELISA I and realtime RT-PCR. In a 1997 study, 20 BVDV was detected with equal sensitivity by IHC methods and cell culture virus isolation. On the other hand, a monoclonal antibody-based antigen detection ELISA was less sensitive than IHC. 20 Immunohistochemistry has been shown to be an accurate method for detection of neonatal PI calves and occasionally acute infections. 4,6,14 In a recent study, IHC and Ag-ELISA both showed high sensitivity for detecting BVDV-infected calves. 6 Immunohistochemistry can be applied on snap-frozen or formalin-fixed paraffin-embedded skin biopsies or a wide range of tissues, such as thyroid gland, bone, brain, and mucosal membranes. Virus can be detected in persistently infected animals, including those suffering from mucosal disease. 7,11,15,20,21
Antigen and antibody ELISAs are readily available and easy to use, and viral antigen can be detected in serum as well as whole blood and even skin biopsies. 2,6,12 Reverse-transcription PCR (RT-PCR) has become a routine diagnostic method and is highly sensitive as well as specific, and has the advantage that it allows pooling of samples, 16 and viral RNA can be detected independently of specific antibodies. 16
Virus isolation is the classical method and is still the gold standard, but the technique is laborious and sometimes requires more than 1 passage. The growth of BVDV in cell culture may or may not cause any visible cytopathic effects, leading to the classification of the BVDV into 2 biotypes: cytopathic (cp) and noncytopathic (ncp). In the laboratory, most (70%-90%) BVDV isolates are of the ncp biotype. 15 Saliki and Dubovi 15 suggest that acutely infected animals will not have a significant amount of antigen in the skin tissue in the vast majority of cases and that skin samples should be obtained for PI status determination; they further suggest that antigen-capture ELISAs are primarily useful in screening for PI cattle. Brodersen et al. 4 also state that skin samples from transiently infected cattle are rarely positive by IHC.
Two studies have claimed that in some cases acutely infected animals can be detected by IHC in skin. In 1 study, viral antigen was detected between days 10 and 14 postinfection in 4 out of 10 calves by IHC on paraffin-embedded skin samples. The calves had been infected intranasally with 108 TCID50 of ncp type 2 BVDV. The viral antigen distribution was patchy and restricted to superficial epithelial structures. 14 This distribution, however, was not typical for persistently infected animals, where, according to the present authors' experience, in addition to epithelial structures smooth muscle cells of blood vessels regularly show positive immunostaining. 14,21 Another recent study detected at initial screening 8 acutely infected calves out of 559 calves tested for BVDV by different methods. Six out of these 8 cases were positive by ear notch IHC with an intensity and distribution of BVDV antigen similar to PI calves, and all 8 were positive by antigen-ELISA. Virus could not be isolated from these calves. 6 These 2 studies 6,14 contrast with the results of the current study, in which only persistently infected calves could be identified by IHC and antigen ELISA I on skin biopsies. There are 2 possible explanations for the apparent disagreement between the results of the present study and the 2 previous studies with regard to detectability of BVDV antigen in skin biopsies from PI cattle: in the study by Njaa et al., 14 calves were infected with a high dose of virulent type 2 BVDV; both the high dose and the genotype could be explanations for the virus detection in the skin in these acutely infected calves. In the other report by Cornish et al., 6 most cases belonged to genotype 1 and subtype a. Here again, genotypic differences among the viruses present in various geographic locations may be a possible explanation for the discrepancies. Indeed, a recent survey of virus isolates in Austria described 70 BVDV-1 isolates and only 1 BVDV-2 isolate. The BVDV-1 isolates were mostly of the 1f genetic group, followed in descending order by 1h, 1b, 1d, and 1g; no subtype 1a (which is highly prevalent in the UK and the USA) 22 was found. In the present study, the 3 BVDV isolates were subtyped as 1b, 1h, and 1e, which is in agreement with previous findings in Switzerland 18 and different from USA and UK isolates. These differences in BVDV subtypes circulating in various geographic locations might explain the differences in the distribution of the viral antigen.
Two samples scored as positive by real-time RT-PCR were interpreted as false positives and were ascribed to cross-contamination. The antigen ELISA II failed to detect 2 out of 6 calves below 3 months of age, whereas the 2 other antigen detection methods and real-time RT-PCR were able to detect these 2 animals. The failure to detect the 2 calves by antigen ELISA II was most likely due to the presence of maternal antibodies. This phenomenon is also well known in lambs with pestiviral infections. High levels of colostral antibodies in viremic lambs younger than 2 months of age can mask persistent viremia, resulting in false-negative virus isolation and ELISA results. 3,13 The use of diagnostic methods to detect virus or viral antigen in serum or PBL is not recommended in animals less than 3 months of age, due to presence of colostral antibodies. 4,15 It can be concluded from the present and other studies that IHC from skin biopsies, as well as RT-PCR methods, are able to detect BVDV even in the presence of colostral antibodies. This also seems to be true for the antigen ELISA I from skin biopsies or serum. 3,4,15,16,23 In a previous study, the authors investigated 117 calves from dams that had been kept on Swiss communal alpine pastures and identified 9 calves as persistently BVDV-infected using IHC on skin biopsies. 17 The same animals were tested in parallel using antigen ELISA II and antibody ELISA, but only 6 out of that 9 were antigen-positive. The 3 negative animals were less than 11 days old and antibody-positive, while the other 6 animals were antibody-negative. 17 Skin and serum samples from 8 of these 9 BVDV-positive calves were all positive when retested by antigen ELISA I (data not shown). This is in agreement with the present results, showing that maternal antibodies usually do not interfere with IHC or with the antigen ELISA I.
Another recent study 23 also investigated the effect of maternal antibodies on the detection of BVDV in peripheral blood samples after intrauterine experimental infection of calves by means of RT-PCR, virus isolation, and antigen ELISA z . The RT-PCR was not affected by maternal antibodies, but virus isolation and the antigen ELISA were often false-negative because of interference by maternal antibodies. The higher the maternal antibody titer, the lower the frequency of positive virus isolation. 23
For unknown reasons, 2 animals tested positive for antigen by antigen ELISA II but were negative by the other 2 antigen detection methods and RT-PCR. They were interpreted as false positives.
In conclusion, the 3 antigen detection methods and real-time RT-PCR used in parallel in this study had a high correlation rate of 96.5% in recognizing PI BVDV-infected animals; 10.8% of 258 animals were PI, and 1% were recognized as transiently infected. A proportion of PI calves (7.1%) were falsely negative by the antigen ELISA II due to the interference of maternal antibodies. Another 0.77% were falsely positive in the real-time RT-PCR, possibly due to cross-contamination; similarly, 0.77% were falsely positive in the BVD-Virus-II ELISA for undetermined reasons. The risk of failing to detect PI calves is of high significance, since these animals shed large amounts of virus and should be identified with absolute reliability. IHC on skin biopsies, antigen ELISA I, and real-time RT-PCR appear to be reliable tools for this task. Each method had a error rate of 0.77 to 1%, which resulted from failure to detect transiently infected animals (1%). However, this is a negligible rate, because transiently infected animals do not represent a major risk for herdmates and are not of epidemiologic relevance. The 3 antigen detection methods and real-time RT-PCR used in this study had a high sensitivity and specificity. The unweighted kappa value revealed very good agreement among the tests. Therefore, the choice of which test a particular laboratory should use will depend on other criteria such as cost, ease or convenience of handling samples, and simplicity of the test method.
Footnotes
a.
HerdChek*BVDV Ag/Serum Plus: HerdChek Bovine Diarrhea Virus Test Kit, Idexx Laboratories, Westbrook, ME.
b.
CHEKIT-BVD-Virus-III. Dr. Bommeli AG, Bern-Liebefeld, Switzerland.
c.
CHEKIT-BVD-Sero-II. Dr. Bommeli AG, Bern-Liebefeld, Switzerland.
d.
Ca3/34-C42, dilution 1:100, specificity: E2/gp55 and Erns/gp48. Dr. Bommeli AG, Bern-Liebefeld, Switzerland.
e.
C16, dilution 1:100, specificity: NS2-3/nsp125/80. Dr. Bommeli AG, Bern-Liebefeld, Switzerland.
f.
DAKO Cytomation, EnVision+, peroxidase, mouse, Zug, Switzerland.
g.
AEC (Amino-Ethyl-Carbazole), 00-2007 Substrate Kit, Zymed Laboratories, San Francisco, CA.
h.
DAKO Cytomation, Pronase, Zug, Switzerland.
i.
C42, dilution 1:400, specificity: gp48. Prof. Moennig, Institute of Virology, Hannover, Germany.
j.
15C5, dilution 1:10,000, specificity: gp48. E. Dubovi, New York State College of Veterinary Medicine, Cornell University, Ithaca, NY.
k.
DAKO Cytomation, ChemMate, peroxidase, mouse, Zug, Switzerland.
l.
QIAmp Viral RNA Mini Kit, Qiagen, Hombrechtikon, Switzerland.
m.
RNA storage solution, Ambion (Europe), Cambridgeshire, UK.
n.
TaqMan One-Step RT-PCR Master Mix Reagents Kit, Applied Biosystems, Rotkreuz, Switzerland.
o.
ABI Prism 7700 Sequence Detection System, Applied Biosystems, Rotkreuz, Switzerland.
p.
QIAGEN OneStep RT-PCR Kit, Qiagen, Hombrechtikon, Switzerland.
q.
Biometra T3 thermocycler, BIOLABO, Chatel-St-Denis, Switzerland.
r.
HotSarTaq PCR Kit, Qiagen, Hombrechtikon, Switzerland.
s.
QIAEX II Gel Extraction Kit, Qiagen, Hombrechtikon, Switzerland.
t.
TOPO TA Cloning Kit, Invitrogen, Basel, Switzerland.
u.
Wizard Plus Minipreps Dann Purification System, Promega, Wallisellen, Switzerland.
v.
Perkin Elmer Dye Terminator DNA Sequencing methods, Perkin Elmer, Foster City, CA.
w.
ABIprism 310 or Applied Biosystem Model 373A, Perkin Elmer, Foster City, CA.
x.
Genetics Computer Group (GCG), Madison, WI.
y.
StatView 5.1, SAS Inc., Cary, NC.
z.
Serelisa BVD/MD Ag Mono Indirect, Synbiotics Europe, Lyon, France.