
Abstract
Emerging evidence suggests that microRNAs are critical regulators of cancer development and progression. MicroRNA-195 has been reported as a cancer-related microRNA in many human cancers. However, the role of microRNA-195 in pancreatic cancer remains largely unknown. Here, we show that microRNA-195 is downregulated in pancreatic cancer tissues and cell line. Also, we show that overexpression of microRNA-195 inhibits the proliferation and invasion of pancreatic cancer cells, whereas suppression of microRNA-195 promotes proliferation and invasion. We show that microRNA-195 directly targets the fatty acid synthase enzyme and negatively regulates the expression of fatty acid synthase. Also, we show that fatty acid synthase expression is inversely correlated with microRNA-195 expression in pancreatic cancer tissues. Moreover, our results show that microRNA-195 inhibits Wnt signaling in pancreatic cancer cells. By restoring fatty acid synthase expression, we were able to reverse the antitumor effects of microRNA-195 in pancreatic cancer cells. Taken together, our findings show that microRNA-195 inhibits pancreatic cancer cell proliferation and invasion by regulating the fatty acid synthase/Wnt signaling pathway, suggesting a tumor suppressive role for microRNA-195 in the development and progression of pancreatic cancer. Thus, inhibiting fatty acid synthase by microRNA-195 may serve as a novel therapeutic approach for the treatment of pancreatic cancer.
Introduction
Pancreatic cancer remains one of the deadliest malignancies, being the fourth leading cause of cancer-related death. 1 Despite advances in cancer treatment over recent decades, pancreatic cancer patients continue to have an extremely low survival rate.2,3 Pancreatic cancer is often diagnosed at an advanced stage with extensive local tumor metastasis and is unresponsive to many treatments. 4 Thus, there is an urgent need to develop novel diagnostic and treatment approaches for pancreatic cancer.
In recent years, microRNAs (miRNAs), a group of small and noncoding RNAs, have emerged as critical regulators for gene expression. 5 miRNAs negatively modulate gene expression by targeting the 3′-untranslated region (3′-UTR) of target messenger RNAs (mRNAs). 6 Thus, miRNAs are involved in the regulation of various physiological and pathological processes. 7 Various miRNAs are dysregulated during the development and progression of cancers, functioning as oncogenes and tumor suppressors. 8 These miRNAs can serve as potential biomarkers for diagnosis and prognosis, as well as therapeutic molecular targets. 9 An increasing number of studies show that various miRNAs are involved in the progression of pancreatic cancer.10,11 Targeting specific miRNAs such as miR-506 and miR-122 shows promising inhibitory effects on pancreatic cancer progression.12,13 Thus, miRNA-based therapy for pancreatic cancer may have potential future applications.
Recent studies have suggested that lipid metabolism (e.g. aerobic glycolysis) plays an important role in maintaining the malignancy of cancer cells by regulating membrane synthesis, signaling functions, and energy storage.14,15 Fatty acid synthase (FASN) is a pivotal enzyme in lipid metabolism, 16 catalyzing the biosynthesis of palmitate in a nicotinamide adenine dinucleotide phosphate reduced–dependent reaction. 17 High FASN expression has been found in many human cancers, including breast, ovarian, colorectal, and prostate cancer.18–21 High levels of FASN are correlated with increasing tumor burden and poor prognosis. FASN has been suggested as a critical regulator of various oncogenic signaling pathways, including protein kinase C, human epidermal growth factor receptor-2, and the Akt and Wnt signaling pathways. 16 Inhibition of FASN induces apoptosis and suppresses proliferation, invasion, and metastasis of cancer cells.22,23 Recent evidence has shown that FASN is overexpressed in serum and cancer tissues of pancreatic cancer patients.24–26 Suppression of FASN is associated with the prevention of pancreatic cancer development.27,28 Therefore, FASN serves as a potential prognostic marker and candidate therapeutic target for pancreatic cancer.
A growing body of evidence has suggested microRNA-195 (miR-195) as a tumor suppressor in many human cancers, including breast, prostate, and colorectal cancer.29–31 In a recent study, Rachagani et al. 32 report that miR-195 expression is decreased in a mouse model of pancreatic cancer. However, little is known about the biological function of miR-195 in the pathogenesis of pancreatic cancer. Here, we aimed to investigate the functional significance of miR-195 in pancreatic cancer. We found that miR-195 expression is downregulated in pancreatic cancer tissues and cell lines and that overexpression of miR-195 significantly represses the proliferation and invasion of pancreatic cancer cells. We identify FASN as a downstream target of miR-195 and show that miR-195 negatively regulates FASN expression in pancreatic cancer cells. Also, we show that miR-195 expression is inversely correlated with FASN expression. Our study demonstrates that miR-195 inhibits the proliferation and invasion of pancreatic cancer cells by targeting FASN, indicating a tumor suppressive role of miR-195 in pancreatic cancer. Thus, miR-195 can be used a novel inhibitor for FASN and, therefore, is a potential novel therapeutic target for pancreatic cancer.
Materials and methods
Ethics statement
This study was approved by the Institutional Human Experiment and Ethics Committee of The First Affiliated Hospital of Xi’an Jiaotong University and performed in accordance with the World Medical Association Declaration of Helsinki regarding the use of human clinical tissues. Every participant agreed to tissue donation and provided written informed consent.
Tissue specimens
A total of 15 pairs of pancreatic cancer tissue samples and matched adjacent normal pancreatic tissue samples were obtained from The First Affiliated Hospital of Xi’an Jiaotong University. All patients were histologically diagnosed as pancreatic ductal adenocarcinoma. The resected specimens were immediately frozen in liquid nitrogen and stored at −80°C for RNA extraction, and human invasive pancreatic ductal adenocarcinoma samples were histologically confirmed.
Cell culture
The human pancreatic cancer cell lines (PANC-1, AsPC-1, BxPC-3, and SW1990) and human normal pancreatic ductal epithelial cells, HPNE, were purchased from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). PANC-1 cells were grown in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Rockville, MD, USA); AsPC-1, BxPC-3, and SW1990 cells were grown in Roswell Park Memorial Institute (RPMI)-1640 medium (Gibco) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin mix (Sigma, St. Louis, MO, USA). HPNE cells were grown in DMEM containing 5% FBS, 10 ng/mL human recombinant epidermal growth factor, 5.5 mM
Real-time quantitative polymerase chain reaction
Total RNA was extracted using RNAiso Reagent (TaKaRa, Dalian, China) according to the standard protocols. For miR-195 detection, complementary DNA (cDNA) was synthesized using miScript Reverse Transcription Kit (QIAGEN, Dusseldorf, Germany). For mRNA expression analysis, cDNA was synthesized using M-MLV Reverse Transcriptase (TaKaRa). Real-time quantitative polymerase chain reaction (RT-qPCR) was performed using SYBR Premix Ex Taq TM II Kit (TaKaRa) on an ABI7500 real-time PCR system (Applied Biosystems, Foster City, CA, USA). Small nuclear RNA U6 served as the internal control for miR-195. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) served as the internal control for mRNA expression. The 2−ΔΔCt method was applied to calculate the relative gene expression.
Cell transfection
miR-195 mimics, inhibitor, and negative control (NC) were synthesized by RiboBio (Guangzhou, China). FASN expression vector was constructed by sub-cloning the open reading frame of FASN into the pcDNA3.1 vector (Sangon Biotech, Shanghai, China). The empty pcDNA3.1 vector served as a NC. Cell transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) in line with the manufacturer’s instructions. Cells were harvested after 48 h of transfection for subsequent experiments.
Cell Counting Kit -8 assay
Cells were plated into 96-well plates at a density of 1 × 104 cells/well and cultured overnight. After transfection with miR-195 mimics or inhibitor for 48 h, 10 µL of Cell Counting Kit -8 (CCK-8) solution (Sigma) was added to each well and subsequently incubated for another 1.5 h at 37°C. The optical density (OD) was determined at 490 nm using a microplate analyzer (BioTek Instruments, Inc., Winooski, VT, USA).
Colony formation assay
Cells (transfection for 48 h) were seeded into six-well plates at 1 × 103 cells/well. The plates were coated with 0.6% agarose in DMEM containing 10% FBS. Cells were grown for 14 days and then used for detection. Cell colonies were fixed and stained with 0.1% crystal violet (Sigma). Colonies were observed and counted under a microscope.
Cell cycle assay
Cells were serum-starved for 24 h and then transfected with miR-195 mimics or inhibitor for 48 h. Cells were harvested, digested with trypsin, washed with ice-cold phosphate buffer saline, and fixed with 70% ethanol. Then, cells were stained with 100 µg/mL of propidium iodide (Sigma) in the presence with 10 µg/mL of RNase A for 30 min in the dark. Cells were detected using a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA), and data were analyzed by TModFit 3.3 software (BD Bioscience).
Transwell invasion assay
Cells (transfection for 48 h) were re-suspended in 200 µL of serum-free medium and added to the upper chamber of Transwell filters precoated with Matrigel (BD Biosciences). Meanwhile, 500 µL of medium containing 10% FBS in the lower chamber was used as chemoattractant. After culture for 24 h, cells on the lower surface were harvested, fixed with 4% formaldehyde, and stained with 0.1% crystal violet (Sigma). Cell numbers were counted under a microscope.
Western blot analysis
Protein extraction was performed using radioimmunoprecipitation assay (RIPA) buffer (Beyotime Biotechnology, Haimen, China). Equal amounts of protein (50 µg) from each sample were separated by electrophoresis on 10% sodium dodecyl sulfate polyacrylamide gels and then transferred onto a polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA). After blocking with 5% non-fat milk, the membrane was hybridized with primary antibodies including anti-FASN and anti-GAPDH (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C overnight followed by incubation with horseradish peroxidase–conjugated secondary antibody (1:2000; Santa Cruz Biotechnology) for 1 h at room temperature. GAPDH served as the loading control. The immuno-signals were visualized using an enhanced chemiluminescence system (Millipore). Band intensity was quantified with Image-Pro Plus 6.0 software (Media Cybernetics, Inc., Rockville, MD, USA).
Dual-luciferase reporter assay
The wild-type or mutant 3′-UTR of FASN containing the predicted binding site of miR-195 was inserted into the pmirGLO reporter vector (Promega, Madison, WI, USA). Cells were cotransfected with pmirGLO reporter vector and miR-195 mimics or inhibitor using Lipofectamine 2000 (Invitrogen) and incubated for 48 h. For measurement of Wnt signaling activity, cells were cotransfected with T-cell factor (TCF)/β-catenin vector, phRL-TK vector (Promega), and miR-195 mimics or inhibitor using Lipofectamine 2000 (Invitrogen) and cultured for 48 h. The luciferase activity was assessed using a Dual Luciferase Reporter Assay Kit (Promega) in accordance with the manufacturer’s protocol.
Data analysis
Experimental data were represented as mean ± standard deviation. Differences between the two groups were compared using Student’s t-test. Pairwise multiple comparisons were performed using one-way analysis of variance with Bonferroni post hoc test. Spearman’s rank test was used for correlation analysis. Statistical analyses were performed with SPSS 18.0 statistic software (SPSS Inc., Chicago, IL, USA). Values of p < 0.05 were considered as statistical significant.
Results
miR-195 is aberrantly downregulated in pancreatic cancer tissues and cell lines
To investigate the potential significance of miR-195 in pancreatic cancer, we first detected its expression pattern in pancreatic cancer tissues by RT-qPCR. We found that miR-195 expression was significantly decreased in pancreatic cancer tissues compared with adjacent normal tissues (Figure 1(a)). Furthermore, we examined the expression pattern of miR-195 in pancreatic cancer cell lines. We observed a significant decrease in miR-195 expression in pancreatic cancer cell lines compared to normal cells (Figure 1(b)). These findings suggest that the downregulation of miR-195 may be involved in the carcinogenesis of pancreatic cancer. The PANC-1 and AsPC-1 cell lines exhibited the lowest levels of miR-195 expression and were therefore selected for use in subsequent experiments.

Decreased expression of miR-195 in pancreatic cancer. (a) The expression level of miR-195 in pancreatic cancer tissues and adjacent normal tissues was determined by RT-qPCR. *p < 0.05. (b) The expression level of miR-195 in pancreatic cancer cell lines PANC-1, AsPC-1, BxPC-3, and SW1990 and normal pancreatic ductal epithelial (HPNE) cells was determined by RT-qPCR. *p < 0.05 versus HPNE.
Overexpression of miR-195 suppresses the proliferation and invasion of pancreatic cancer cells
To investigate the precise biological function of miR-195 in pancreatic cancer, we restored miR-195 expression in PANC-1 and AsPC-1 cells by transfecting miR-195 mimics. We found that overexpression of miR-195 significantly reduced cell proliferation (Figure 2(a)) and colony formation (Figure 2(b)) of pancreatic cancer cells. Overexpression of miR-195 markedly induced cell cycle arrest in the G0/G1 phase (Figure 2(c)). Next, we investigated the effect of miR-195 overexpression on cell invasion using a Transwell assay. We found that overexpression of miR-195 significantly repressed cell invasion of pancreatic cancer cells (Figure 2(d)). Whereas the suppression of miR-195 by transfection with the miR-195 inhibitor had the opposite effect (Figure 2(a)–(d)). Overall, these findings suggest that miR-195 exerts antitumor effects through inhibiting cell proliferation and invasion of pancreatic cancer cells.
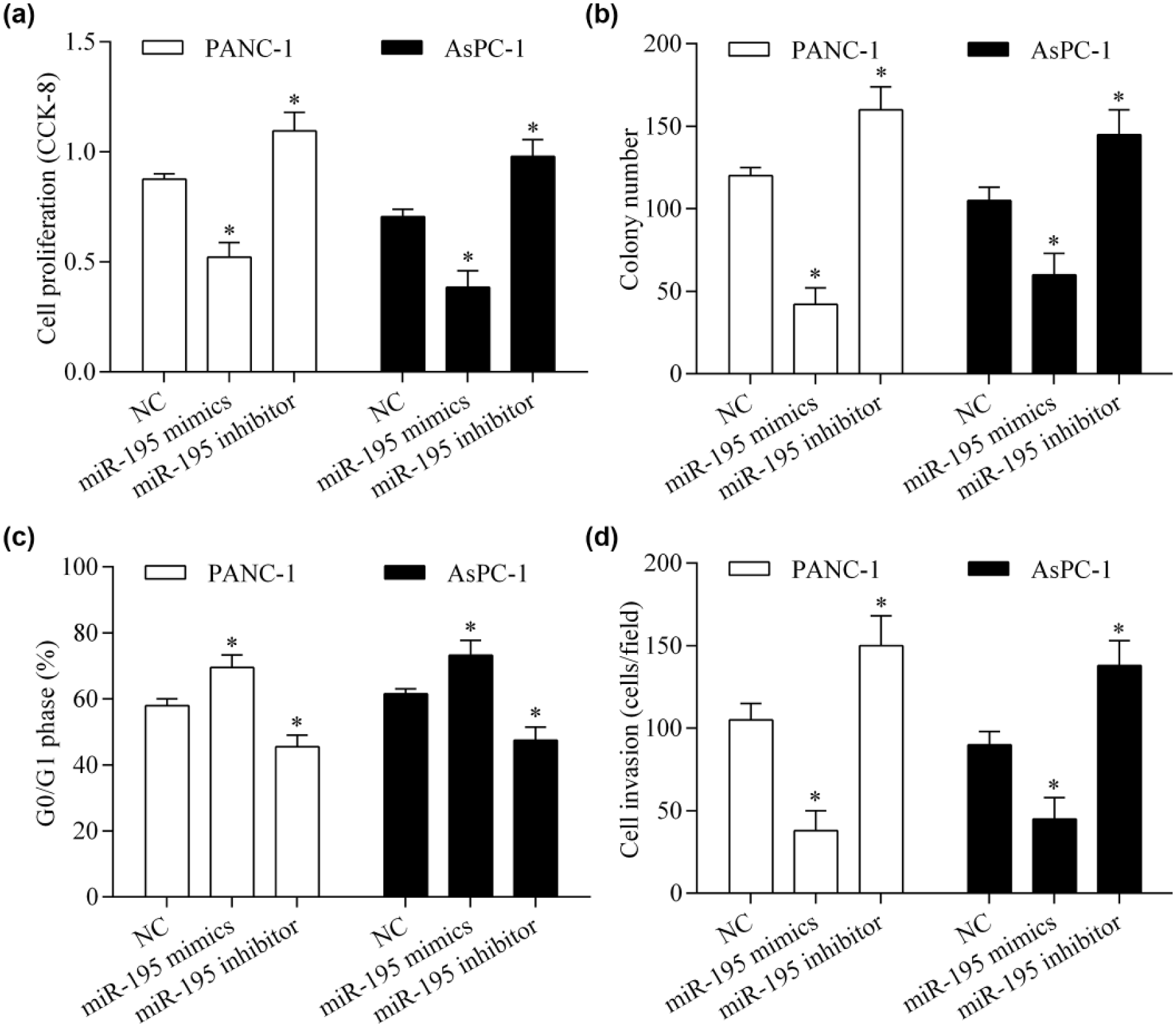
miR-195 inhibits pancreatic cancer cell proliferation and invasion. (a) The effect of miR-195 on pancreatic cancer proliferation was tested using a CCK-8 assay. PANC-1 and AsPC-1 cells were transfected with miR-195 mimics or inhibitor for 48 h. (b) Colony formation was measured by colony formation assay. Cells (transfected for 48 h) were seeded into six-well plates coated with 0.6% agarose for 14 days. (c) The proportion of cells in the G0/G1 phase was detected by flow cytometric analysis. (d) Cell invasion was detected using the Transwell invasion assay. *p < 0.05 versus NC.
FASN is a downstream target gene of miR-195 in pancreatic cancer cells
To elucidate the underlying mechanism by which miR-195 inhibits pancreatic cancer cell proliferation and invasion, we investigated the candidate targets for miR-195 using a bioinformatics approach. We found that FASN was a potential target gene of miR-195. The 3′-UTR of FASN mRNA contained two predicted binding sites for miR-195 (Figure 3(a)). To validate whether miR-195 directly targeted the 3′-UTR of FASN, we performed dual-luciferase reporter assays. We found that overexpression of miR-195 significantly reduced the luciferase activity of reporter vector containing the wild-type 3′-UTR of FASN, whereas suppression of miR-195 increased the luciferase activity (Figure 3(b) and (c)). However, these effects were abolished by mutation of the miR-195 binding sites (Figure 3(b) and (c)). To further investigate the direct regulatory effect of miR-195 on FASN expression, we performed RT-qPCR and western blot experiments. We found that the mRNA (Figure 4(a)) and protein (Figure 4(b)) expression levels of FASN were suppressed by miR-195 overexpression. By contrast, suppression of miR-195 promoted FASN expression (Figure 4(a) and (b)). Next, we tested the correlation between miR-195 and FASN in pancreatic cancer tissues. We found that the FASN mRNA expression level was upregulated in pancreatic cancer (Figure 5(a)). Correlation analysis showed that miR-195 was inversely correlated with FASN mRNA expression in pancreatic cancer tissues (Figure 5(b)). Taken together, these data indicate that FASN is a direct target gene of miR-195 in pancreatic cancer cells.

miR-195 targets the 3′-UTR of FASN. (a) The target sites for miR-195 in the 3′-UTR of FASN. Dual-luciferase reporter assay in (b) PANC-1 and (c) AsPC-1 cells. Cells were cotransfected with a reporter vector containing wild-type or mutant 3′-UTR of FASN and miR-195 mimics or inhibitor for 48 h. *p < 0.05 versus NC.
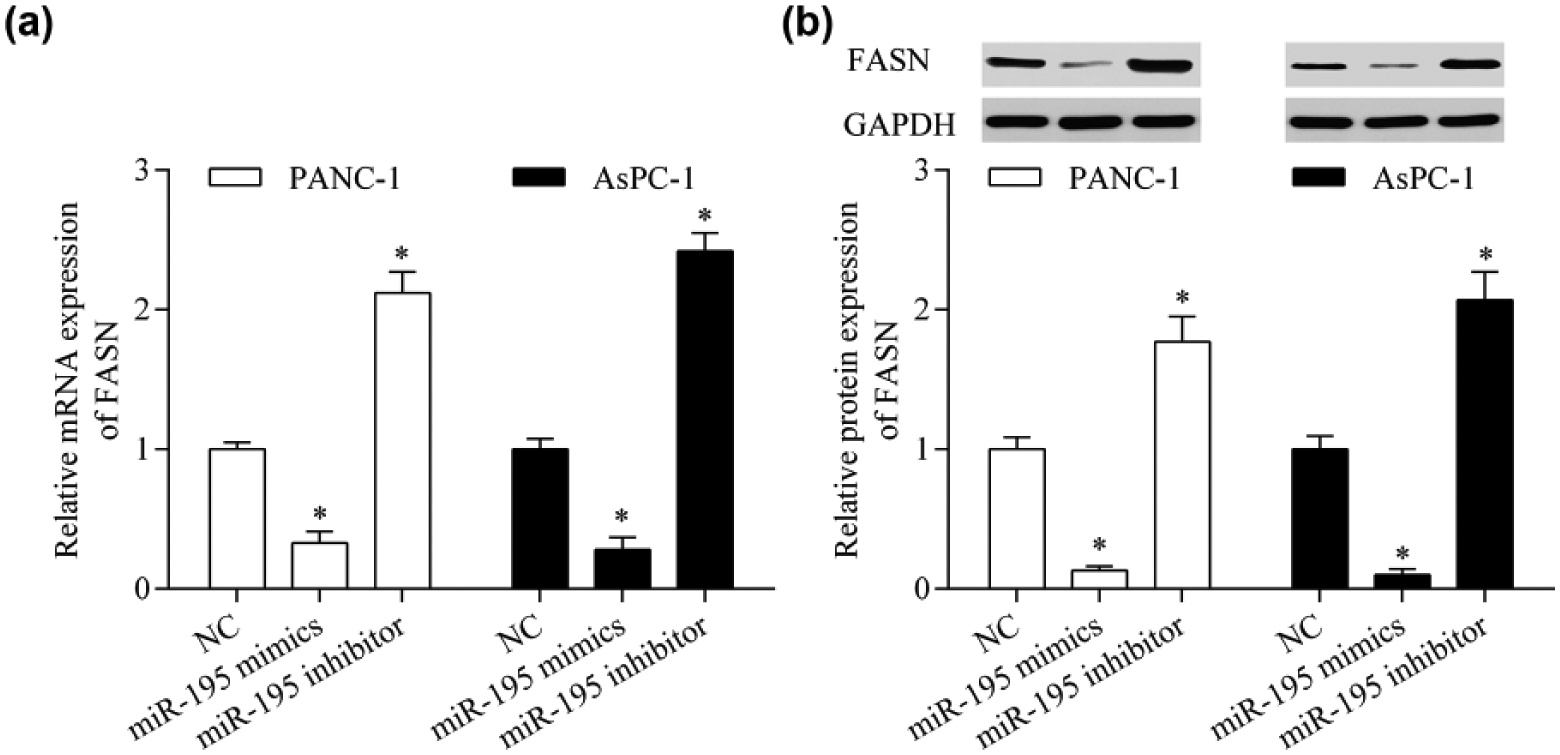
miR-195 suppresses FASN expression. PANC-1 and AsPC-1 cells were transfected with miR-195 mimics or inhibitor for 48 h. (a) The mRNA expression of FASN was detected by RT-qPCR. (b) The protein expression of FASN was assessed by western blot. *p < 0.05 versus NC.

miR-195 is inversely correlated with FASN expression in pancreatic cancer tissues. (a) RT-qPCR analysis of FASN mRNA expression in pancreatic cancer tissues and adjacent normal tissues. *p < 0.05 versus normal. (b) Spearman’s rank test of miR-195 and FASN expression in pancreatic cancer tissues. r = −0.8937; p < 0.0001.
Overexpression of miR-195 represses Wnt signaling in pancreatic cancer cells
To further investigate the molecular basis of miR-195-mediated antitumor effects, we detected the role of miR-195 in regulating Wnt signaling. We found that miR-195 overexpression significantly inhibited Wnt signaling activity (Figure 6(a)) and decreased the expression of c-myc (Figure 6(b)), a downstream target of Wnt signaling. By contrast, suppression of miR-195 produced the opposite effects. These data indicate that miR-195 represses Wnt signaling in pancreatic cancer cells.

miR-195 represses Wnt signaling in pancreatic cancer cells. (a) Dual-luciferase reporter assays of Wnt signaling activity. PANC-1 and AsPC-1 cells were cotransfected with TCF/β-catenin vector, phRL-TK vector, and either miR-195 mimics or inhibitor for 48 h. (b) The mRNA expression of c-myc was detected by RT-qPCR. *p < 0.05 versus NC.
Overexpression of FASN reverses the antitumor effects of miR-195 in pancreatic cancer cells
To further confirm whether miR-195 exerts antitumor effects through targeting FASN, we restored FASN expression in miR-195 mimics-transfected cells. We found that transfection of the pcDNA3.1/FASN expression vector (without 3′-UTR) significantly upregulated FASN expression in miR-195 mimics-transfected cells (Figure 7(a)). The inhibitor effects of miR-195 on Wnt signaling activity were significantly abolished by FASN overexpression (Figure 7(b)). Furthermore, the antitumor effect of miR-195 on cell proliferation (Figure 8(a)) and invasion (Figure 8(b)) was significantly reversed by FASN overexpression. Taken together, these results suggest that miR-195 inhibits pancreatic cancer cell proliferation and invasion by inhibiting FASN expression.

Restoration of FASN expression reverses the inhibitory effect of miR-195 on Wnt signaling. (a) Protein expression of FASN was detected by western blot in PANC-1 and AsPC-1 cells cotransfected with miR-195 mimics and pcDNA3.1/FASN expression vector (without 3′-UTR) for 48 h. (b) Dual-luciferase reporter assays of Wnt signaling activity. PANC-1 and AsPC-1 cells were cotransfected with TCF/β-catenin vector, phRL-TK vector, miR-195 mimics, and pcDNA3.1/FASN vector for 48 h. *p < 0.05.

Restoration of FASN expression reverses the inhibitory effect of miR-195 on pancreatic cancer cell proliferation and invasion. PANC-1 and AsPC-1 cells cotransfected with miR-195 mimics and pcDNA3.1/FASN expression vector (without 3′-UTR) for 48 h. (a) Cell proliferation was detected by CCK-8 assay. (b) Cell invasion was detected using the Transwell invasion assay. *p < 0.05.
Discussion
The dysregulation of miRNAs has been suggested to contribute to the development and progression of pancreatic cancer.10,11 Here, we found that miR-195 was downregulated in pancreatic cancer tissues and cell lines. We show that miR-195 inhibits pancreatic cancer cell proliferation and invasion by targeting FASN. Overexpression of FASN reversed the suppressive effects of miR-195. These results suggest a tumor suppressive role of miR-195 in pancreatic cancer.
A growing body of studies have suggested miR-195 as an important regulator in various human cancers. Various studies have shown that miR-195 suppresses the tumor progression of colorectal cancer by targeting CARMA3, 33 BCL2L2, 34 Bcl-2, 35 CARM1, 36 fibroblast growth factor 2, 31 and Raf1. 37 A tumor suppressive role of miR-195 was also evidenced in hepatocellular carcinoma, which can target and inhibit the expression of PHD finger protein 19, 38 chromobox homolog 4, 39 Wnt3a 40 and steroid receptor coactivator-3 protein. 41 Moreover, miR-195 is also reported as a tumor suppressor in many other cancer types, including prostate, 42 breast, 29 cervical, 43 glioma, 44 and lung 45 cancers by targeting various oncogenes. In a recent study, Rachagani et al. 32 report that miR-195 expression is decreased in a mouse model of pancreatic cancer. Consistent with previous studies, our results demonstrate that miR-195 is significantly decreased in pancreatic cancer tissues and cell lines. Overexpression of miR-195 inhibits the cell proliferation and invasion of pancreatic cancer cells. Our findings further confirm that miR-195 is a tumor suppressor.
In recent years, FASN has been well recognized as an oncogene in various cancers. 46 FASN is overexpressed in pancreatic cancer tissues, and high levels of FASN is correlated with high histological grade, frequent recurrence, and poor prognosis.25,26 Interestingly, FASN expression is also highly upregulated in the serum of pancreatic cancer patients, suggesting a potential application in diagnosis. 24 Overexpression of FASN promotes the tumor growth of pancreatic cancer. 47 Inhibition of FASN promotes cell apoptosis and suppresses cell proliferation of pancreatic cancer cells. 48 Moreover, knockdown of FASN increases the sensitivity of pancreatic cancer cells to chemotherapeutic drug and radiation treatments. 49 FASN also maintains the survival of pancreatic cancer stem cells. 50 Therefore, FASN may serve as a potential therapeutic target for pancreatic cancer. Here, we identified FASN as a downstream target gene of miR-195. We show that miR-195 negatively regulates FASN expression in pancreatic cancer. Inhibition of FASN by miR-195 represses the proliferation and invasion of pancreatic cancer cells. We also show that FASN expression is inversely correlated with miR-195 expression in pancreatic cancer tissues. This decreased miR-195 expression may contribute to the high expression of FASN during the development of pancreatic cancer. Thus, our study demonstrates a new molecular mechanism for the dysregulated expression of FASN in pancreatic cancer.
FASN can catalyze the synthesis of palmitate, which is critical for activation of various oncogenic signaling pathways, such as the Wnt signaling pathway. 16 FASN overexpression promotes the palmitoylation of Wnt-1 and the activation of the Wnt/β-catenin signaling pathway. 23 In this study, we show that inhibiting FASN expression by miR-195 overexpression inhibits the Wnt signaling pathway in pancreatic cancer cells, suggesting a novel approach for preventing Wnt signaling in cancer.
Various inhibitors of FASN have been developed, which show preclinical antitumor activity in cancer cell lines.51,52 However, these chemical inhibitors have limited clinical usefulness because of their side effects. miRNAs have been suggested as novel and promising inhibitors of FASN. Overexpression of miR-107 induces lipid accumulation in hepatocytes through the targeting of FASN. 53 miR-142-3p inhibits the proliferation of osteosarcoma cells by inhibiting FASN. 54 miR-193b is involved in metformin-induced cytotoxicity through the targeting of FASN in breast cancer cells. 55 Furthermore, FASN is suggested as a primary target of miR-15a and miR-16-1 in breast cancer. 56 In a recent study, Zhao et al. 57 report that miR-1207-5p suppresses the proliferation and invasion of hepatocellular carcinoma cells through the targeting of FASN. These reports suggest that FASN is epigenetically regulated by miRNAs. In this study, we identified FASN as a direct target gene of miR-195. Consistent with this finding, the functional relationship of miR-195 and FASN has been reported in osteosarcoma and breast cancer.58,59 Here, our findings further confirm that miR-195 targets and inhibits FASN expression. Therefore, miR-195 may represent an effective inhibitor of FASN, which may be used as a potential anticancer agent for FASN-associated cancers.
In conclusion, our study demonstrates that miR-195 inhibits the proliferation and invasion of pancreatic cancer cells by targeting FASN, suggesting a tumor suppressive role for miR-195. Our results show that miR-195 might function as an endogenic inhibitor of FASN, which could potentially be applied as a pancreatic cancer treatment.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Natural Science Foundation of Shaanxi Province (No. 2016JM8019), Sci-tech Research Development Program of Shaanxi Province (No. 2012K13-01-10), National Natural Science Foundation of China (No. 81101523), Fundamental Research Funds for the Central Universities (No. xjj2017048), and Scientific Research Fund Project of The First Affiliated Hospital of Xi’an Jiaotong University (No. 2016QN-02).