
Abstract
A disintegrin and metalloproteinase 9 (ADAM9) possesses potent metastasis-inducing capacities and is highly expressed in several cancer cells. Previous work has shown that ADAM9 participates in the adhesive–invasive phenotype in lung cancer cells in vitro. In this study, we evaluated whether ADAM9 expression plays a critical role in metastatic processes in vivo and in angiogenesis. We first found that high ADAM9 expression was correlated with poor lung adenocarcinoma patient prognosis on Prognoscan data base. In vivo model based on intravenous injection in nude mice showed that a stable downregulation of ADAM9 in A549 (TrA549 A9-) cells was associated with a lower number of nodules in the lung, suggesting lower potentials for extravasation and metastasis. On a subcutaneous xenograft we showed that TrA549 A9− produced significantly smaller tumours and exhibited fewer neovessels. In addition, in vitro human umbilical vein endothelial cells exposed to supernatant from TrA549 A9− could reduce the formation of more vessel-like structures. To further understand the mechanism, a human antibody array analysis confirmed that five cytokines were downregulated in TrA549 A9− cells. Interleukin 8 was the most significantly downregulated, and its interaction with CXCR2 was implicated in angiogenesis on an in vitro model. These results emphasize the critical influence of ADAM9 on lung cancer progression and aggressiveness. ADAM9 should at least be a marker of cancer aggressiveness and a potential therapeutic target for cancer treatment.
Background
Cancer progression, metastasis and angiogenesis remain major concerns in cancer patients. These events are associated with decreased life expectancy. Research on the genes and molecular processes involved in oncogenesis is essential for the diagnosis and treatment of cancer and for the evaluation of the prognosis.
In many circumstances, metastasis and angiogenesis seem to be interdependent. New vessels are required for cell growth, and capillary and postcapillary venules develop because of endothelial sprouting or intussusceptive microvascular growth. 1 A balance between positive and negative molecules controls angiogenesis. This concept led to the notion of the ‘angiogenic switch’, which depends on the increased production of one or more positive angiogenesis regulators,2–4 including growth factors such as vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), metalloproteinase or interleukin 8 (IL-8), interacting with CXCR2.5–11 The activation and modification of growth factors, the regulation of chemokine activity and the release of membrane-anchored cytokines into the tumour cell microenvironment implicate transmembrane metalloproteases12,13 such as matrix metalloproteinases (MMPs) and A Disintegrin And Metalloproteases (ADAMs). Multiple functions of the ADAMs family contribute to angiogenic activity.14–16
Guaiquil et al. 17 demonstrated the role of ADAM9 in pathologic neovascularization using a model of retinopathy. In particular, they suggested that the metalloprotease activity of ADAM9 contributes to the cleavage and release of pro-angiogenic factors. Some studies have suggested the involvement of ADAM proteins, specifically ADAM9 protein, in the process of angiogenesis.14–17 However, the precise role of ADAM9 and the mechanism whereby ADAM9 mediates tumour angiogenesis are not fully understood.
ADAM9 is highly expressed in prostate, pancreas and breast cancers and is associated with cancer progression and a poor clinical outcome.15,18–23 Mazzocca et al. 23 showed that ADAM9 promotes liver cancer cell invasion by inducing stromal modulation in vitro. In another model of gastric cancer, ADAM9 promotes cancer cell proliferation and invasion, and its expression is upregulated by hypoxia. 24 In several aggressive lung adenocarcinoma cell lines (Bm7, H1299, A549), ADAM9 expression is upregulated and enhances migration of lung cancer cells.25–28 We reported that human lung cancer cell invasion and adhesion induced by oxidative stress is mediated by ADAM9 protein and its secreted form in vitro. 29
In this study, we sought to elucidate whether ADAM9 contributes to lung cancer progression and its mechanisms. Based on a cohort of patients with lung cancer, bioinformatics analysis of databases has revealed that there is a trend between (overall or free relapse) survival and ADAM9 gene expression. An in vivo metastatic model and angiogenesis experiments (in vitro and in vivo) were developed to assess the roles of the ADAM9 protein in metastatic and angiogenic processes. These observations provide strong evidence that ADAM9 plays a crucial role in the aggressiveness of adenoma lung cancer and that it might be an effective target for future therapies in metastatic non–small cell lung cancer (NSCLC).
Materials and methods
Materials
A549 (human lung carcinoma, ATCC CCL-185) cell lines were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA) and were cultured in Dulbecco’s modified Eagle’s medium (DMEM) Glutamax-I (Life Technologies, Cergy Pontoise, France) supplemented with 10% heat-inactivated foetal calf serum (Life Technologies) and antibiotics. Human umbilical vein endothelial cells (HUVECs) were cultured in DMEM supplemented with 10% heat-inactivated foetal calf serum (Life Technologies) and antibiotics. All incubations were performed at 37°C in a 5% CO2-enriched atmosphere. The monoclonal anti-human ADAM9 ectodomain antibody (MAB939) and anti-mouse IgG-HRP antibody (HAF018) were obtained from R&D Systems (Minneapolis, MN, USA). Monoclonal anti-β-actin, peroxidase conjugate (A3854) was obtained from Sigma (St. Louis, MO, USA), and anti-CXCR2 antibody (ab24963) was obtained from Abcam (Cambridge, UK). EC Matrix gel was purchased from Chemicon International (Temecula, CA, USA). Small interfering RNA (siRNA) lentiviral particles (ADAM9− and controls) and siRNA lentiviral particle support reagents were obtained from Santa Cruz Biotechnology (Dallas, TX, USA).
Stable transfection for the in vivo model
A stable line transfected with ADAM9 siRNA was created using lentivirus transduction. This line allows permanent extinction (for more than 60 days) of the ADAM9 protein, according to the manufacturer’s instructions. Briefly, cells were cultured in a 12-well plate to approximately 60%–70% confluency. The medium was then removed and replaced with transfection medium (1 mL), and cells were infected by adding siRNA lentivirus particles (60 μL). After 5 h, the transfection medium was replaced with DMEM containing 20% foetal bovine serum (FBS) and antibiotics. Next, stable clones expressing siRNA were selected with puromycin dihydrochloride. The cells were cultured with fresh puromycin–containing medium every 3–4 days. We confirmed the inhibition of ADAM9 expression in this line (TrA549 A9−) by western blotting. A control was established using a lentivirus carrying a random RNA (TrA549 ct).
To enhance ADAM9 expression in A549 cells, cells were exposed to phorbol 12-myristate 13-acetate (PMA; 50 ng/mL for 10 h), a potent inducer of protein kinase C (PKC), 30 known to enhance the metalloprotease activity of ADAMs. 31
Western blot analysis
Cells were scraped and harvested in radioimmunoprecipitation assay (RIPA) lysis buffer and then were sonicated three times (10 s) and centrifuged at 14,000 r/min for 10 min at 4°C. Total proteins (50 μg/well) were resolved using 7.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) under non-reducing conditions and then were transferred to nitrocellulose membranes and blocked with 5% non-fat dry milk in Tris-buffered saline with Tween 20 (TBST). The membranes were blotted with primary anti-ADAM9 monoclonal antibody (1 μg/mL) or actin (1/10,000) as the molecules of interest and were then washed. Bound antibody was detected using horseradish peroxidase (HRP)-conjugated secondary antibody (1/1000) followed by detection with the enhanced chemiluminescence (ECL) system (Pierce, Rockford, IL, USA).
In vitro angiogenesis assay
Supernatants from TrA549 control cells and TrA549 A9− cells exposed or not exposed for 18 h to PMA (50 ng/mL) were recovered. Extracellular (EC) matrix gel was used according to the manufacturer’s protocol. HUVECs (2 × 104 cells/well) were cultured in 96-well plates with the supernatant of each cell type. The HUVECs were then examined using calcein. Tubular structure formation within the matrix was assessed 22 h later using a fluorescence microscope. The total length of the new capillary structures was measured using ImageJ software (National Institutes of Health (NIH), Bethesda, Maryland, USA). Overall, five pictures were taken per well, and the experiments were performed three times.
Enzyme-linked immunosorbent assay or IL-8 immunoquantification
Next, 2.5 × 104 cells of each type (TrA549 ct, TrA549 A9− and A549 cells) were plated onto 96-well plates and were exposed or not exposed to PMA (50 ng/mL) for 10 h. Equal volumes of the cell culture supernatants were collected.
IL-8 protein quantification was assessed using the Quantikine IL-8 ELISA kit (R&D Systems). Absorbance was measured at 450 nm according to the manufacturer’s instructions. The results were expressed by comparing the absorbances (at 450 nm) of the different cell types to that of A549 cells. Cell quantification was performed by cellular crystal violet staining.
In vivo studies
The experimental plan was validated by the local animal experiment board. The study was approved by the Regional Ethics Committee on Animal Experimentation (protocol CEEA34.CN.023.11) and was performed according to a protocol approved by the Local Committee of Laboratory Investigation and Animal Care of our institution and met the requirements of the European Convention for the Protection of Vertebrates Used for Scientific Purposes.
Tumour invasion
Female nude-Foxn1nu/Fox1+ mice, aged 6 weeks, were acquired from Harlan, France. The first step consisted of subcutaneously injecting 1 × 107 tumour cells into the flanks of one mouse to induce a cancer xenograft for each group (A549, TrA549 A9− and TrA549 ct). On day 14, the mice were sacrificed, and the tumours were collected. The cells were then plated for in vitro culture (1 × 106 cells in 10 mL of DMEM supplemented with 10% FBS for 7 days). Next, the cells were harvested, washed once in serum-free medium, resuspended in phosphate-buffered saline (PBS) and centrifuged. Trypan blue staining was used to assess cell viability.
For the second step, 5 × 106 tumour cells, grown from the xenograft of A549 cells, TrA549 ct cells and TrA549 A9− cells, were injected intravenously into a caudal vein (six mice in each group). On day 45, the mice were sacrificed. The lungs and the brain were removed and embedded in paraffin and were then sliced in the middle and at the bottoms of all blocks. The slices were stained with haematoxylin and eosin for routine histological examination.
The percentage of pulmonary invasion was calculated as the ratio between the total sizes of the metastases within the total size of the lung using ImageJ software. For this analysis, three tumour sections were cut, one at the top, one in the middle and one at the back. Overall, five representative pictures were taken per available field.
Tumour growth
Female nude-Foxn1nu/Fox1+ mice, aged 6 weeks, were acquired from Harlan. Next, 1 × 107 tumour cells (TrA549 A9−, TrA549 ct and A549 cells) were injected subcutaneously into the right flanks of mice (three mice per group) to generate a subcutaneous tumour. Tumour growth was measured every 3 days, and the mice were euthanized on day 32. The size of the tumour mass was measured using a digital calliper, and the tumour volume was calculated as 1/2 × length × width2 in mm3 for the A549 xenograft.
Immunohistochemistry and quantification of microvascular density
Paraffin-embedded fragments of the tumour species were sectioned (5-μm thicknesses). Sections were pre-incubated with 2% bovine serum albumin in PBS for 1 h, and then, the primary anti-CD31 antibody was added and incubated for 45 min (1:50; Abcam). The sections were incubated with HRP-conjugated anti-rabbit secondary antibody (1:250; Dako, Glostrup, Denmark). After development with diaminobenzidine solution (Vector Laboratories Burlingame, CA, USA), the sections were counterstained with haematoxylin. Sections were evaluated under a microscope (Olympus BX60; Olympus, Tokyo, Japan) at 20× magnification and analysed with ImageJ software (NIH). Blood vessel density was quantified by counting the total number of CD31-positive vessels across the entire section of the tumour. For this analysis, three tumour sections were cut, one at the top, one in the middle and one at the back. Overall, five representative pictures were taken per available field.
Bioinformatics
The associations between the gene expression levels and NSCLC patient prognosis were obtained from the PrognoScan database (http://www.prognoscan.org/). PrognoScan is a large collection of publicly available cancer microarray datasets with clinical annotations and a tool for assessing the biological relationships between gene expression and prognosis. In the PrognoScan database, the association of gene expression with the survival of patients was evaluated using the minimum p value approach and the corrected p value. Briefly, patients were first arranged by expression levels of a given gene. They were then divided into high- and low-expression groups at all possible cutoff points, and the risk differences between any two groups were estimated by the log-rank test. Finally, the cutoff point resulting in the most pronounced p value was selected.
Statistical analysis
Statistical analysis was performed using the means of data obtained from at least three independent experiments. All results are presented as the mean ± standard error of the mean (SEM). Student’s t test was used to compare significant differences in the experimental values between the two samples. When the analysis included more than three groups, one-way analysis of variance was used (GraphPad Prism; GraphPad Software, San Diego, CA, USA). A p value of 0.05 was considered to be statistically significant.
Results
First, we investigated the correlation between the level of ADAM9 protein expression and overall survival of patients with lung adenocarcinoma using ‘Gene expression data for pathological stage I–II lung adenocarcinomas’, obtained from the PrognoScan database (http://www.prognoscan.org/). Data without a corrected p value were not analysed. Two datasets revealed a correlation between a very high ADAM9 expression and overall survival (corrected p < 0.05; Table 1, Figure 1). In the six other datasets, there is a very high trend for an influence of high ADAM9 protein expression and a poor overall survival. In the last study, there is a negative association but not significant (corrected p = 0.286). For the correlation of ADAM9 expression regarding relapse-free survival, two datasets were listed and showed that high expression of ADAM9 protein was correlated with a poor relapse-free survival (corrected p < 0.01; Table 1, Figure 1). Analysis of data from another database (The Cancer Genome Atlas (TCGA) database) supplied similar results (data not shown).
Correlation between ADAM9 gene expression and the survival (overall and relapse-free) in lung adenocarcinoma cancer patients. Data were obtained from PrognoScan database.
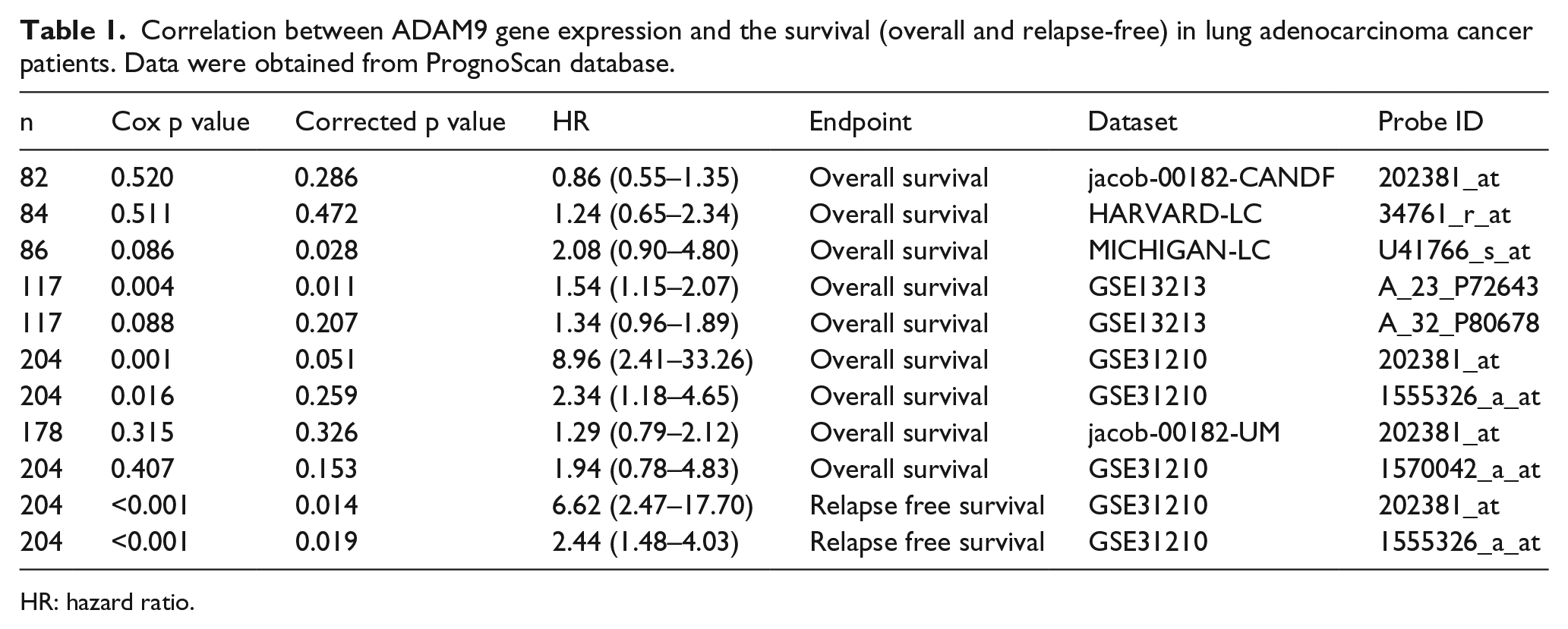
HR: hazard ratio.
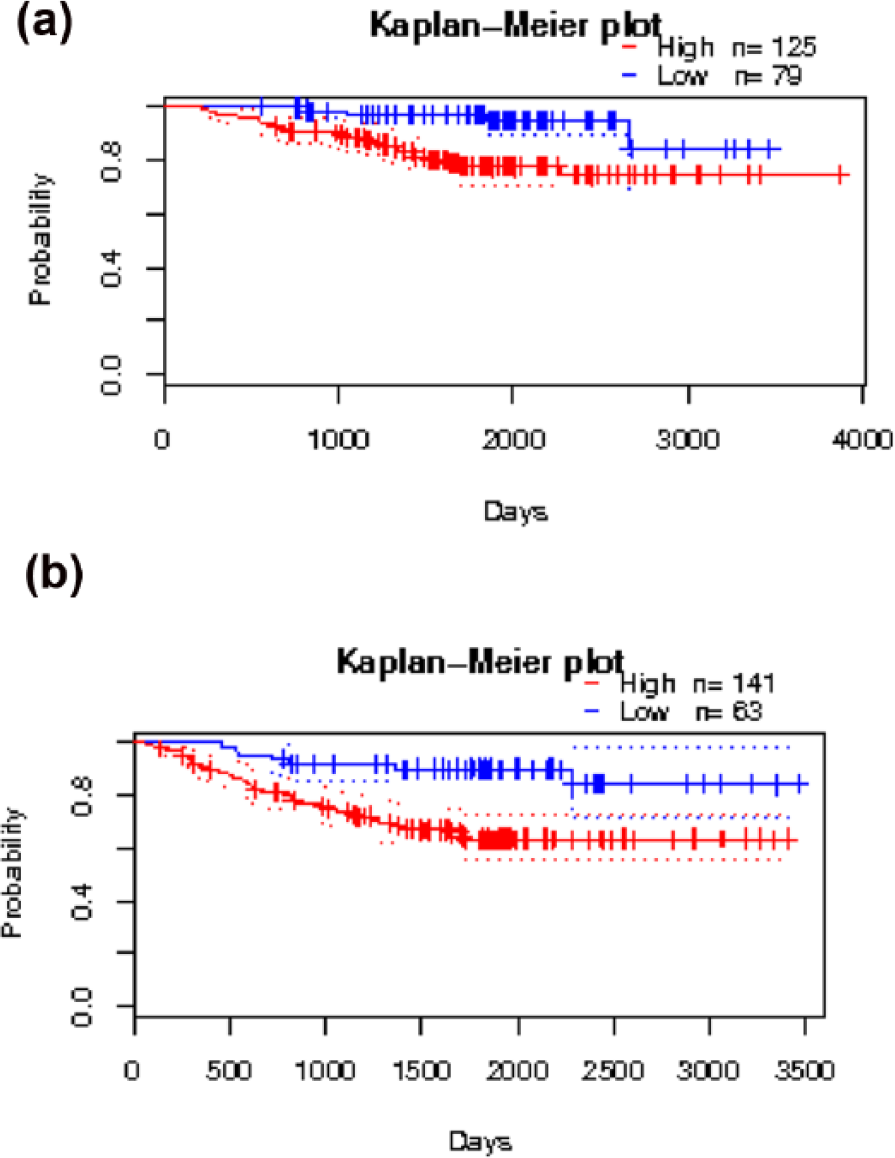
High ADAM9 expression in lung adenocarcinoma is associated with poor cancer prognosis. (a) Correlation between the overall survival of patients with lung adenocarcinoma and ADAM9 expression. The Kaplan–Meier graph was obtained from a dataset of the PrognoScan database. (b) Correlation between the relapse-free survival of patients with lung adenocarcinoma and ADAM9 expression. The Kaplan–Meier graph was obtained from a dataset of the PrognoScan database.
To explore the influence of ADAM9 expression in aggressive adenocarcinoma lung cancer, a model of stable downregulation of ADAM9 expression in A549 cells was developed. ADAM9 protein expression was stably downregulated in A549 lung cancer cells with a lentiviral vector carrying ADAM9-specific siRNA (TrA549 A9−). A control sample was produced in A549 cells with a lentiviral control vector (TrA549 ct). Both cell lines were used in subsequent experiments.
As shown in Figure 2(a), high knockdown efficiency was achieved in A549 cells. After 10 h of exposure to PMA, known to enhance the metalloprotease activities of ADAMs, ADAM9 expression was increased in both native A549 cells and A549 cells transfected with the control vector; however, the expression remained low in A549 cells transfected with vectors carrying ADAM9-specific siRNA.
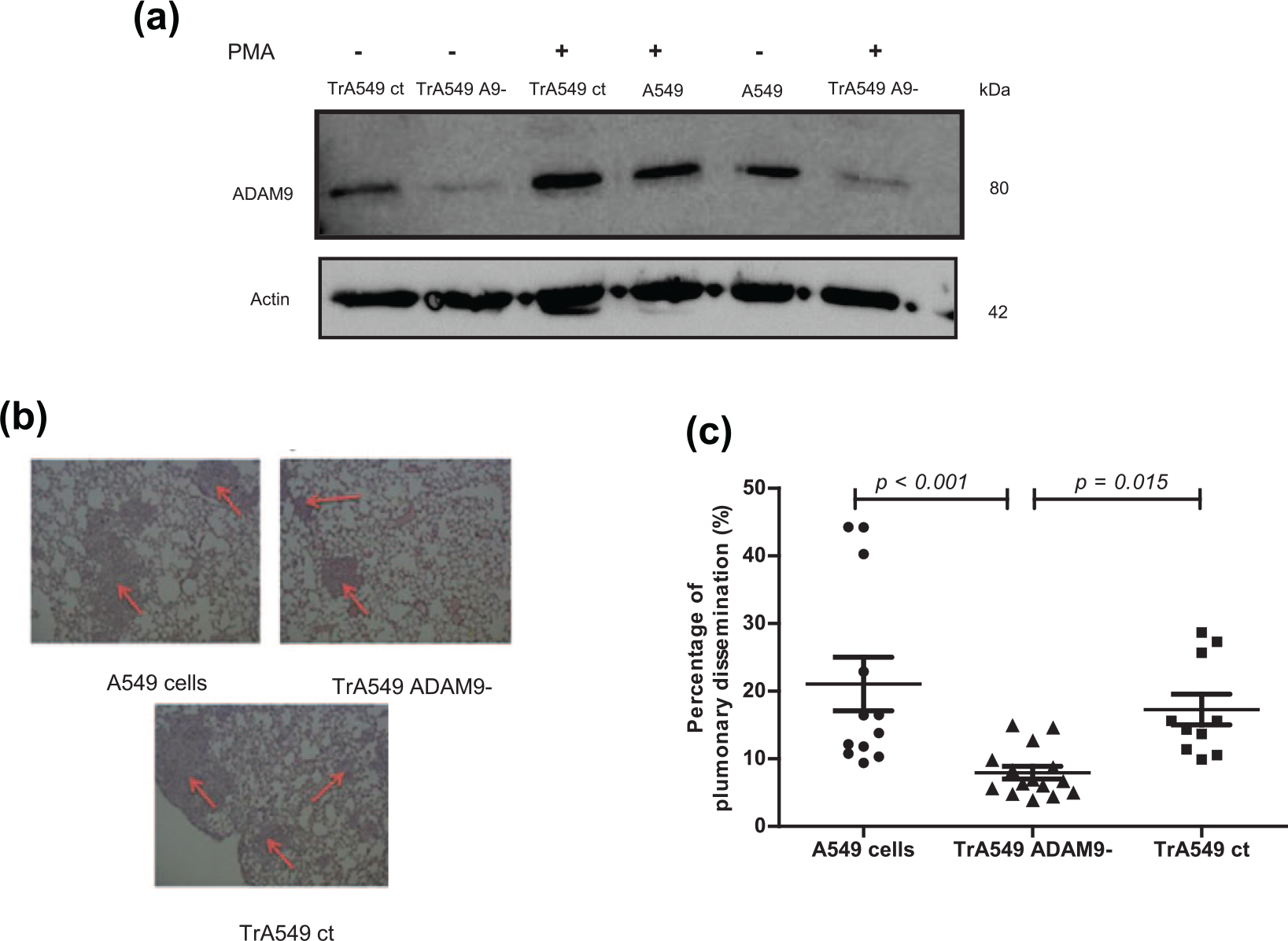
Role of ADAM9 expression in lung metastatic dissemination using an in vivo model. A549 cells were transfected with siRNA against ADAM9− (TrA549 A9−) or with siRNA control (TrA549 ct). For the in vivo model, stable cell lines were first established after lentiviral transduction and were then xenografted (subcutaneous injection of 107 cells into mice for 14 days) followed by intravenous injection into nude mice (106 cells diluted in medium). Three groups of cells were injected: A549, TrA549 ct and TrA549 ADAM9− cells. Nude mice were sacrificed on day 32. (a) Transmembrane ADAM9 protein expression was detected by western blotting. After A549 transfection, the cells were exposed or not exposed to PMA (50 ng/mL) for 10 h. (b) Images of pulmonary metastasis in nude mice. The tissues were stained with haematoxylin and eosin at a magnification of 20×. (c) Evaluation of the pulmonary invasion percentage in nude mice. This percentage was calculated as the ratio between the total size of the metastases and the total size of the lung using ImageJ software (mean ± SD).
Inhibited ADAM9 expression reduces lung metastasis formation in a mouse model
The murine model used is based on the intravenous injection of A549 cells that had previously been subcutaneously xenografted. The first step of subcutaneous xenografting exerts a characteristic selection pressure in favour of increasingly malignant tumour phenotypes. All nude mice developed lung metastases (Figure 2(b)). Among 18 mice, only one mouse developed a brain tumour in the A549 cell group compared with the TrA549 ADAM9− group and TrA49 control group (no significant). At day 32, in mice implanted intravenously with A549 cells, cancer cells represented 18.9% of the lung, whereas the group xenografted with TrA549 A9− tumour cells represented only 8.36% of the lung (Figure 2(c)). We revealed that the percentage of pulmonary dissemination in mice injected with TrA549 ADAM9− cells is decreased compared to mice injected with A549 cells (p < 0.001) and mice injected with TrA549 control cells (p = 0.015).
Inhibition of ADAM9 expression in lung cancer cells resulted in a significant (p < 0.01) decrease (77%) in the pulmonary metastatic process of A549 lung cancer cells.
ADAM9 downregulation decreases in vivo A549 tumour growth and tumour neovascularization
To confirm the effects of ADAM9 protein expression on growth and tumour neovascularization, we established an in vivo model based on the development of subcutaneous tumours generated by injection of stable cell lines after lentiviral transduction. From day 25, we observed that tumours derived from the parental cell line grew much faster than those from the TrA549 A9− cell line (Figure 3(a)).
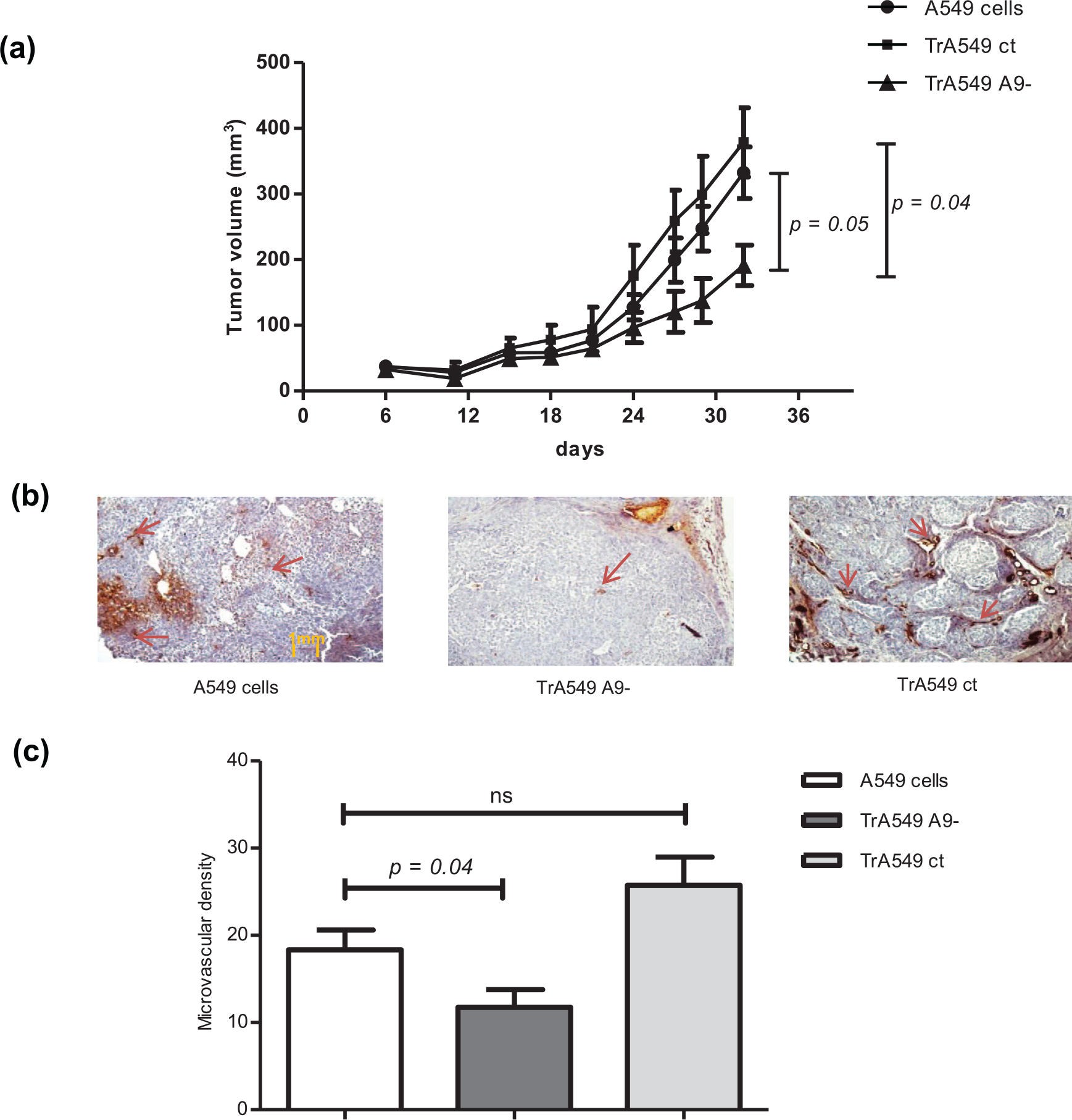
Effect of ADAM9 expression on the growth and angiogenesis of human lung cancer cells in nude mice. A549 cells were transfected with siRNA against ADAM9 protein (TrA549 ADAM9−) or with siRNA control (TrA549 ct). Nude mice were injected subcutaneously with 107 cells diluted in medium. Three groups of cells were injected: A549, TrA549 ct and TrA549 ADAM9− cells. (a) Mice with subcutaneous injection of A549 lines were monitored for tumour growth twice weekly. Values are the mean ± SD of tumour volume. (b) Tumours were excised after 32 days, fixed in formalin and embedded in paraffin. The formation of capillary-like structures was assessed in sections by CD31 staining. For this analysis, three tumour sections were cut, one at the top, one in the middle and one at the back. Overall, Five representative pictures were taken per available field to quantify microvascular density with mean ± SD (ns = no significance).
To assess the role of ADAM9 in tumour neovascularization, immunohistochemical staining for the endothelial cell marker CD31 was performed on excised tumours. Staining revealed a statistically significant decrease (36%) in the number of CD31-positive vessels in tumours derived from TrA549 A9− cells compared with tumours derived from the parental cell line (p = 0.04; Figure 3(b)). These findings indicate decreased tumour growth and angiogenesis in the absence of ADAM9 protein expression.
ADAM9 expression in A549 cells increases the formation of vessel-like structures by HUVECs in vitro
To address whether ADAM9 protein has an impact on angiogenesis in vitro, endothelial vascular cells were exposed to the supernatants of A549 cells stably transfected or not exposed to ADAM9 siRNA. Each cell line (A549 cells, TrA549 ct and TrA549 A9−) was exposed to PMA for 10 h to induce ADAM9 overexpression in native cells. As shown in Figure 4(a), endothelial vascular cells exposed to the supernatant of the TrA549 A9− cell line appeared more rounded and retracted than HUVECs exposed to the supernatant of the parental cell line. A549 cells are supposed to release factors that can induce angiogenesis. Indeed, PMA-pre-treated A549 cells increased the total length of new vessel formation by HUVECs on Matrigel by 2.86- and 2.93-fold (A549 cells and TrA549 ct; p < 0.001, p = 0.009, respectively) compared with untreated A549 cells. In contrast, no increase was observed in the branching length of the capillary-like structures when ADAM9-knockdown A549 cells were pre-treated with PMA (p = 0.19; Figure 4(b)). Taken together, these results clearly indicate that ADAM9 protein is a crucial mediator of the pro-angiogenic stimulus, emanating from tumour cells.

Role of the ADAM9 protein in A549 cancer cells in the formation of capillary-like structures in HUVECs. A549 cells were transfected with siRNA against the ADAM9 protein (TrA549 A9−) or with siRNA control (TrA549 ct). (a) Image of HUVECs on Matrigel cultured with different supernatants: A549 cells exposed or not exposed to PMA or TrA549 A9− exposed or not exposed to PMA. (b) Supernatants of A549-transfected cells or not were cultured with HUVECs and were plated on Matrigel for 22 h. Next, HUVECs were examined with calcein, and the numbers of branches in the capillary-like structures were calculated. Overall, five representative fields at a magnification of 40× were quantified in each experiment (mean ± SD of three independent experiments; ns = no significance).
ADAM9 protein expression induces IL-8 secretion in A549 cells and enhances the capillary-like structures formed by HUVECs in vitro
To further define the mechanism whereby ADAM9 protein promotes angiogenesis, different conditioned supernatants from A549 cells (TrA549 A9− and TrA549 ct cells exposed or not exposed to PMA) were analysed using a human antibody array (Figure 5(a)) capable of detecting the expression of 43 angiogenesis-related proteins. As shown in Figure 5(a), five angiogenesis mediators were modulated by ADAM9 expression in A549 cells: IL-8, leptin, ENA78, uPAR and ITAC, with the greatest effect observed for IL-8.
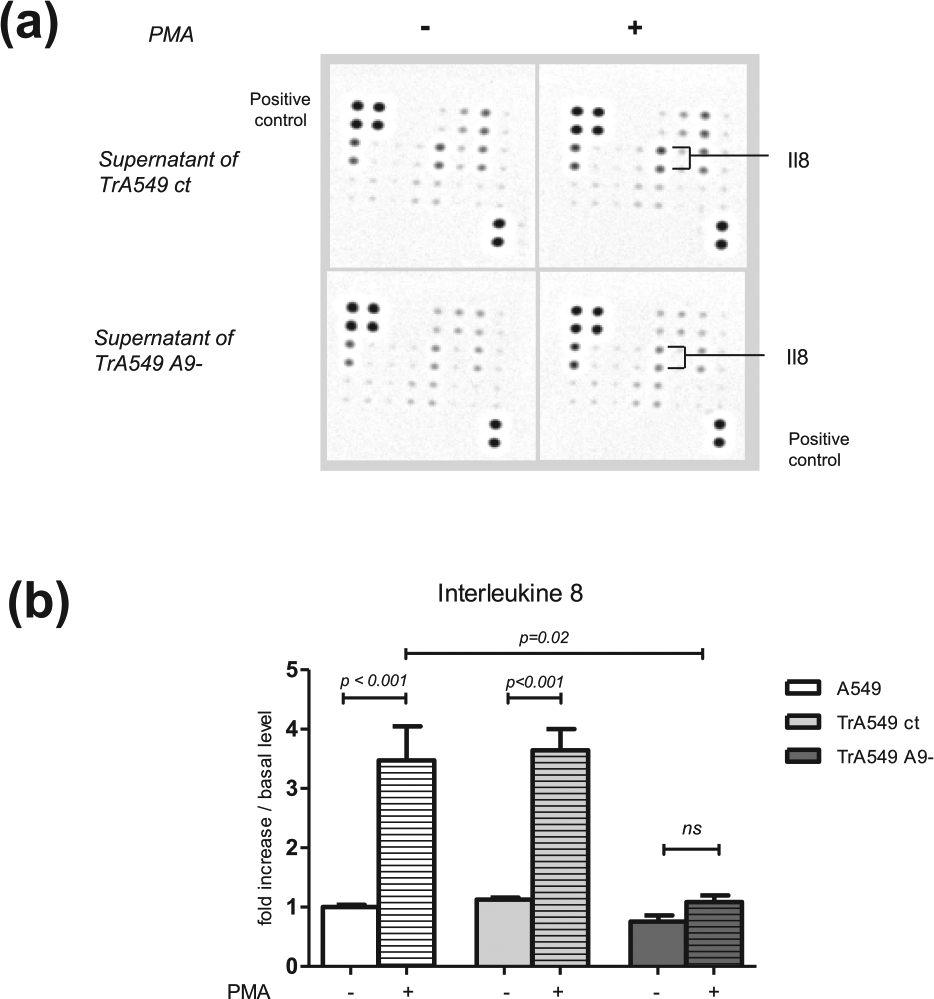
Detection and quantification of IL-8 expression modulated by ADAM9 expression in lung cancer cells. A549 cells were transfected with siRNA against ADAM9− (TrA549 A9−) or siRNA control (TrA549 ct) and incubated or not with 50 ng/mL PMA for 10 h. (a) Human antibody array analysis was used to determine the release of 43 human angiogenic factors from the conditioned medium of A549 cells transfected with siRNA ADAM9− or control and treated or not with PMA. (b) ELISA to measure IL-8 production from ADAM9(−)-transfected, control-transfected and parental A549 cancer cells. The results were normalised to the cell numbers. The data represent the mean ± SD of three independent experiments performed in triplicate.
Using enzyme-linked immunosorbent assay (ELISA), IL-8 secretion was determined to be 3.82-fold enhanced in wild-type (WT) A549 cells exposed to PMA compared with that in untreated cells (p < 0.01; Figure 5(a)). While this induction was highly significant for A549 cells stably transfected with the control vector (3.54-fold; p < 0.01), the level of IL-8 secretion was not significantly increased in TrA549 A9− cells exposed to PMA. Moreover IL-8 secretion is decreased significantly (p = 0.02) when A549 cells do not express ADAM9 protein. Taken altogether, these results strongly support the hypothesis that ADAM9 protein expression promotes IL-8 secretion in A549 cells.
IL-8 enhances angiogenesis in vitro through its interaction with CXCR2
IL-8 is known to bind to and activate the high-affinity receptor CXCR2. The specific implication of this receptor in angiogenesis was assessed by adding a blocking anti-CXCR2 antibody to the supernatant to measure tube formation, whereas the supernatant from WT A549 cells stimulated with PMA induced 171% increase in the total length of new vessel formation by HUVECs (p < 0.0001; Figure 6), the same experiment performed in the presence of blocking anti-CXCR2 antibody resulted in complete inhibition of this induction (p = 0.0001). When the same experiment was performed on TrA549 A9− cells, the anti-CXCR2 antibody did not modulate new vessel formation. These results consider previous findings supporting the hypothesis whereby ADAM9 leads to the synthesis of IL-8, which interacts with CXCR2 to stimulate angiogenesis.
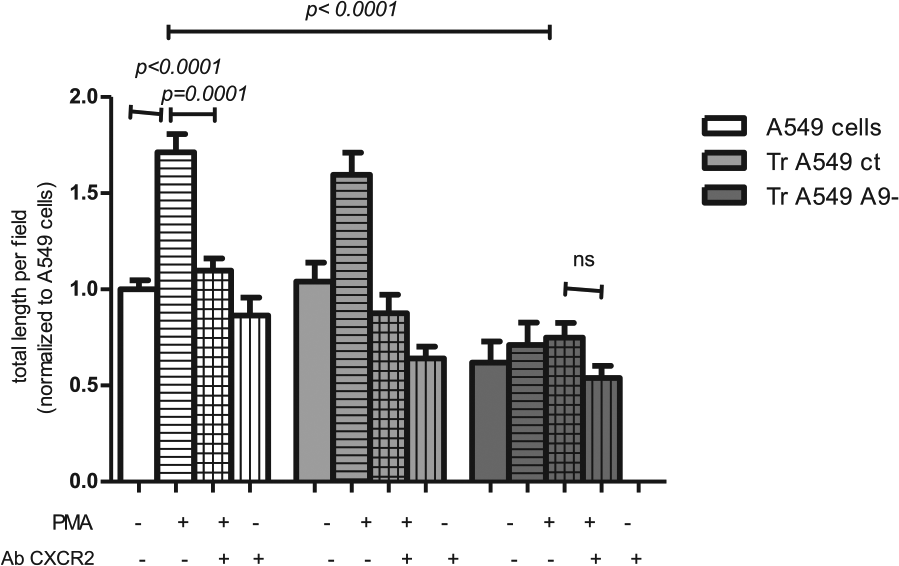
Interleukin 8 expression induced by ADAM9 expression in lung cancer cells modulates angiogenesis in vitro through its receptor, CXCR2. A549 cells were transfected with vectors encoding siRNA against ADAM9 (TrA549 A9−) or control siRNA (TrA549 ct). Next, cells were stimulated or not by PMA (50 ng/mL for 10 h). The supernatants of different lines were cultured with HUVECs and were plated on Matrigel for 22 h. Blocking CXCR2 antibody (5 µg/mL) was added to the culture medium. Next, tube formation was assessed with calcein, and the numbers of branches of the capillary-like structures were evaluated. Three representative fields at a magnification of 20× were quantified in each experiment (mean ± SD of three independent experiments; ns = no significance).
Discussion
In a previous study, we showed in vitro that ADAM9 expression in lung adenocarcinoma cells correlates with an aggressive phenotype. 29 Here, using A549 cells stably transfected with anti-ADAM9 siRNA, we report for the first time that metastatic dissemination and angiogenesis are dependent on ADAM9 protein expression and its interactions with IL-8/CXCR2.
The intravenous xenograft murine model showed that knockdown of ADAM9 protein expression in lung cancer cells decreased the capacity of cells for metastasis formation. Lin et al. also found that the in vivo inhibition of ADAM9 protein expression on lung cancer cells decreased brain metastasis by enhancing the tissue plasminogen activator. 26 Mammadova-Bach et al. 32 showed that the interplay between platelet α6β1 and tumour cell-expressed ADAM9 promotes efficient lung metastasis. ADAM9 overexpression portends a poor prognosis for patients with non–small cell lung cancer. Our study demonstrated that ADAM9 expression in lung cancer cells plays an important role in their capacity to disseminate.
The subcutaneous xenograft murine model showed that ADAM9 protein expression knockdown in lung cancer cells strongly decreased tumour growth in mice compared with controls, demonstrating that ADAM9 protein also plays a role in tumour proliferation. This activity on tumour growth certainly reflects the aggressive phenotype of the cells, but our work also showed that mice xenografted with WT A549 expressing ADAM9 developed more intense tumour angiogenesis than mice transfected with TrA549 ADAM9− cells that could not express ADAM9. Another model of pathologic neovascularization in retinopathy described by Guaiquil et al. demonstrated the importance of ADAM9 expression in angiogenesis. They showed that ADAM9 expression affects pathologic neovascularization by increasing the shedding of EphB4, Tie 2, FLK-1, CD40, VCAM and VE cadherin along with other membrane proteins from endothelial cells. 17 Roy et al. 33 also showed that the ADAM9 protein can stimulate capillary formation by vascular endothelial cells, confirming that ADAM proteins play a very important role in angiogenesis. These metalloproteinases participate in the degradation and remodelling of extracellular matrix and vascular cells through their ectodomain. Although these phenomena are important, ADAM9 could have a direct influence on vascular endothelial cells, allowing the formation of new vessels.
To explain its mechanism in endothelial cells, we observed that five pro-angiogenic factors were upregulated when ADAM9 was expressed. Among these five factors, the most significant was IL-8, a chemokine that acts on endothelial cells to promote migration, invasion and in vivo angiogenesis in several cancers.4,5,34,35 Our results revealed that ADAM9 expression in lung cancer cells promotes IL-8 release. Moreover, the use of anti-CXCR2 antibodies blocked the formation of neovessels induced by ADAM9 expression in vitro. Keane et al. showed that the depletion of the CXCR2 receptor inhibits tumour growth and angiogenesis in a mouse model of lung cancer, demonstrating the important role of CXCR2 in angiogenesis.34,35 Our results suggest that IL-8 expression induced by ADAM9 expression in lung cancer cells modulates angiogenesis in vitro through its receptor CXCR2. These findings strongly highlight that ADAM9 plays an important role in carcinogenesis and the metastatic process. In particular, it may be involved in VEGF-independent angiogenesis, which might be responsible for resistance to anti-angiogenic therapies.
Guaiquil et al. 17 demonstrated a reduction in retinal vasculature development in a model of retinopathy in ADAM9−/− mice. We have demonstrated that the ADAM9 protein has an impact on tumour neovascularization through angiogenesis using both in vitro and in vivo models and may indirectly induce stunted tumour growth and proliferation. That study, combined with our results, confirms the involvement of ADAM9 protein in tumour neovascularization. To summarise our previous study and our results, ADAM9 expression and its effects on tumour metastasis can be envisaged as depicted in Figure 7. 29

ADAM9 expression in lung cancer cells and its potential effects on tumour metastatic process.
Conclusion
We have demonstrated that ADAM9 isoforms in lung cancer cells can induce metastatic processes such as angiogenesis in a paracrine manner. Indeed, the ADAM9 protein allows the release of IL-8 (a pro-angiogenic factor) in cancer cells. Its interaction with its receptor, CXCR2, leads to the modulation of neovascularization.
Footnotes
Acknowledgements
The authors thank Lorna Saint Ange for editing. C.M.K. and M.A. performed all experiments and drafted the manuscript. A.T.-S. performed the in vivo experiments and helped in drafting the manuscript. C.C. participated actively in cell cultures and technical aspects in vitro. C.N.-O. participated in the in vivo models and technical aspects and in the design of the in vivo models. F.B. participated in the design of the study and coordination and helped drafting the manuscript. J.A. conceived the study, participated in the design of the study and helped drafting the manuscript. F.L. conceived the study, established the design of the study, participated in the coordination and drafted the manuscript. All authors read and approved the final manuscript. C.M.K. and M.A. are co-authors.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
The study was approved by the Regional Ethics Committee on Animal Experimentation (protocol CEEA34.CN.023.11). The study was performed according to a protocol approved by the Local Committee of Laboratory Investigation and Animal Care of our institution and met the requirements of the European Convention for the Protection of Vertebrates Used for Scientific Purposes.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partly supported by both ‘Société Française de Pharmacie Oncologique’ and the French National Academy of Pharmacy.