
Abstract
Some surface markers are used to discriminate certain leukemic subpopulations that retain a greater oncogenic potential than others, and, for this reason, they were termed as leukemic stem cells, similar to the concept of cancer stem cells in carcinoma. Among these surface markers are proteins involved in cell–cell adhesion or cell–matrix adhesion, and they may play a role in the relapse of leukemia, similar to metastasis in carcinomas. The most important are epithelial cadherin, neural cadherin, epithelial cell-adhesion molecule, and CD44, which can be cleaved and released, and their soluble forms were found increased in serum levels of cancer patients, being implicated, in some cases, with progression, metastases, and relapse. In this review, we highlighted the role of these four adhesion molecules in carcinomas and hematological malignancies, mainly leukemia, and discuss if the serum levels of soluble forms can be correlated with the surface protein status on the leukemic cells. Accession of the soluble forms looks attractive, but their use as markers in cancer must be studied in association with other parameters, as there are significant changes in levels in other pathological conditions besides cancer. Studies correlating the levels of the forms with the status of the membrane-bound proteins in leukemic (stem) cells and correlating those parameters with relapse in leukemia may afford important knowledge and applicability of those serum markers in clinical practice. For instance, the expression of the membrane-bound forms of these adhesion proteins may have promising clinical use in leukemia and other hematological malignancies.
Introduction
Several cell-surface proteins are important in cancer, and hematological malignancies are not an exception. These cellular components are used for immunophenotyping, a procedure critical for diagnosis and treatment of leukemia and other hematological disorders. For example, the blast cell count (CD34+ cells) is critical in leukemia treatment, and monitoring that parameter serves as an indicator of active disease, minimal residual disease, and remission. 1 Some patients, considered to be in morphological remission, had in excess of 5% leukemic blasts. Those patients considered resistant disease have more than 15% blasts. Minimal residual disease (MDR) is considered if patients present less than 1% of blasts. 2 However, other surface markers are important for clinical use, as some attribute different properties to leukemic blast cells, like CD123, and/or are lineage markers, like CD19. 3 Several surface markers can be helpful for therapy strategy choice.4,5
In that trend of thought, some surface markers, such as CD123, lineage markers (CD2, CD5, CD7, CD11b, CD11c, CD19, CD22, CD56), C-type lectin-like molecule-1 (CLL-1), CD96, CD47, and T-cell immunoglobulin and mucin-domain containing-3 (TIM-3), are widely used to discriminate certain leukemic subpopulations that retain a greater oncogenic potential than others, and, for this reason, they were termed leukemic stem cells (LSCs), like the concept of cancer stem cells (CSCs) in carcinoma. In a general way, these LSCs retain several characteristics of normal hematopoietic stem cells (HSCs), such as the potential of generating “mature” leukemic cells. Notably, some studies found that mature leukemic cells can recapitulate stem properties, asserted by surface markers related with normal HSCs, like CD34 and eventually CD38. However, other surface markers besides those related to HSCs contribute to that oncogenic property. 3
Some of these surface markers are proteins involved in cell–cell adhesion or cell–matrix adhesion. For example, CD34 participates in adhesion of HSCs with bone marrow matrix and stromal cells. 6 HSCs interact with several stromal cells and extracellular matrix components in the bone marrow, and it is not different with leukemic cells, mainly those that retain stem cell–like properties.3,7 For example, CD44 is involved in bone marrow homing and also with stem cell properties in acute myeloid leukemia, and its depletion by antibodies resulted in the clearance of medullary and free leukemic cells in graft mice models. 8
There are some organs and tissues, such as the central nervous system and bone marrow, that treatment-resistant leukemic cells are set and permanent in quiescence, rising after completion or in the interval of an aggressive treatment (relapse).1,9 In this matter, some of these adhesion molecules may play a role in relapse of leukemia,3,7 similar to metastasis in carcinomas.
The most important and well-studied adhesion molecules that play a role in metastases are epithelial cadherin (E-cadherin), neural cadherin (N-cadherin), epithelial cell-adhesion molecule (EpCAM), and CD44. These membrane proteins exert critical roles in cell–cell or cell–matrix interaction as well as differentiation, motility, and metastasis.10–17 Those surface markers can be cleaved and released,13,18–20 and their soluble forms were found increased in serum levels of cancer patients,19,21–28 being implicated, in some cases, with progression and metastases. 19 Moreover, the soluble forms of some of these surface markers increase in the serum of patients with hematological malignancies, such as leukemia.29–34
As the source of the soluble forms are the membrane-bound proteins, one question can be raised: can the serum levels of soluble forms be correlated with the surface protein expression on the leukemic cells? In this review, we highlighted the role of these four adhesion molecules in carcinomas and hematological malignancies, mainly leukemia, and discuss evidence about this matter.
In the first topic, we will briefly describe the biology of the four cell-adhesion molecules (CAMs) and biogenesis of the soluble forms. The second topic will be divided into two subtopics: one for carcinomas, briefly describing the role of membrane forms, then their soluble forms in tumor progression and metastasis and the second subtopic will focus on their role in leukocyte malignancies, mainly leukemia, and thereafter the findings about their soluble forms in hematological malignancies will be presented. The third topic will discourse about their non-specific role, like inflammation, and discriminate between general processes and cancer. Finally, in the fourth topic, we will resume the important data for this review and propose a hypothetical use of the soluble forms in leukemia.
Biology and biogenesis
E-cadherin is a member of the cadherins (calcium-dependent adhesion integrins) family, formerly found abundant in epithelial tissues, with an important role in cell–cell adhesion, like desmosomes and adherents junctions. It interacts with cadherin residues of other cells to establish homophilic interactions. 35 E-cadherin also perform heterophilic interactions with immune cell through their E-cadherin ligands (CD103, killer cell lectin-like receptor subfamily G member 1 (KLRG1)), inhibiting immune cell activation and proliferation. 35 Cadherins have important role in cell architecture by indirect interaction with cytoskeleton through their cytoplasmic (amino-terminal) domain. The cytoplasmic domain directly binds either β-catenin or γ-catenin/plakoglobin that, in turn, mediate interaction with actin. In addition, p120 is another catenin member that interacts with the juxta membrane region of E-cadherin, strengthening its adhesive properties. 36
In another perspective, the E-cadherin cytoplasmic domain acts as a docking area for β-catenin, thus repressing the proliferative function of β-catenin 37 in an adhesion-independent manner. 38 Loss of E-cadherin leads to β-catenin release, promoting cell proliferation, gain of motility, and other characteristics typical of a migrating cell. 35 In addition to the well-known role in inhibiting β-catenin transcriptional properties, E-cadherin also modulates phosphatidylinositol 3-Kinase (PI3K)/protein kinase B (Akt) signaling pathway, Ras homolog guanosine triphosphate (Rho GTPase) and inhibits nuclear factor κB (NF-κB) activity. 35
N-cadherin is another member of the cadherins family. Similar to E-cadherin, N-cadherin plays a role in cell–cell adhesion, like desmosomes and adherents junctions of mesenchymal and neural cells and other non-epithelial cells. 39 Its cytoplasmic domain, similar to E-cadherin, interact with catenin members, impacting cell architecture through indirect interaction with cytoskeleton. 36 Similar to E-cadherin, N-cadherin represses the pro-proliferative properties of β-catenin in several cell types. 39 However, in other cells, it was observed that N-cadherin interact with Wnt co-receptor LRP6, activating the Wnt/β-catenin pathway, and promote Akt activation, impeding β-catenin degradation, 40 that, strikingly, occurs the opposite in other cell types. 41
The soluble forms of these cadherins, that is, the extracellular domain of them is released by proteolytic cleavage activity of sheddases, like matrix metalloproteases (MMPs), membrane-type matrix metalloproteases (MT-MMPs), a disintegrin and metalloproteinases (ADAMs), kallikrein, plasmin, and other sheddases. 18
By intuition, cleavage of E-cadherin and N-cadherin could release β-catenin, leading to increased cell proliferation and migration because they act as cytoplasmic docking of β-catenin. 42 However, it was observed a counterintuitive role regarding this matter. It was observed that their intracellular domain inhibits β-catenin signaling. 43 Also, it was observed that N-cadherin intracellular domain inhibits β-catenin but abolishes contact-dependent inhibition of cell proliferation and promotes anchorage-independent cell growth by modulating other pathways. 44 It is important to state that these studies were performed by ectopically expressing a construct of the cytoplasmic domain only and can differ from the physiological cytoplasmic release (proteolytic cleavage). They can be intracellularly cleaved by caspase 3 or presenilin, releasing their cytoplasmic domain, and can be extracellularly by ADAM10, for example, releasing their extracellular domain. Interestingly, ADAM10, that is, extracellular cleavage of E-cadherin and N-cadherin leads to release and activation of β-catenin signaling. 42 Yet, the proteolytic pathway of these cadherins, although could result in increased cell proliferation, has several differences regarding cell signaling. In accordance, presenilin could favor degradation of β-catenin. However, it was also observed that presenilin-mediated proteolysis of N-cadherin lead to nuclear translocation and transcriptional activity of β-catenin. 42 Further discussion in this article will focus in extracellular proteolytic cleavage, as it releases the soluble form of the four cell-adhesion molecules presented.
EpCAM is another abundant, calcium-independent, transmembrane glycoprotein that exerts homophilic cell–cell interaction in epithelial cells. 45 Recent studies observed that EpCAM is also involved in the β-catenin/Wnt (wingless-type MMTV integration site family) signaling pathway;46–48 cleavage of EpCAM by ADAM or presenilin leads to cell signaling by an EpCAM intracellular domain/FHL2 (four and a half LIM domains 2)/β-catenin complex, 12 that result in cell proliferation,49,50 and release of the extracellular domain. Although EpCAM is an adhesion molecule, it could act as a negative dominant of E- and N-cadherin on cell–cell adhesion11,12 and has an important role in cell motility. 10 For a review, refer the study performed by Schnell et al. 51
CD44 is a transmembrane glycoprotein, and, different from previous, is involved in cell–extracellular matrix interaction. Its extracellular domain interacts can recruit matrix metalloproteinases to cell surface, that, in turn, cleaves cell surface and extracellular proteins, such as pro-form of transforming growth factor (TGF)-β, type IV collagen, and others. However, the most known interaction is with hyaluronic acid (HA). The intracellular domain is responsible of interactions with cell skeleton proteins, and, thus, can regulate cell shape and migration. 52
By far, the most notorious roles of CD44 in cell migration is leukocyte recruitment and trans-endothelial migration. Lymphocytes use CD44-HA interactions to roll to inflamed sites. In contrast, neutrophils can firmly adhere by CD44-HA interaction. Several studies demonstrated that CD44 or HA impairment led to decrease leukocyte recruitment to disease. 53 For more details about CD44 role in cellular biology, see Chen et al. 54 Its cleavage by membrane type 1 matrix metalloproteases (MT1-MMP) and presenilin-1/y-secretase leads to extracellular domain release (soluble CD44) and translocation of the intracellular domain that acts as a transcription factor, increasing expression of CD44 itself, inflammation, survival, and stem markers, depending on context. 13
Roles in cancer
Solid cancer
Membrane-bound proteins
Epithelial-to-mesenchymal transition (EMT) is the most important mechanism that allows epithelial malignant cells to migrate, leading to distant establishment of a secondary tumor (metastasis). During this cell reprogramming process, epithelial cells, which are naturally attached to their tissue, lose their epithelial markers, like E-cadherin, and gain mesenchymal characteristics, like N-cadherin expression, being able to detach from tissue and migrate to a distant site where they settle and return to an epithelial state by the reverse event, mesenchymal-to-epithelial transition (MET). Several transcription factors regulate EMT and MET.14,16 Although traditional EMT is recognized as loss of E-cadherin and gain of N-cadherin, co-existence of E-cadherin and N-cadherin, by only N-cadherin upregulation, occurs in a process named partial EMT. 15
EpCAM, first known as epithelial specific antigen (ESA), was first described upregulated in colorectal cancer with clinical relevance. 55 EpCAM+ cells have a greater potential to initiate tumor engraftment. Al-Hajj et al. 56 demonstrated that breast CSCs (CD24−/CD44+) EpCAM+ cells are capable of inducing tumors in mice, different than their EpCAM− counterparts. These findings apply to other cancer tumor-initiating cells such as colorectal, ovarian, and hepatocellular carcinoma CSCs.57–60 Also, CSCs EpCAM expression has been associated with therapy resistance and tumor bulk maintenance.56,58,59,61 Importantly, EpCAM is one of the most reliable markers of circulating tumor cells (CTCs) in various carcinomas, and the presence of EpCAM+ CTCs is related to poor outcome, like colorectal cancer, head and neck cancer, ovarian cancer, and breast cancer.62–67
CD44 is another CAM abundant in CSCs13,17,68,69 and is involved in cell motility, invasion, angiogenesis, and metastasis.13,17 Its intracellular domain that remains after extracellular domain cleavage plays an important role in cell signaling to promote metastasis. 13 CD44 is widely expressed as the common (standard) isoform (CD44s), but it has several variants (CD44vs) that interact with different growth factors. For example, CD44v3 binds to fibroblast growth factor 2 (FGF2), and CD44v6 can bind to vascular endothelial growth factor (VEGF), osteopontin (OPN), and hepatocyte growth factor (HGF). These properties of CD44 variants have a great impact in pro-tumoral features. 70 Interestingly, CD44s and CD44vs have particular roles in different cancers.13,69
Therefore, these CAMs have important roles in carcinoma progression. E-cadherin loss is the most common event in cancer progression, but increases in N-cadherin, EpCAM, and CD44, independent of increases in E-cadherin, are of greater importance, as these three molecules are involved with invasion, motility, and stemness.
Soluble proteins
Soluble epithelial cadherin (sE-cadherin) plays an important role in cancer progression, since several signaling pathways associated with tumor progression are activated, like epidermal growth factor receptor (EGFR) family members (EGFR and human epidermal growth factor receptor-type 2 (HER2)) and insulin-like growth factor 1 (IGF-1R), thus activating mitogen-activated protein kinase (MAPK)/extracellular signal–regulated kinases (ERKs) and PI3K/Akt signaling. Also, the antitumor signaling pathway Hippo is inactivated. sE-cadherin changes cytotoxic cell activity by binding to KLRG1 of natural killer (NK) and TCD8+ cells, maintaining the cells in an inactive state 19 by triggering the inhibitory signaling of KLRG1 similar to membrane-bound E-cadherin. 35 Interestingly, sE-cadherin levels have a positive correlation with cancer progression, with higher levels in advanced/metastatic states compared to initial stages and decrease in breast cancer patients after chemotherapy. 19 Moreover, increases in sE-cadherin and their levels serve as predictive of relapse in prostate and bladder cancer.71,72 This corroborates with the hypothesis that tumors may be the main source of sE-cadherin. Importantly, sE-cadherin inhibits the adherent role of E-cadherin in cell–cell adhesion by binding with E-cadherin, dislocating membrane-bound E-cadherin homophilic interaction between two cells.18,19 So, even if primary tumors express high levels of E-cadherin that indicate less chance of metastasis, the association with higher levels of sE-cadherin can be associated with elevated chances of metastasis by all signaling that it triggers and by inhibiting E-cadherin homophilic interaction. 19
Soluble neural cadherin (sN-cadherin) stimulates migration of endothelial cells, angiogenesis via fibroblast growth factor receptor (FGFR) activation, neurite outgrowth, and perineural invasion in prostate cancer. 18 Increased levels of serum sN-cadherin in patients with bone sarcoma disease and soft tissue sarcoma, compared to healthy controls, were also observed and correlated with tumor stage and grade, respectively. 24 Although scarce knowledge exists about sN-cadherin in tumor progression, the information suggests an important role for mesenchymal and endothelial cells in angiogenesis and invasion.
Regarding soluble epithelial cell-adhesion molecule (sEpCAM), although the intracellular domain (EpICD) has the main role of EpCAM in cancer, the extracellular domain of EpCAM/soluble form (EpEX) enhances the cleavage of EpCAM, leading to intracellular signaling by EpICD. 12 In accordance, elevated sEpCAM levels in ascites associated with ovarian tumor were correlated with positive cytology for malignancy. 26 It was associated with shorter overall survival in patients with ovarian cancer treated with catumaxomab, a bispecific antibody that targets EpCAM. 25 Elevated urinary sEpCAM concentrations were observed in bladder cancer and found as an independent indicator of poor prognosis, 22 and elevated serum concentrations were also observed in epithelial ovarian cancer. 27
Proteolytic cleavage of CD44 is important for cell signaling by its intracellular domain. 13 Yet, the importance and biological functions of the soluble form (extracellular domain) of CD44 (sCD44) is not known. However, correlations between serum levels of sCD44 and tumor progression in colorectal cancer 28 and overall survival in HER2-overexpressing breast cancer patients 21 and in pre-malignant lesions of cervical cancer 23 were observed.
Therefore, although not well elucidated, the soluble forms of E-cadherin, N-cadherin, EpCAM, and CD44 play important roles in cancer progression and metastasis, and some of them act as ligands of several receptors involved in this process. Moreover, several studies demonstrated that these soluble forms originate from the tumor tissue and are suggestive of tumor status and progression. Thus, it is reasonable to hypothesize that something similar occurs in hematological malignancies.
Hematological malignancies
Membrane-bound proteins
Regarding hematological diseases, E-cadherin can be modulated by genetic and epigenetic events in leukemias. CDH1 (E-cadherin coding gene) is widely repressed by epigenetic alterations (methylation), leading to excess of leukemic blast cells and poor prognosis in acute lymphoblastic leukemia (ALL), chronic myeloblastic leukemia (CML), acute myeloblastic leukemia (AML), and acute promyelocytic leukemia (APL)73–76 and has a trend of correlation with N/K-RAS mutations in AML. 75 Concordantly, restoration of E-cadherin expression (with inhibitors of histone deacetylases) in leukemic cells led to cell homophilic adhesion 77 and downregulation of proliferation markers. 78 Interestingly, different from previously mentioned leukemias that repress E-cadherin expression, it was observed that erythroid AML expresses surface E-cadherin. 79
It was observed that N-cadherin is important for LSC homing to the marrow niche. 80 In concordance, in AML, N-cadherin+ cells were enriched in the CD34+ cell compartment (LSCs), retained stem cell–like properties, and had greater oncogenic potential compared to N-cadherin− (CD34+ LSCs) cells. 81 Also, N-cadherin is the most abundant cadherin expressed by T-cell lymphomas and other leukemic lineage cells, promoting homophilic N-cadherin interaction with mesenchymal cells. 82
One study observed an increase of cytoplasmic stabilized β-catenin in B acute lymphoblastic leukemia (B-ALL) with (1;19) (q23;13) translocation. Interestingly, it was observed that β-catenin was docking in the N-cadherin intracellular domain, increasing N-cadherin levels and reinforcing the interaction between leukemic cell and bone marrow stroma. 20
In another perspective, the expression of E-cadherin and N-cadherin by stromal cells have an important role in hematological malignancies. In mice, CDH1 and CDH2 (N-cadherin coding gene), as well as their coding proteins, are expressed by epithelial and mesenchymal thymus stromal cells, respectively. It was observed a biphasic expression pattern of expression of CDH1 and CDH2, as well as their proteins, in thymic stromal cell of mice models of thymic lymphoma: during acute gamma-irradiation, an upregulation is observed before establishment of lymphoma, but, when efficiently established, a strong loss of their expression is observed. 83 Because human thymic stromal cells exert heterophilic interaction with CD103+ thymocytes through E-cadherin, resulting in a positive regulation of cell proliferation, 84 De Yzaguirre et al. 83 suggest a pro-tumoral role of stromal cadherin upregulation at initial stages, with downregulation during lymphoma progression. These data altogether suggest an important role of cadherin expression not only by hematologic cells but also by stromal cell during progression of hematological malignancies.
The scientific literature presents a few studies regarding EpCAM in hematological malignancies. One study demonstrated that EpCAM is frequently overexpressed in leukemic cells of AML patients, compared to healthy bone marrow cells, and these cells exhibit enhanced chemoresistance and oncogenesis. They found that anti-EpCAM antibody depleted subcutaneous, disseminated, and intramedullary engrafted mice with AML cells, suggesting that this adhesion molecule may be used as a therapeutic target. Interestingly, it was observed that EpCAM is enriched in AML LSCs (CD34+) and EpCAM+ cells co-express several LSC markers, like CD123 and CD44. Importantly, it was observed that EpCAMhi AML cells also express high levels of CD44. 85
CD44v are widely expressed in stem cells, like HSCs, in stark contrast to differentiated cells. Leukemic cells, mainly LSCs, overexpress CD44 in resemblance to HSCs but mostly the variant isoforms. 70 As mentioned previously, CD44, mainly CD44v, bind to several growth factors, and this characteristic supports leukemic cell apoptosis resistance, drug resistance, and proliferation. 70 One study observed that CD44 plays an important role in the establishment of leukemic cells in AML. A specific antibody against CD44 was used to treat leukemia in mice models. They demonstrated that mice treated with the antibody did not develop the tumor compared to mice not treated with the antibody, and the medullary portion did not present with any leukemic cells. Also, they observed that CD44 is enriched with LSCs. 8 Although being observed in gastrointestinal lymphoma, it is interesting that weak CD44 expression was associated with better prognosis than strong CD44 expression. 86
It can be appreciated that these adhesion molecules in hematological malignancies have roles similar to carcinomas. For example, E-cadherin loss in AML led to increases in blasts, but its expression is of concern in histiocytosis, plasmacytomas, and malignancies of leukocytic origin that allocate in epithelial tissue, as might be expected by their adherent nature. Also, N-cadherin is involved with homing to bone marrow (mesenchymal tissue), like the metastasis process. And, finally, CD44 and EpCAM are linked with stemness, motility, and therapy resistance in leukemias. These data demonstrate a great resemblance with carcinomas (Figure 1).
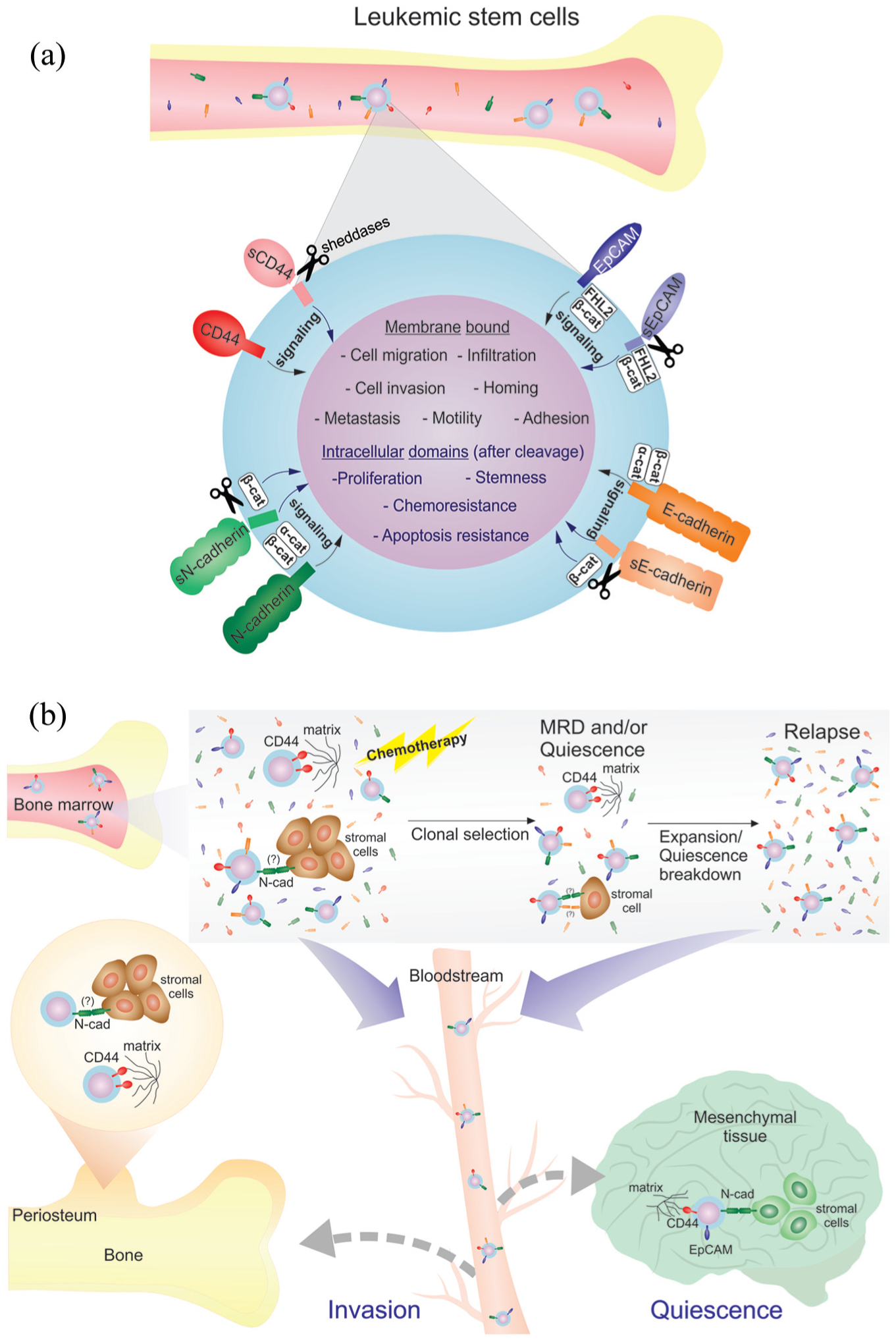
Role of E-cadherin, N-cadherin, EpCAM and CD44 in leukemic progression. (a) Interaction of adhesion molecules evokes several cell signaling related to aggressiveness, like invasiveness and migration in leukemic cells (black arrows). Proteolytic cleavage leads to increase of the soluble forms of adhesion molecules, participating in disease progression by acting as ligand and releasing the intracellular domain of these domains, evoking different cell signaling (gray arrows). (b) Evolution of leukemic disease and hypothetical role of adhesion molecules in progression. At primary disease, therapy eventually lead to selection and expansion of resistant clones (relapse), with de novo increase of serum forms of adhesion molecules, and/or pressure to quiescence. Adhesion molecules may play important role allowing homing and invasion to extramedullary tissue, cell–cell and cell–matrix adhesion, until quiescence breakdown and relapse.
Soluble protein
sE-cadherin seems to participate in leukemia development. Patients with ALL, acute non-lymphocytic leukemia, CML, myelodysplastic syndrome (MDS), and non-Hodgkin lymphoma (NHL) presented with increased serum sE-cadherin than healthy controls and were positively correlated with stages and progression. An important finding is that serum sE-cadherin in blast crisis of CML is higher than in the chronic phase. 34 Another study observed similar findings in a cohort of MDS patients: elevated serum sE-cadherin correlates with poor prognosis. 30
Moreover, in newly diagnosed multiple myeloma (MM), high serum sE-cadherin levels (~six-fold) were observed compared to healthy controls and correlated with lactate dehydrogenase (LDH) levels, a non-specific tumor serum marker that was found as an independent prognosis factor of survival. 31
These data suggest that sE-cadherin may be produced from malignant hematological cells and in accordance with the abundance of these cells. Also, it may be associated with proliferation and extramedullary infiltration of malignant cells.30,31,34
Increases in serum sCD44 were observed in pediatric ALL, independent of lineage, and decreased with treatment and remission to levels similar to that of healthy controls, without correlation between sCD44 and LDH levels, but had a weak correlation with peripheral leukocyte counts. 33 In addition, increases in serum sCD44 levels were correlated with Hodgkin’s disease (HD), NHL, Burkitt’s lymphoma (BL), and ALL, but not AML, compared to healthy controls, with decreases to normal values in complete remission. There was positive correlation with stage and poor prognosis in HD and NHL, and positive correlation between serum sCD44 levels with tumor tissue CD44 levels in HD and BL. 32
Another study observed similar levels in serum sCD44 in newly diagnosed children and adults with B-ALL compared to healthy controls. However, sCD44 was higher in patients with white blood cells (WBCs) >50 × 109/L than patients with WBC <50 × 109/L and in patients with a peripheral blast percentage >50 compared to patients with a peripheral blast percentage <50, presence of splenomegaly, extramedullary involvement, and in some cytogenetic conditions. 29 Elevation in serum sCD44 levels was detected in patients with MDS compared to healthy controls and correlated with poor prognosis. 30
These data suggest that sCD44 has its origins in tumors by proteolytic cleavage of the CD44, making it a potential reliable biomarker of clinical monitoring and prognosis marker in hematological diseases.29,32
As far as we know, no data are available about serum levels of sN-cadherin and sEpCAM in hematological malignancies.
Despite scarce data, the soluble forms of E-cadherin and CD44 are indicative of active disease in hematological malignancies and play roles in progression of these cancers.
Soluble proteins: noise, non-specific, or something else?
The soluble forms of these adhesion molecules are mostly studied in cancer by their potential role as biomarkers, because most of them are derived from tumor tissue surface proteins. However, an important question arises: can their serum levels vary in other diseases? As an example, bacterial proteases can cleave E-cadherin, producing sE-cadherin. 87 Thus, its “unspecific” role must be elucidated.
Elevated serum sE-cadherin levels was observed in bacterial and viral infections, organ failure, and benign tumors, and increased levels were observed in psoriasis, acute pancreatitis, diabetes, diabetic nephropathy, HIV infection, and sepsis. 87
It is important to point out that echinococcosis alters hepatic parenchyma that could be misinterpreted as hepatic cancer. One study observed lower plasma sE-cadherin concentrations in patients with echinococcosis compared to healthy controls, with a positive correlation between sE-cadherin levels and CDH1 expression by quantitative polymerase chain reaction (qPCR) from leukocytes. In that study, no difference was observed in sN-cadherin plasma concentrations of patients with echinococcosis. In this scenario, the authors suggest sE-cadherin as a potential differential marker between echinococcosis and hepatic cancer, which has elevated sE-cadherin levels. 88 These data are of importance because some cases of echinococcosis clinically mimic hepatic malignancy. 89
The adhesion molecule sCD44 did not change in serum from patients with bacterial infection compared to healthy controls. 33 However, sCD44 levels increased in the acute phase of Trypanosoma cruzi infection compared to healthy controls, but levels decreased in the chronic phase with advanced cardiopathy compared to healthy controls and patients with milder disease. 90
Recently, it was observed that sCD44 increases in hepatic steatosis 91 and chronic and aggressive periodontal disease, 92 and its levels correlate with the evolution of endometriosis. 93
One study analyzed sEpCAM in endometriosis, and did not find any correlation. 94
Thus, the soluble forms of these adhesion molecules suffer significant changes in other pathological conditions besides cancer.
Leukemic relapses: “metastasis” of white blood cell cancer?
As mentioned previously, these CAMs appear to have roles in hematological malignancies. Interestingly, some studies observed that the transcription factors that regulate EMT and MET in carcinomas have a role in hematological malignancies, including leukemia. Importantly, it was observed that CML LSCs are marked by “EMT” transcription factors. 95 Thus, it is not illogical that other similar gene control and cell reprogramming processes in carcinomas occur in leukemia. So, monitoring traditional surface markers of carcinomas may have applicability in leukemia. Importantly, as mentioned previously, EpCAM is one of the most reliable markers of CTCs in various carcinomas,62–66 and, intriguingly, it appears to have a similar role in AML. 85
In carcinomas, the preconized cancer progression has metastasis as “the final step.” As reviewed in this article, some processes are critical for this downregulation of E-cadherin and/or upregulation of N-cadherin with concomitant upregulation of CD44 and, eventually, EpCAM. These processes promote cell motility, invasion, and migration as well as cell adhesion with endothelial cells and mesenchymal tissue, allowing cell attachment in bone, lung, cerebral, and other tissues as well as stemness, therapy resistance, and general cell robustness.
Hiding of leukemic cells in “sanctuaries” can be related to the metastasis process of carcinoma, if looked at closely. However, quiescence of LSCs is a common mechanism involved in leukemic recurrence,96,97 that is rarely involved with metastasis in solid cancer, although quiescence is a property of CSC, cells that poses strong metastatic features. 98 Interestingly, it was observed that N-cadherins are important for quiescence and self-renewal in several adults stem cells. 99 E-cadherin, although a marker of epithelial (differentiated) cells, as well has a role in quiescence and self-renewal in adults stem cells, including neuronal stem cells. 99 EpCAM is widely expressed in tumor-initiating cells (also known as CSCs) that are almost quiescent (EpCAM and its potential role in tumor-initiating cells). CD44 is important for quiescence and differentiation in neuronal 100 and other stem cells 101 and leukemia as well. 70
Interestingly, it was observed that reduced β-catenin increases adhesiveness, quiescence, and cadherin expression, including CDH1 in HSCs. 102 As aforementioned, E- and N-cadherins and EpCAM can stabilize and inhibit β-catenin signaling. So, these adhesion molecules could have a role in hiding and quiescence of leukemic cells, thus being factors associated with recurrence. De novo expression of CAMs or halt of proteolysis of CAM may be involved in this process. In accordance with this trend of thought, degradation of several surface molecules, such as integrins, by MMPs from granulocyte-colony stimulating factor (G-CSF)-induced neutrophils lead to HSCs quiescence break and mobilization to bloodstream, which is also observed in xenograft models of leukemia. 96 In this context, the soluble forms of these adhesion molecules probably have predictive value in relapse and may have use for relapse monitoring in leukemia, as occured in several carcinomas.71,72
In this trend of thought, one study observed an association between the percentage of cells expressing CD44 and the occurrence of disease relapse in children with B-cell progenitor ALL lacking adverse or favorable genetics. 103 Importantly, CD44 was observed in extramedullary manifestations of myeloid leukemia. 104 These data support the hypothesis that these classic adhesion molecules with great importance in carcinoma metastasis have a role in leukemic relapse and invasiveness.
Concluding remarks
The adhesion molecules, mainly E-cadherin, N-cadherin, CD44, and EpCAM, may have similar roles in the infiltration and relapse of leukemia as their roles in the metastases of carcinomas.
Accession of the soluble forms looks attractive, since they are produced from the membrane-bound forms of leukemic cells. However, there are significant changes in levels in other pathological conditions besides cancer. For this reason, their use as markers in leukemia must be studied in association with clinical data, leukemia type, and specific mutations and translocations. Yet, studies correlating the levels of the forms with the status of the membrane-bound proteins in LSCs and correlating those parameters with relapse in leukemia may afford important knowledge and applicability of those serum markers in clinical practice.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Pro-Rectory of Research and Post-Graduation of the Federal University of Uberlândia funded the fee related with this publication.