
Abstract
Objectives:
Adiponectin is a cytokine secreted from adipose tissue that regulates energy homeostasis, inflammation, and cell proliferation. Obesity is associated with increased risk of various cancers, including ovarian cancer. Adipokines, including adiponectin, have been implicated as a factor linking obesity and carcinogenesis. The oncogenic role of adiponectin is not known with regard to various cancer types. We sought to determine the role of adiponectin in angiogenesis in ovarian cancer in vitro.
Methods:
We transfected SKOV3 cells with vascular endothelial growth factor small interfering RNA in order to identify the independent angiogenic role of adiponectin in ovarian cancer. The vascular endothelial growth factor knockdown SKOV3 cell lines were treated with adiponectin for 48 h. The cytokines involved in adiponectin-mediated angiogenesis were explored using the human angiogenesis cytokine array and were verified with the enzyme-linked immunosorbent assay. The angiogenic effect of adiponectin was evaluated using the human umbilical vein endothelial cell tube formation assay. We also investigated the effects of adiponectin treatment on the migration and invasion of SKOV3 cells.
Results:
The number of tubes formed by human umbilical vein endothelial cell decreased significantly after knockdown of vascular endothelial growth factor (via transfection of vascular endothelial growth factor small interfering RNA into SKOV3 cells). When these vascular endothelial growth factor knockdown SKOV3 cells were treated with adiponectin, there was an increase in the number of tubes in a tube formation assay. Following adiponectin treatment, the CXC chemokine ligand 1 secretion increased in a cytokine array. This was confirmed by both enzyme-linked immunosorbent assay and Western blot. The increased secretion of CXC chemokine ligand 1 by adiponectin occurred regardless of vascular endothelial growth factor knockdown. In addition, the induction of migration and invasion of SKOV3 cells were significantly stronger with adiponectin treatment than they were without.
Conclusion:
Adiponectin treatment of ovarian cancer cells induces angiogenesis via CXC chemokine ligand 1 independently of vascular endothelial growth factor. These findings suggest that adiponectin may serve as a novel therapeutic target for ovarian cancer.
Introduction
Obesity is a serious health problem that contributes to metabolic disorders and all-cause morality. 1 The excess fat accumulation in the adipose tissue leads to an imbalance of cytokines and growth factors. 2 Increasing body mass index is also associated with increased risk of various types of cancer, including ovarian cancer. 3 Obese patients with cancer are also at higher risk of death. Up to 14% of cancer-related deaths can be attributed to obesity. 4 Prior studies have investigated which components of obesity are related to carcinogenesis. It is suspected that adipokines secreted in the adipose tissue are associated with the link between obesity and carcinogenesis. 5
Adiponectin (APN) is a cytokine that is secreted from adipose tissue. It plays a role in preventing metabolic and vascular disease through anti-inflammatory and anti-atherogenic effects. 6 Given its important roles in energy homeostasis, inflammation, tissue remodeling, and cell proliferation, APN has been studied as a therapeutic target in the management of several diseases, including metabolic syndrome.7,8 APN is generally found at higher levels in women than it is in men. Its average circulating concentration in women is 5–30 ng/mL. 9 APN levels are generally higher in non-obese women and postmenopausal women than they are in others. 10
Prior studies demonstrate both positive and negative correlations between the APN level and the risk of various cancers.11–13 Several malignancies, including breast and colorectal cancer, are associated with decreased APN levels. 14 Lower levels of serum APN suggest a higher risk of developing endometrial cancer, although the difference was not significant in ovarian cancer. 15 One study found that there was no difference in the serum levels of APN in women with ovarian carcinoma and those with benign tumors. 16
Several reports have suggested that APN has antitumor activity against ovarian cancer. 15 In contrast, others have found that APN is not related to the risk of ovarian cancer. 16 Furthermore, there is controversy regarding whether APN induces or inhibits angiogenesis by modulating various signaling pathways.17–19 Therefore, the precise role of APN in angiogenesis and tumor development remains unclear. The role of APN in angiogenesis in ovarian cancer is also largely unknown. In this study, we investigated the role of APN in ovarian cancer in vitro, with a particular focus on angiogenesis. Understanding the pathways and the effect of APN on angiogenesis in ovarian cancer may provide meaningful treatment options for this lethal disease. We also investigated APN-induced migration and invasion of ovarian cancer cells.
Materials and methods
Cell lines and cell culture
Four human epithelial ovarian cancer (EOC) cell lines (SKOV3, CAOV3, A2780, and ES-2) were used in this study. The CAOV3 was obtained from the Korean Cell Line Bank (Seoul, Korea). The SKOV3, A2780, and ES-2 cells were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). The SKOV3 and ES-2 cells were grown with McCoy’s 5A Medium (ATCC) containing 10% fetal bovine serum (FBS; Life Technologies, Grand Island, NY, USA), 1.5 mM L-glutamine, 100 U/mL penicillin, and 100 mg/mL streptomycin. CAOV3 was grown with Dulbecco’s modified Eagle’s medium (DMEM, Life Technologies) containing 10% FBS (Life Technologies), 100 U/mL penicillin and 100 mg/mL streptomycin. A2780 was grown with Roswell Park Memorial Institute medium 1640 (RPMI1640, Life Technologies) containing 10% FBS (Life Technologies), 25 mM HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), 100 U/mL penicillin, and 100 mg/mL streptomycin. Cells were grown in a 100 mm dish at 37°C and 5% CO2. The cell growth and morphology were routinely monitored.
Cell transfection, siRNA gene knockdown, and APN treatment
Adiponectin receptor 1 (ADIPOR1) small interfering RNA (siRNA; Santa Cruz) is a pool of three target-specific 19-25 nt siRNAs designed to knockdown gene expression. The siRNA that targets human vascular endothelial growth factor (VEGF) was purchased from Bioneer Inc (Daejeon, South Korea). The double-stranded RNA oligonucleotides used specific for the VEGF coding region are as follows: siRNA sense: 5′-CAGAACGAUCGAUACAGAA-3′ and siRNA anti-sense: 5′-UUCUGUAUCGAUCGUUCUG-3′. Transient transfections were performed according to the manufacturer’s instructions. The SKOV3 cells were plated (2 × 105cells per well) to obtain 70%–80% confluency in a six-well plate. It was transfected with siRNA using Lipofectamine RNAiMax Reagent (Invitrogen, Carlsbad, CA, USA). Eight hours after transfection, the fresh media were added to the plates. Transfection efficiency was confirmed by real-time polymerase chain reaction (PCR) analysis (performed in triplicate). The stably transfected SKOV3 cells were called SKOV3/control-siRNA (transfected with control-siRNA) cells and SKOV3/VEGF-siRNA (transfected with VEGF-siRNA) cells. The transfected cells were then treated with 80 ng/mL of human recombinant APN (R&D Systems) for 48 h.
Real-time PCR
Real-time PCR was performed on the Applied Biosystems 7300 PCR system (Applied Biosystems, Foster City, USA) using the SYBR-Green Master PCR mix (Applied Biosystems) following the manufacturer’s instructions (performed in triplicate). The total RNA was extracted from SKOV3 cells treated with or without APN using the RNA Mini Kit (Invitrogen). cDNA synthesis was performed using the Reverse Transcription Kits (Applied Biosystems). The threshold cycle (Ct) value and efficiency of PCR amplification for each set of primers were determined using SDS 2.2 software (Applied Biosystems). The gene expression levels of each transcript were determined using the comparative Ct method using the human glyceraldehyde-3′-phosphate dehydrogenase (GAPDH) mRNA as a reference. The following oligonucleotides were used as VEGF PCR primers (Cosmo Genetech): sense: 5′-CTGTCTTGGGTGCATTGGAG-3′ and anti-sense: 5′-TGATGATTCTGCCCTCCTCC-3′.
Western blot analysis
Cell lysates were prepared in lysis buffer (protein extraction solution, iNtRon biotechnology). The lysates were centrifuged at 13,000 r/min for 10 min at 4°C. The protein concentration in the supernatants was then determined using the BCA protein assay kit (Thermo Fisher Scientific, Seoul, Korea). A 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was performed, and 30 μg of protein of each sample was analyzed. The proteins in the SDS gels were transferred to a nitrocellulose membrane (Amersham hybond ECL) using an electroblot apparatus. The membranes were incubated overnight at 4°C with primary antibodies against the ADIPOR protein (Abcam, Cambridge, UK) and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA). After extensive washing, bands were detected using horseradish peroxidase–conjugated secondary goat anti-mouse IgG antibodies (BIO-RAD Laboratories, Hercules, CA, USA). Visualization of the secondary antibody was performed using a chemiluminescence detection procedure according to the manufacturer’s protocol (Amersham Biosciences, Japan). Western blot analyses were performed in triplicate.
Cytokine array and enzyme-linked immunosorbent assay
The production of angiogenesis-related factors were determined using a human angiogenesis cytokine array (C-Series 1000, RayBiotech Inc, Norcross, GA, USA), which detects the expression of 43 angiogenic factors in two membranes. The array was conducted according to the manufacturers’ protocols using medium from SKOV3 cells in triplicate, which were transfected with siRNA followed by APN treatment (for 48 h). The membranes from the cytokine array kit were incubated with 1 mL of concentrated medium at room temperature for 2 h. The membranes were then washed three times with wash buffer I and twice with wash buffer II at room temperature. Each wash lasted 5 min. The membranes were then assayed using chemiluminescence technique. In order to verify the results, the same samples and concentrated cultured medium were assayed using enzyme-linked immunosorbent assay (ELISA). The commercially available sandwich ELISA kits (RayBiotech Inc) were used to evaluate the cytokine levels of VEGF and CXC chemokine ligand 1 (CXCL1) in the cultured medium derived from SKOV3 cells.
In vitro tube formation assay
Cells were cultured in an Endothelial cell Basal Medium-2 bullet kit (Lonza, Seoul, Korea) according to the manufacturer’s instructions. The human umbilical vein endothelial cells (HUVECs) were used for these studies after three to four passages. The cultures were negative for mycoplasma contamination. The HUVECs were characterized by their homogeneous and typical cobblestone morphology on electron microscopy. All cells were maintained in a 5% CO2 atmosphere at 37°C. For all of the functional and biological assays, cells with >95% viability were used at 80%–90% confluence.
The SKOV3 cells were cultured in six-well plates (performed in triplicate) with fresh complete medium for 24 h, and 1 mL cultured medium was collected. For the tube formation assay, the 96-well plates were coated with Matrigel (20 μL per well; Corning, Steuben County, NY, USA) and kept in 5% CO2 atmosphere at 37°C for 30 min. The 1 × 104 HUVECs were then suspended in 50 µL cultured medium and applied to the pre-coated 96-well plates. After incubating at 37°C for another 6 h, the images were captured under a microscope, and the tubular structures formed in the Matrigel were counted at ×100 magnification in three randomized fields.
Cell migration and invasion
The SKOV3 cell lines were seeded (1 × 105 cells/mL) into the upper chamber of a BD-Falcon Transwell system (12-well format; 8 µM/L pore size, Corning Costar, Tewksbury, MA, USA) in serum-free McCoy’s medium. Cell migration was induced by adding APN 1 µg to the lower chamber, which was filled with the same medium containing 10%–20% FBS. After 48 h, the migrated cells on the bottom surface of the membrane were fixed with 4% paraformaldehyde for 30 min. The cells were then stained with 0.1% crystal violet for 30 min. The migrated cells were counted in three randomly selected fields (×100), and the average number was determined. In order to detect invasion, we used the same protocol that was used for migration, except for the use of Matrigel in the invasion assay. Matrigel (1.2 mg/mL; Becton, Dickinson and Company, NJ, USA) was spread in the upper chamber of a transwell. They were performed in triplicate.
Statistical analysis
All of the experiments were performed at least in triplicate. The data were compiled from three separate experiments. The results of multiple observations were calculated as means ± standard deviations (SDs). Student’s t-test was used to analyze the differences between the two groups using SPSS for windows Version 20.0 (SPSS Inc, IL, USA). P-values <0.05 were considered statistically significant.
Results
Basal expression of ADIPOR1 and ADIPOR2
In order to select suitable cell lines to study the role of APN, we first analyzed the basal expression of ADIPOR1 and ADIPOR2 in four ovarian cancer cell lines (SKOV3, CAOV3, ES-2, and A2780; Figure 1). The ADIPOR1 expression was relatively overexpressed in SKOV3 cells. In contrast, there were comparable expression levels of ADIPOR1 across all EOC cell lines. The expression of ADIPOR2 was lower than that of ADIPOR1 in all of the cell lines.
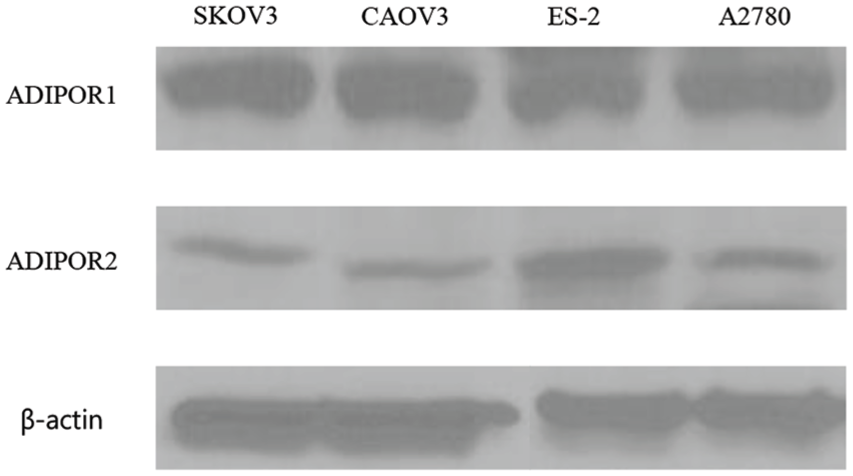
Basal expression of ADIPOR 1/2.
The effect of APN treatment on VEGF expression
The expression of VEGF in ovarian cancer cell lines grown with APN is shown in Supplementary Figure S1. In the SKOV3 cells (Supplementary Figure S1A), APN did not produce an effect on the VEGF levels in either a time-dependent or dose-dependent manner. APN also did not have a significant effect on VEGF in A2780, CAOV3, and OVCAR3 cells (Supplementary Figure S1B–1D).
When ovarian cancer cell lines were treated with APN, there was no significant change in tube formation compared to that of the control group (data not shown). Therefore, we failed to identify an effect of APN on angiogenesis. However, it remains possible that APN has an effect on angiogenesis in the absence of the effects of VEGF, which is a major pro-angiogenic factor in ovarian cancer cell lines.
Downregulation of VEGF by VEGF-siRNA in SKOV3 cells
The expression of ADIPOR1 was relatively increased in SKOV3 cell lines than that of other ovarian cancer cell lines. Therefore, we investigated the SKOV3 cell lines further. We established a VEGF-downregulated EOC cell line that could evade the angiogenic effect of VEGF in order to evaluate APN’s role in angiogenesis in ovarian cancer in the absence of VEGF. Either VEGF-siRNA or non-specific control-siRNA was transfected into the SKOV3 cells. Eight hours later, we measured the expression levels of VEGF mRNA using real-time PCR. The VEGF mRNA level declined significantly in the SKOV3/VEGF-siRNA than that of the SKOV3/control-siRNA (P < 0.01; Supplementary Figure S2A). The VEGF protein expression was also decreased in the SKOV3/VEGF-siRNA, which was confirmed by ELISA (Supplementary Figure S2B). These results suggest that VEGF-siRNA could effectively suppress VEGF expression at both the mRNA and protein levels in SKOV3 cells.
APN induces angiogenesis in SKOV3/VEGF-siRNA
In order to determine the actual role of APN on ovarian cancer angiogenesis, we used the tube formation assay with medium of SKOV3/VEGF-siRNA. As shown in Figure 2(a), HUVEC had low ability to form tubes in the medium from the SKOV3/VEGF-siRNA. The mean number of complete tubular structures was significantly lower in the medium from the SKOV3/VEGF-siRNA cells (5.5 ± 2.1 in SKOV3/VEGF-siRNA) than it was in the corresponding control (38 ± 2.8 in SKOV3 and 28.5 ± 3.5 in SKOV3/control-siRNA, P < 0.05 for all; Figure 2(b)). However, there was a significant increase in the mean number of complete tubular structures in the cultured medium from SKOV3/VEGF-siRNA with APN treatment (14.5 ± 0.71, P < 0.05) than in those without APN treatment. These data suggest that APN treatment significantly enhanced the angiogenic capacity of ovarian cancer cells in the absence of VEGF.
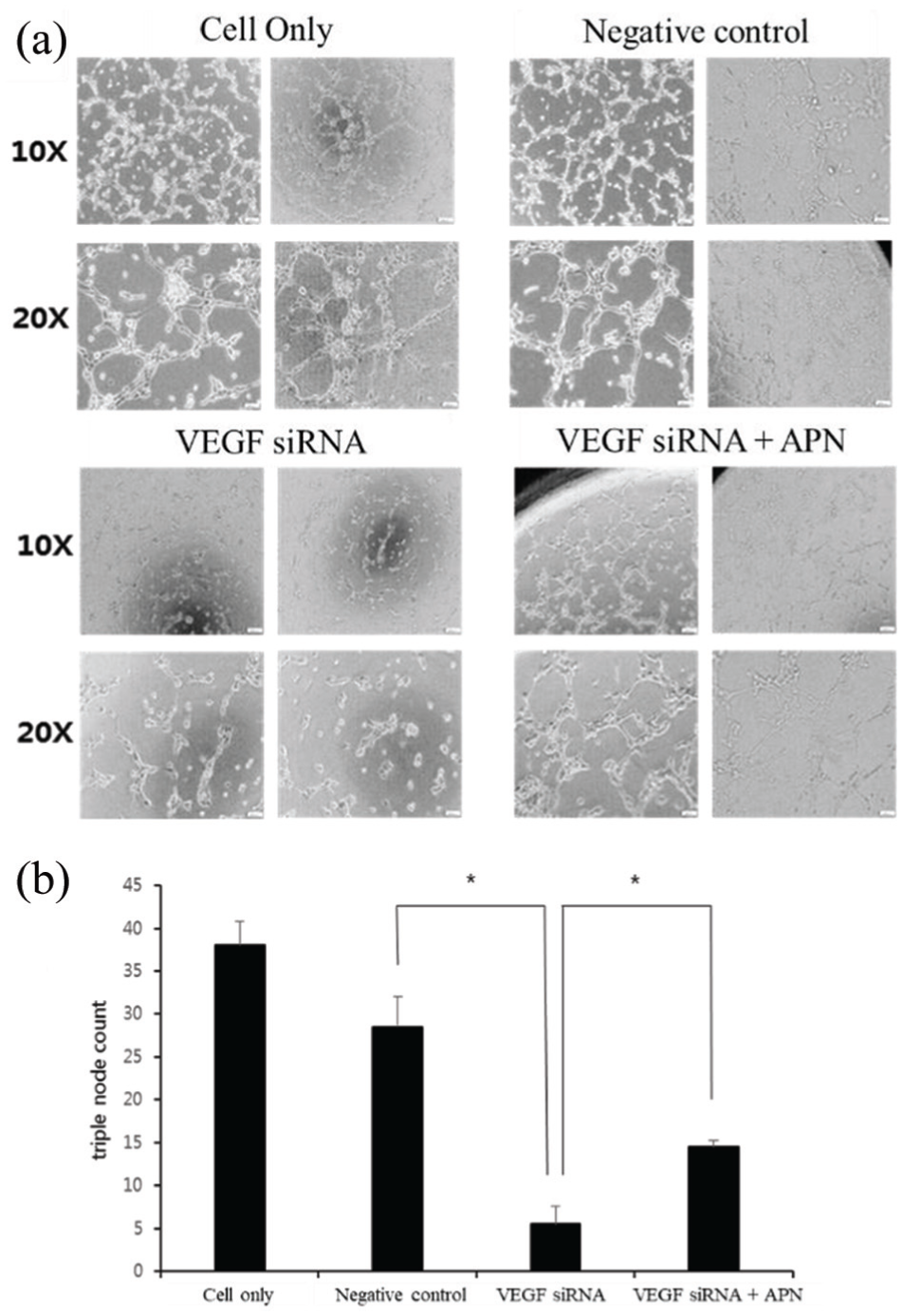
Tube formation in HUVECs following adiponectin treatment in SKOV3 cells: (a) SKOV3 cells were transfected with either negative control-siRNA or VEGF-siRNA. VEGF knockdown in SKOV3 inhibited HUVECs tube formation. APN-treated cultured medium in VEGF-knockdown SKOV3 cell lines induced tube formation of HUVECs. (b) The number of triple node was counted in SKOV3, SKOV3/control-siRNA, SKOV3/VEGF-siRNA, and APN-treated SKOV3/VEGF-siRNA.
APN induces angiogenesis via CXCL1 overexpression
We studied the expression of various cytokines following APN treatment in SKOV3 cells in order to determine which molecule was involved in APN-mediated angiogenesis in ovarian cancer. We used a cytokine array that could detect the expression of 43 angiogenic factors in two membranes. The secreted levels of VEGF were not significantly altered by APN treatment (not shown in figure). However, the CXCL1 level was higher in the cultured medium from APN-treated SKOV3 cells than it was in those that were not treated with APN. This CXCL1 overexpression after APN treatment was also shown in the cultured medium from both SKOV3/control-siRNA and SKOV3/VEGF-siRNA cells (Figure 3(a)). Furthermore, decreased tube formation in the cultured medium from SKOV3/VEGF-siRNA cells was reversely increased after adding recombinant CXCL1 (Supplementary Figure S3).
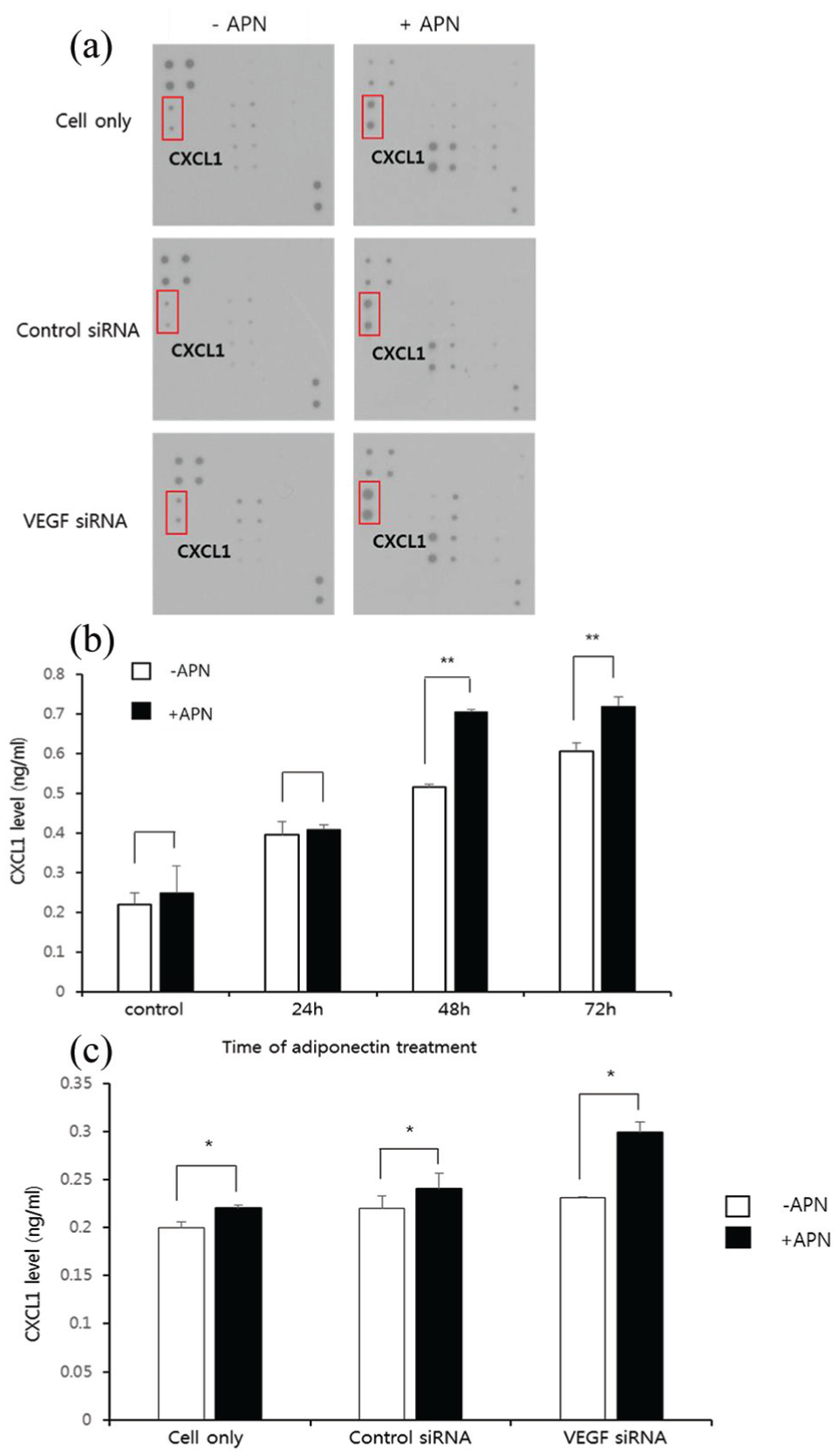
Cytokine array and APN-induced CXCL1: (a) cytokine expression array shows 20 cytokines on membrane secreted by SKOV3/control-siRNA, SKOV3/VEGF-siRNA. VEGF knockdown did not disturb CXCL1 secretion induced by APN. (b) The effect of APN on expression of CXCL1 was validated by ELISA by time of APN treatment at different time points. (c) CXCL1 level of medium from SKOV3, SKOV3/control-siRNA, and SKOV3/VEGF-siRNA after 48 h treatment with or without APN.
We confirmed that APN treatment increased the CXCL1 level in the cultured medium from SKOV3 cells using ELISA (Figure 3(b)). The significant increase in the CXCL1 level affected by APN was shown after 48 h (P < 0.01). After 48 h of APN treatment, the CXCL1 level was measured in the medium from the SKOV3, SKOV3/control-siRNA, and SKOV3/VEGF-siRNA cells. The CXCL1 levels were significantly increased in the cultured medium from the SKOV3, SKOV3/control-siRNA, and SKOV3/VEGF-siRNA cells after APN treatment than were those without (P < 0.05). This finding suggests that APN mediates CXCL1 overexpression irrespective of VEGF knockdown (Figure 3(c)). In order to confirm that these cascades were mediated by the interaction between APN and its receptor, ADIPOR1 was downregulated by transfecting the SKOV3 cells with ADIPOR1-siRNA. We later confirmed that the CXCL1 level was lower in the transfected SKOV3 cells than it was in those with control-siRNA. However, the statistical significance was not reached in the ADIPOR1-siRNA 200 pg cells (P = 0.056; Figure 4).
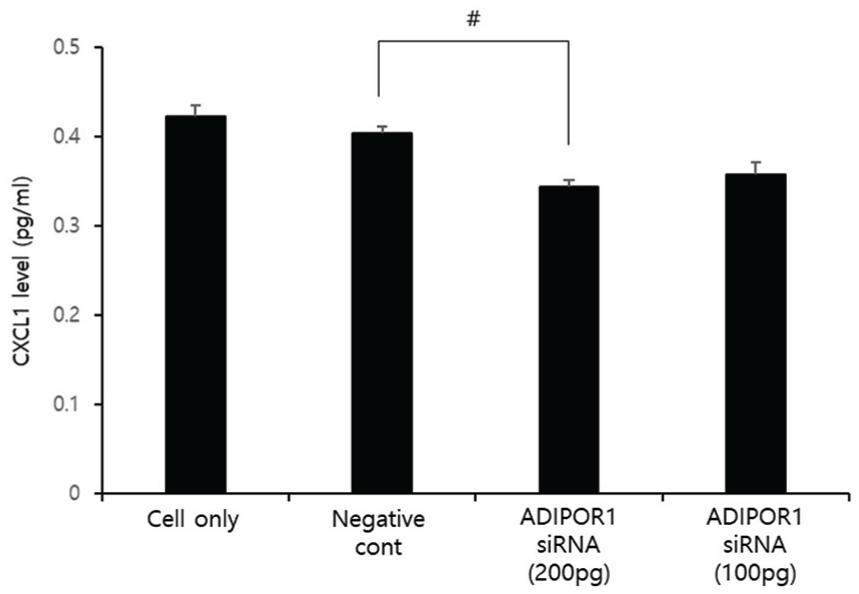
Blocking APN receptor attenuates CXCL1 secretion.
Effect of APN on ovarian cancer cell migration and invasion
Compared to the control, cells that were treated with APN (1 µg for 48 h) demonstrated a significant increase in APN-induced migration (287.7 ± 13.7 vs 226.3 ± 6.5; P < 0.01; Figure 5(a) and (b)). Similarly, the treatment of SKOV3 cells with APN also led to an increase in cell invasion compared to that of controls (307.7 ± 6.5 vs 207 ± 17.6, P < 0.01; Figure 5(c) and (d)). Together, these results suggest that APN plays an important role in tumor progression in vitro.

Effects of APN treatment on migration and invasion of SKOV3 cell lines: (a) transwell migration assays and (c) invasion assays shown at ×100. (b) Numbers of migrated and (d) invaded SKOV3 cells (1 × 105 cells/mL) after 48 h of treatment with APN 1 µg.
Discussion
EOCs have poor prognosis and remain the most lethal cancers in women. Angiogenesis is the most important step in tumor progression. It is therefore important to understand its components. APN appears to play an important role in the development and progression of malignant tumors, including gynecologic cancers. 20 This study demonstrates that APN has a pro-angiogenic effect on ovarian cancer. APN promoted ovarian cancer cells to secrete CXCL1, which stimulates angiogenesis independently of VEGF. To the best of our knowledge, our study is the first to suggest an angiogenic role of APN in ovarian cancer cells.
Prior studies were controversial regarding APN’s role in angiogenesis.11,17,18,21,22 APN induced the senescence of stromal cells and activated p16 and p53 signaling pathways, which promote angiogenesis. 23 In chondrosarcoma, one prior study showed that APN promoted angiogenesis through the PI3K, Akt, mTOR, and HIF-1α pathways, all of which induce VEGF-A expression. 24 Furthermore, another study of an APN knockout mouse model of mammary cancer had reduced vascularization with increased hypoxia and apoptosis, which are consistent with our results. 22 However, several different studies have found that APN inhibit angiogenesis. For instance, one group found that APN treatment downregulated VEGF and attenuated microvessel density and tumor growth in an orthotopic liver tumor nude mice model. 21 Another study found that APN suppressed tumor growth and angiogenesis both in vivo and in vitro in colon cancer. 25 APN increased the apoptotic cells of cleaved caspase-3 and inhibited the following angiogenic factors: CD31, VEGFb, and VEGFd. These inconsistencies in the literature regarding APN’s angiogenic role suggest that it has a complex influence on cancer cells that depends on the tumor environment and cell type. The discrepancy between studies may have also been caused by different protocols and materials.
Two types of ADIPOR forms have shown to have different distributions and affinities for the circulating APN. 26 In our results, ADIPOR1 was expressed higher than ADIPOR2 in ovarian cancer cell lines. ADIPOR1 is affluent in skeletal muscle and endothelial cells, while ADIPOR2 is abundant in the liver. 20 Therefore, we proved the role of ADIPOR1 as a precursor of angiogenesis, since ADIPOR1 was supposed to have a role in ovarian cancer cell lines. APN-induced pro-inflammatory cytokines such as IL-6, while it inhibited protective growth factors such as epidermal growth factor or fibroblast growth factor; this combination of effects likely contributes to tumorigenesis. 27 Physiological angiogenesis is essential for embryo development, wound healing, and organ perfusion. In contrast, abnormal angiogenesis plays a crucial role in tumor progression. 28 There are various well known stimulators of angiogenesis, as follows: VEGF, transforming growth factor (TGF)-α, TGF-β, granulocyte colony-stimulating factor, fibroblast growth factor, and plasminogen activator inhibitor-1. 29 VEGF has been demonstrated to be a major contributor to angiogenesis. 30 In this study, APN treatment had no effect on tube formation. This lack of effect can be explained by the fact that VEGF is one of the main pro-angiogenic factors in ovarian cancer. We therefore suppressed VEGF by transfection with VEGF-siRNA in order to determine the exact role of APN in angiogenesis in ovarian cancer cells. This is the first in vitro study to confirm the pro-angiogenic effect of APN in ovarian cancer, especially in the absence of VEGF’s effect. Thus, APN may promote tumor progression through enhanced angiogenesis.
This study characterizes the effect of APN on CXCL1 expression in SKOV3 cells. SKOV3 cells treated with APN demonstrated CXCL1 augmentation in a time-dependent manner. During a woman’s reproductive years, there is continuous ovarian cycling, which triggers repetitive wound healing and repair. This pathway is associated with an inflammatory response and chemokine secretion. 31 CXCL1 is one member of the CXC chemokine family that promotes angiogenesis, and secretes pro-angiogenic factors like VEGF. 32 In previous studies regarding CXCL1 expression in patients with EOC, the CXCL1 was elevated in the ascitic fluid as well as serum and tissue from patients. 33 In addition, CXCL1 has also been suggested to be a useful serum biomarker in ovarian cancer.34,35 CXCL1 binds to the CXC chemokine receptor 2 (CXCR2), which induces the angiogenic activity of CXCL1. CXCL1 was found to induce endothelial migration and proliferation in EOC, which leads to metastatic spread. 36 In addition, CXCL1 can secrete pro-angiogenic factors like VEGF. 32 However, we could not find that the VEGF expression increased following APN treatment (Supplementary Figure S1).
CXCL1, a CXCLR2 ligand, has prompted cell migration and invasion in ovarian cancers. Previous report suggested that CXCL1 was related to nuclear factor-κB (NF-κB) signaling. 37 CXCL1 binding to CXCR2 in ovarian cancers induced cell proliferation, migration, and invasion through NF-κB activation. For instance, NF-κB inhibitor attenuated cell migration and invasion in CXCR2-positive ovarian cancer cells. Furthermore, NF-κB was activated through epidermal growth factor receptor (EGFR)-transactivated Akt signaling. 37 Blocking CXCR2 decreased EGFR activity, and Akt blocker attenuated NF-κB promoter activity. Therefore, NF-κB is likely to play an important role in the CXCL1-mediated angiogenesis by APN. In addition, phospho-signal transducer and activator of transcription 3 (p-STAT3) and phosphor-Janus kinase 2 (p-JAK2) were also involved in the induction of migration and invasion by CXCL1 in gastric cancer. 38
Plasma APN levels are lower in obese women than they are in non-obese women. 9 The expression of ADIPORs is also suppressed in obese women than that in non-obese women. 39 Therefore, the carcinogenic effect of APN identified in this study may be more activated in non-obese patients with ovarian cancer than it is in obese cancer patients. Therefore, inhibitory agents that work against APN or ADIPOR are expected to have more prominent anti-tumorigenic effects on non-obese ovarian cancer patients than on obese patients. The ADIPOR levels on tissue array and immunohistochemistry correlated with cancer prognosis, including overall survival and disease-free survival. 40 ADIPORs are also more highly expressed in patients with progressive courses, such as cases of deep invasion and lymph node metastasis. Although prior studies have not addressed the association between the ADIPOR expression and prognosis in ovarian cancer, its upregulation was associated with poor survival in colorectal cancer. 41 The correlation between ADIPOR expression and tumor aggressiveness has also been demonstrated in esophageal and breast cancer.42,43 Further studies are needed to clarify the predictive value of the ADIPOR in EOC and to identify therapeutic target against APN.
This study is the first to identify the pro-angiogenic role of APN in ovarian cancer cells. It is important to explore the new target for therapeutic drugs. We also investigate the migratory and invasive properties of APN, which are associated with the metastatic characteristics of ovarian cancer. These suggest that suppressing APN or the ADIPOR could prevent tumor progression and metastasis in ovarian cancer. Previously, APN-induced migration and invasion have also been shown in prostate and breast cancer cells.44–46 Regardless, our study has several limitations. First, we did not identify the pro-angiogenic function of CXCL1 or CXCR2 by blocking CXCL1 or CXCR2. However, the binding of CXCL1 to CXCR2 has trophic and angiogenic properties in tumor cells in previous studies, and CXCR2 is the major chemokine receptor mediating angiogenic response.32,33,35,36,47 More in vivo experimentation is needed to clarify the role of APN on angiogenesis. Further clinicopathologic investigation using ovarian cancer tissue samples derived from patients of APN and ADIPOR is needed.
Our results cannot directly explain why obesity, which is characterized by low plasma APN levels, increases the risk for various cancers. Still, there are controversial findings regarding APN’s role in angiogenesis. Therefore, further investigation is needed to establish the role of APN in the relationship between obesity and carcinogenesis.
Conclusion
APN stimulates CXCL1 secretion from ovarian cancer cells, which promotes angiogenesis irrespective of VEGF. These results suggest that APN is a key molecule in the initiation of angiogenesis in ovarian cancer cells. Therefore, it may be a novel therapeutic target in the treatment of ovarian cancer. These findings provide a better understanding of angiogenesis and can contribute to the development of new therapies for ovarian cancer. Nevertheless, further studies are needed to understand the role of APN in the relationship between obesity and ovarian cancer. We must also further investigate the clinical relevance of the APN.
Supplemental Material
Supplementary_figure – Supplemental material for CXC chemokine ligand 1 mediates adiponectin-induced angiogenesis in ovarian cancer
Supplemental material, Supplementary_figure for CXC chemokine ligand 1 mediates adiponectin-induced angiogenesis in ovarian cancer by Yung-Taek Ouh, Hyun Woong Cho, Jae Kwan Lee, Song Hee Choi, Hyun Jin Choi and Jin Hwa Hong in Tumor Biology
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a Korea University Grant.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.