
Abstract
Recent technological advancements have shown tremendous mechanistic accomplishments in our understanding of the mechanism of messenger RNA translation in eukaryotic cells. Eukaryotic messenger RNA translation is very complex process that includes four phases (initiation, elongation, termination, and ribosome recycling) and diverse mechanisms involving protein and non-protein molecules. Translation regulation is principally achieved during initiation step of translation, which is organized by multiple eukaryotic translation initiation factors. Eukaryotic translation initiation factor proteins help in stabilizing the formation of the functional ribosome around the start codon and provide regulatory mechanisms in translation initiation. Dysregulated messenger RNA translation is a common feature of tumorigenesis. Various oncogenic and tumor suppressive genes affect/are affected by the translation machinery, making the components of the translation apparatus promising therapeutic targets for the novel anticancer drug. This review provides details on the role of eukaryotic translation initiation factors in messenger RNA translation initiation, their contribution to onset and progression of tumor, and how dysregulated eukaryotic translation initiation factors can be used as a target to treat carcinogenesis.
Introduction
Genome-wide analysis of messenger RNA (mRNA) translation by ribosome profiling revealed that the cell proliferation controlling process is maintained by protein synthesis, which shows that translation importantly contributes to overall gene expression. 1 In translation process, regulation is mainly achieved during the initiation phase of translation, as initiation is the rate-limiting step and is highly regulated.2,3 Initiation rate can be altered either by alterations in initiation factors’ availability and activity regulations by key signaling pathways, such as phosphatidylinositol-3-OH kinase (PI3K)/AKT, mammalian target of rapamycin (mTOR), and mitogen-activated protein kinase (MAPK) pathways, or highly structured 5′-untranslated region (5′-UTR). 4 Eukaryotic translation initiation factors (eIFs) are proteins that help in stabilizing the formation of the functional ribosomal complex around start codon and providing the regulatory machinery in translation initiation.
Currently, there are six eIFs reported to be involved in translation initiation, and most of the eIFs are a protein complex of several subunits, and dysregulations in their expression by overexpression, downregulation, or phosphorylation lead to oncogenic progression. Table 1 lists the general functions of these eIFs or their subunits in translation initiation.
Functions of eIFs and subunits in translation initiation.

eIF: eukaryotic translation initiation factor; mRNA: messenger RNA; GTP: guanosine triphosphate; GDP: guanosine diphosphate; GAP: GTPase activating protein; tRNA: transfer RNA; TC: ternary complex; PIC: pre-initiation complex; ATP: adenosine triphosphate; m7G: 7-methylguanosine; UTR: untranslated region; PABP: poly(A)-binding protein; ATPase: adenosine triphosphatase; GTPase: guanosine triphosphatase.
The mRNA must be recruited to ribosomes in order to decode information stored in mRNA The mRNAs have been known by two key features, 5′ 7-methylguanosine (m7G) cap (eIF4A, eIF4B, eIF4E, and eIF4G)25,26 and 3′ poly(A) tail, 27 which help in protecting mRNA from degradation and promoting translation initiation 28 (Figure 1). During translation initiation, mRNA is recruited to the 40S ribosome; the start codon and the 60S ribosome join to form an elongation-competent 80S ribosome. The 80S ribosome is functionally assembled on mRNA and the translation is ready to go 29 (Figure 2). The 43S pre-initiation complex (PIC) comprises ternary complex (TC) (eIF2-GTP-Met-tRNAiMet), eIF1, eIF1A, eIF3, and small ribosomal subunit 40S and probably eIF5.29,30 Each factor in this complex plays a vital role in translation initiation; eIF1 and eIF1A scan the mRNA for first start codon (AUG) in 5′ UTR region till complementary tRNA anticodon loops to first AUG; the scanning is achieved by altering the structure of mRNA-binding cleft. EIF3 acts as a scaffold and also interacts with the 40S ribosome and also interacts with eIF4F complex (eIF4A, eIF4E, and eIF4G) via eIF4G. 31 The eIF4F complex is required for a number of steps in translation initiation, including recruiting the 43S PIC to the 5′ cap structure of messenger RNA to promote ribosomal scanning along the mRNA to reach an AUG start codon.

The mRNA 5′ end contains a 7-methylguanosine (m7G) cap and the 3′ end of a poly(A) tail. The coding frame is the region of the mRNA that encodes the protein; usually begins with an AUG start codon and ends with one of three possible stop codons (UGA is one of the examples). 5′ UTR region lies between the cap and the start codon and 3′ UTR region is found between the stop codon and poly(A) tail.

Pathway of eukaryotic translation initiation: translation is a cyclical process, in which termination follows elongation and leads to recycling, which generates separated ribosomal subunits. Eukaryotic translation initiation is divided into six phases: (1) starting from formation of a 43S PIC consisting of a 40S subunit, eIF2.GTP-Met-tRNAMeti eIF1, eIF1A, eIF3, and possibly eIF5; (2) attachment of the 43S complex to unwounded mRNA region; (3) scanning of the 5′ UTR in a 5′ to 3′ direction by 43S complexes; (4) recognition of the initiation codon and 48S initiation complex formation; (5) joining of 60S subunits to 48S complexes and sequent detachment of eIF2–GDP and other translation factors mediated by eIF5B; and (6) GTP hydrolysis by eIF5B and release of eIF1A and GDP-bound eIF5B from assembled elongation-competent 80S ribosomes.
Dysregulations in translation initiation are most frequently implicated in cancer development and progression and is of utmost interest for targeting cancer. In this review, we tried to provide the detailed up-to-date understandings on the roles of eIFs in translation initiation, explored aberrant expressions of eIFs and contribution to the onset and progression of tumorigenesis and how dysregulated eIFs can be used as a target to treat carcinogenesis.
EIFs and cancer
Dysregulated mRNA translation plays an important role in the etiology and pathogenesis of malignancies. Tumor growth and cellular transformation are promoted by the aberrant translational pathway. In non-cancerous cells, this pathway is sensitive to cellular conditions such as availability of nutrients, energy or stress, ribosome production, and gene expressions, but in cancerous cells, it is hyper-activated and pro-oncogenic.32,33 Studies in the past two decades showed that a number of initiation factors (higher expressions levels of eIF4A, eIF4E, eIF4G, as well as low expressions levels of 4E-BPs or phosphorylation eIF2) are involved in various types of cancer.34–37
EIF1 and eIF1A or eIF1AX
EIF1 is a 113-amino-acid-long protein and eIF1A is a 144-amino-acid-long protein which are encoded on chromosome X; therefore, they are also known as x-linked eIF1a or eIF1AX. Both proteins are termed to be necessary for protein translation initiation in mRNA screening, delivery of tRNA, and recognition of start codon.30,38 These proteins are an important part of 43S PIC 2 (Figure 2) that binds to 5′m7G cap and shifts to the first AUG initiation codon. EIF1 binds to the region that is near to ribosomal P-site and eIF1A binds near the ribosomal A-site region, in structurally and functionally similar way related to bacterial IF3 and IF1, respectively. 39 Functionally, eIF1 and eIF1A have different roles, AUG recognition and formation of PIC, respectively, but interactively, both are required for the formation of 48S complex and binding at the initiation codon.40,41 The study using nuclear magnetic resonance (NMR) technology has shown that the interaction between eIF1A and eIF5 stops the eIF1 to scan 43S mRNA PIC. The interaction was studied and validated between the eIF1A and eIF5’s carboxyl-terminal domain (eIF5-CTD), as the eIF1A did not bind to mutant eIF5-CTD. The interaction is conserved between humans and yeast. 40
A recent study on eIF1 and eIF1A was carried out to evaluate the energetics of initiator tRNA binding to different near-cognate codons on the yeast 40S ribosomal subunit. The results revealed that the eIF1 and eIF1A together induce a comparatively uniform and high discrimination against near-cognate codons; that is, the eIF1 boosts the discrimination against a first position near-cognate G-U mismatch and also against a second position A-A base pair, although eIF1A mainly acts on third codon position. 42 In another study, Pisareva and Pisarev proposed that the contact between eIF1A OB domain and unique DHX29 N-terminal domain near mRNA entrance site induces the stimulatory recognition of near-cognate CUG codon, as DHX29 is known to stimulate the recognition of the AUG codon, but not the near-cognate CUG codon, regardless of its nucleotide context during ribosomal scanning. 43
Dysregulation in micro-RNA (miRNA) biogenesis is associated with various human diseases, particularly cancer. Ago2 gene is required for RNA-mediated gene silencing (RNAi) by the RNA-induced silencing complex (RISC). Biochemical and NMR studies have shown the interaction between the eIF1A and Ago2, and the activity of Ago2 in RNAi and miR-451 biogenesis is promoted by this interaction. EIF1A binds to the middle domain of Ago 2 (MID), and this interaction does not vitiate translation initiation. 44 Table 2 shows the role of eIF1 and eIF1A/eIF1AX in tumorigenesis.
Aberrant expression of eIF1 and eIF1A/eIF1AX in human cancer.

EIF2
EIF2 is a multimeric guanosine triphosphatase, comprising the three-subunit complex (eIF2α, eIF2β, and eIF2γ) that forms the core of the GTP/eIF/Met-tRNAiMet TC, which is crucial for translation initiation. After formation, TC combines with the 43S ribosomal unit to form 43S PIC. EIF2 cycles between the GTP-bound active and GDP-bound inactive forms, and this GTP/GDP cycle is regulated primarily through reversible phosphorylation of eIF2α.
In response to phosphorylation of eIF2α, eIF2 is locked in GDP-bound inactive form resulting in reducing the translation, and in all eukaryotes, this is a major cell stress response to preserve their energy sources to overcome the stress. 52 Certain stress responses such as oxidizing agents, nutrient deprivation, heat shock, heavy metals, high salt levels, and hypoxia have been reported to cause the phosphorylation of eIF2α. 32 Phosphorylation of eIF2α can be induced by four stress-responsive kinases: PERK kinase, 53 GCN2 kinase, 54 PKR kinase, 55 and HRI kinase. 56 Studies have shown that eIF2α phosphorylation is responsible for protein synthesis inhibition when the different cell lines exposed to ultraviolet (UV) light, including fission yeast,57,58 rat fibroblast, 59 human cells, 60 maize leaves, 61 and mouse embryonic fibroblasts.62,63 Inversely, the nonphosphorylation of eIF2α increases protein synthesis and supports cell proliferation.
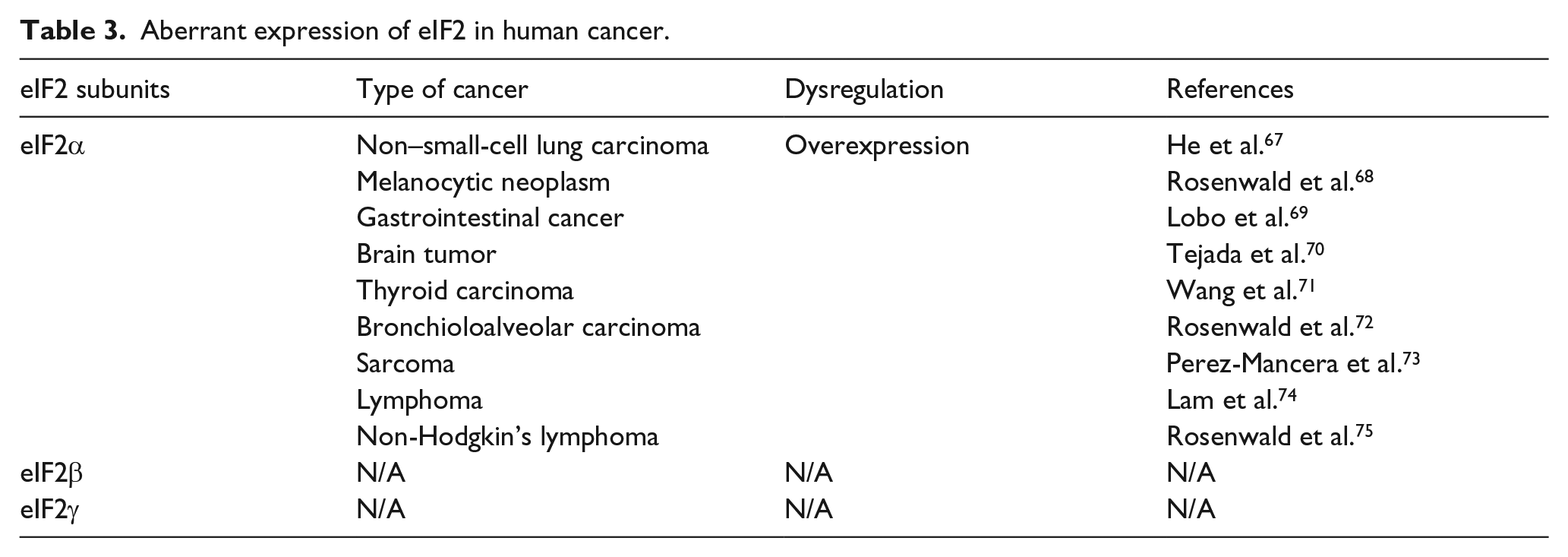
The eIF2 complex, more specifically eIF2α, appears to be an important hub of translation that participates in cell proliferation and survival, malignant transformation, tumor initiation, progression, and metastasis. EIF2α regulation role in different carcinomas is signified by the fact that eIF2α’s nonphosphorylatable mutant (eIF2α S51A) overexpression enhances the amount of the TC leading to unrestricted translation initiation, which results in the transformation of normal cells to malignant cells. 64 Even if eIF2α dephosphorylation is not sufficient for inducing malignant transformation by itself, in combination with oncogenic stimuli, it may lead to cellular transformation. 65 In another study, eIF2αS51P (eIF2α-serine 51-phosphorylation) is evident to play an important role in stress-induced tumorigenesis, which proposed that it may have prognostic value and a suitable target for the design and implementation of effective anti-tumor therapies. 66 Table 3 summarizes the role of eIF2 aberrant expression in various tumors.
Aberrant expression of eIF2 in human cancer.
EIF3
Mammalian eIF3 is the largest eIF protein complex (~804 kDa) composed of 13 non-identical subunits (eIF3a-m) with different masses.76,77 Others species such as Drosophila melanogaster and Schizosaccharomyces pombe (fission yeast) also contain these similar eIF3 subunits, but Saccharomyces cerevisiae contains only five core subunits of eIF3 (eIF3a, eIF3b, eIF3c, eIF3g, and eIF3i). In vitro initiation assay in yeast with only these five core subunits of eIF3 suggested that these five mammalian eIF3 subunits may play the major role of its functions, while the other subunits may act as regulators. 78 Binding between eIF3 and 40S ribosomal subunits is stabilized by the eIF3j. 79 This binding may be an early event, as a study in tumor cells showed that eIF3–40S complexes lack Met-tRNAi and mRNA. 80 Cryoelectron microscopy model studies of eIF3–40S complexes showed eIF3 as a solvent in eIF3–40S complex. 81 Significantly, eIF3 binds to eIF4G, which plays an important role in recruiting the 40S subunit to the mRNA. 82 EIF3 complex and its subunits individually bind to other initiation factors to organize the important components of translational machinery on or near the surface of 40S ribosomal subunit for its proper working: eIF3 complex binds to eIF5 and eIF2β, 83 eIF3c binds to eIF1 and eIF5, 84 and eIF3g binds to eIF4B. 85 Despite eIF3’s binding ability to other initiation factors, subunits eIF3a and eIF3d bind to the ribosome’s exit region on mRNA, giving the signal that mRNA dissociation is premature. 86 Despite all information about eIF3, it is unclear that how all the subunits of eIF3 organize the different steps in translation initiation pathways. Due to its broad contributions in initiation pathways, altered expression of eIF3 and/or its subunits may contribute to different malignant transformations (Table 4).
Aberrant expression of eIF3 in human cancer.

EIF4
EIF4 is the first reported translation initiation factor with multiple subunits working together and is known to recruit ribosomal subunits to mRNA.116,117
EIF4A (46.2 kDa) belongs to the family of DEAD or DEAH (aspartic acid (Asp, D), glutamic acid (Glu, E), alanine (Ala, A), and histidine (His, H)) box helicases.118,119 Additional domains of eIF4A help its interaction with eIF4B and eIF4H, 120 eIF4G, 15 PDCD4, 121 RNA, 122 and ATP. EIF4A more actively plays it role when it is a component of the eIF4F complex (eIF4A, eIF4E, and eIF4G)123,124 which is required for 43S PIC recruitment and scanning.
The role of eIF4B (69.2 kDa) in translation initiation is not well-understood as it is reported to be independent of the eIF4F complex and was identified as non-essential for translation initiation. A recent study showed that translation initiation eIF4B stimulates the eIF4A activity by regulating the eIF4A conformational cycle and by binding to the eIF3 complex through the eIF3A subunit. 122 The interaction of eIF4B and eIF3A is strengthened 125 upon eIF4B phosphorylation which is induced by activation of the S6K kinase. EIF4B is a downstream target of mTOR as well. 126 The chemo-resistance and metastasis characteristics of non–small-cell lung cancer (NSCLC) could be suppressed by miR-216a that targets the eIF4B and ZEB1. The miR-216a is a tumor-suppressive miRNA, and enforced expression of miR-216a is a novel therapeutic strategy for NSCLC. 127 EIF4B has also reported assuring cell survival by promoting particular translation of genes involved in cell proliferation (CDC25C, c-MYC, and ODC) and cell survival (BCL-2, XIAP). 128
EIF4E (25.1 kDa) is the cap-binding protein that specifically recognizes the 5′ m7G cap by cupping it between two conserved tryptophans found in the cap-binding pocket of eIF4E. Because of its rate-limiting nature, slight eIF4E overexpression (~2.5-fold) transforms normal cells into immortalized cells in transgenic mice. 129 EIF4E and its phosphorylated forms have been studied in different types of cancers. Overexpression of eIF4E accounted for approximately 30% cancer cases, it plays critical role mRNA recruitment that is one of the most important steps in translation initiation.130,131 EIF4E phosphorylation in response to extracellular stimulation leads to uncontrolled translation and proliferation and inhibits apoptosis. 132
The capability of EIF4E to build the cap-binding complex (eIF4F) and its availability is regulated by the 4E-binding protein 1 (4E-BP1). The interaction between eIF4E and 4E-BP1 is phosphorylation dependent.133,134 Activation of mTOR pathway diminishes the binding between eIF4E and 4E-BP1 by phosphorylation of 4E-BP1; as a result, the eIF4E is hyper-activated. This hyper-activation of eIF4E leads to translation initiation when it interacts with the eIF4F complex 135 ; hence, the hyper-phosphorylation of 4E-BP1 is associated with malignant progression of prostate, 136 colon, 137 breast,138–140 ovarian,141,142 lung, 36 head and neck cancers, 37 and lymphoma. 143
In vivo, eIF4E overexpression leads to the development of angiosarcoma, lung carcinoma, and lymphoma in transgenic mice. 129 One anti-tumor approach can be acquired by inhibiting cap-binding capability of eIF4E with compounds (4Ei-1 or ribaviiin) that interrupt the eIF4e–cap interaction, and a number of experiments have been carried out to achieve this particular goal. 144 In vitro study reported the significant reduction in migratory activity and invasiveness of breast cancer cells by eIF4e knockdown using an inducible RNAi approach and in mouse model had an inhibitory effect on tumor growth and metastasis, 145 and inhibition of hepatocellular carcinoma cells could be inhibited by artificial miRNA targeting eIF4E. 146
EIF4G (175.5 kDa) is a scaffold protein around which mRNA, ribosomal subunits, and the cap-binding complex assemble in order to accomplish translation. EIF4G is a component of cap-binding complex eIF4F and it enhances the association of eIF4E with the cap structure through allosteric mechanisms. 147 EIF4G has binding sites for poly(A)-binding protein (PABP) at its N-terminal, 32 eIF4E, 147 eIF4A, 122 and eIF3A. 12 In vitro study has shown the translation initiation inhibitor 4EGI-1 could reduce the eIF4G–eIF4E interaction in human melanoma and breast cancer cells and lead to growth inhibition of melanoma and breast xenograft tumors in mouse models. 148
EIF4H (27.5 kDa) is a co-factor of eIF4A. EIF4H and eIF4B have similar ability to stimulate the eIF4A’s RNA helicase activity in the absence of eIF4G, 149 but in eIF4G presence, eIF4A is more stimulated by eIF4B than eIF4H, suggesting that these two factors may have distinct roles in translation initiation. 150 EIF4H and eIF4B bind to the same domain of eIF4A 120 and the binding is enhanced in the presence of ATP. 151
Dysregulation in eIF4 expression has been reported to be associated with malignant transformation. EIF4A has been reported with overexpression in malignant hepatoma 152 and melanoma. 153 Overall, eIF4G and eIF4E are reported to associate with most of the malignant transformation. Table 5 depicts detailed alterations of eIF4 expression in carcinogenesis.
Aberrant expression of eIF4 in human cancer.

EIF5
EIF5 is 49 kDa protein and one of the most important proteins of translation initiation pathways possessing two functions: N-terminally it acts as GTPase-activating protein (GAP) (GTP hydrolyzation by eIF2), 182 and an independent GDP dissociation inhibitor (controlled recycling of eIF2). 183 Furthermore, eIF5 also reported as an inhibitor of eIF2B and holds the guanine nucleotide exchange factor activity that promotes eIF2 recycling. 22 Multifactor complex of eIF5 comprises the TC (eIF2-GTP-Met-tRNAi), eIF1A, eIF1, and eIF3. EIF1 binds the eIF5 to restrict its GAP activity 182 and helps in the accurate recognition of start codon. Overexpression of eIF5 and eIF5-mimic protein (5MP: 5MP1 and 5MP2) has been associated with the induced translation of ATF4 and possibly other genes with upstream open reading frames (uORFs) in their mRNA leaders through delayed re-initiation, thereby enhancing the survival of normal and cancer cells under stress conditions. 184
EIF5A (17 kDa) is the gene encoding eIF5 protein. In mammals, eIF5A has two isoforms, that is, eIF5A1 and eIF5A2, and these both isoforms undergo hypusine modification at the same specific lysine residue. 185 EIF5A1 is expressed in all tissues and cells and found abundantly in rapidly proliferating cells, but, on the other hand, eIF5A2 is expressed in a tissue-dependent manner and most of the time is hardly detectable. 186 In a recent study, eIF5A2 has been reported to promote the chemo-resistance to doxorubicin via regulation of epithelial–mesenchymal transition (EMT) in colon cancer cells, concluding that eIF5A2 inhibition may be a new potential strategy for the reversal of drug resistance in colorectal cancer therapy. 187
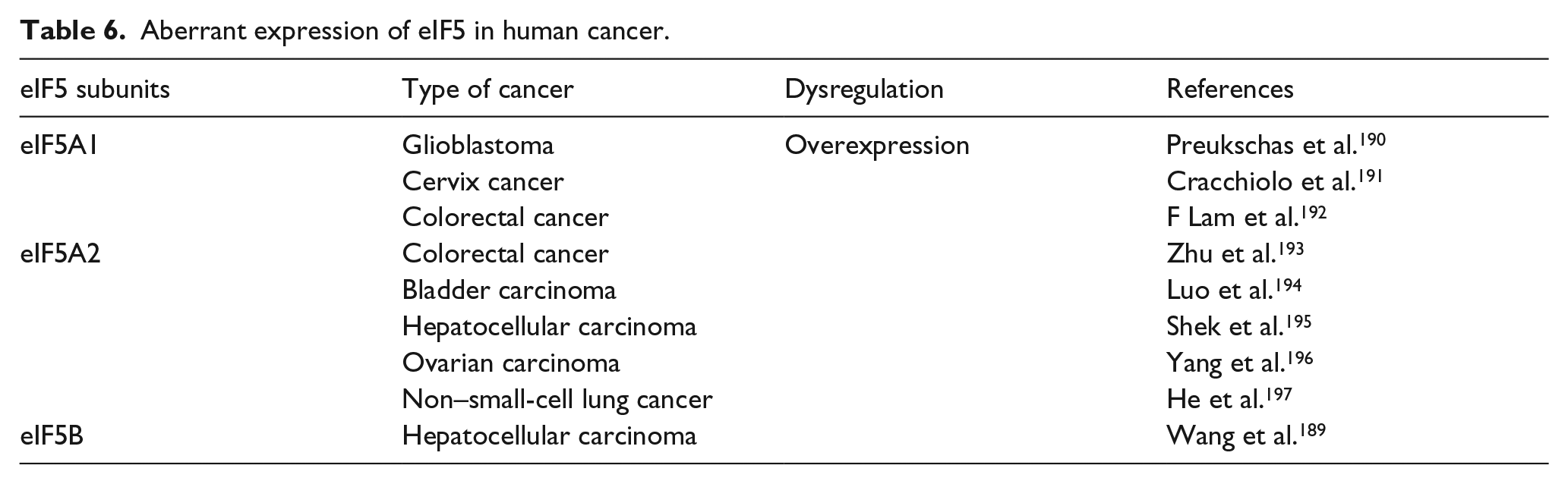
EIF5B (175 kDa) promotes 60S ribosome subunit joining and pre-40S subunit scanning and can indirectly support the Met-tRNAi association with the ribosome in translation initiation. 188 A recent study proposed that the eIF5B promotes hepatocellular carcinoma cell proliferation and migration in vitro and in vivo partly through increasing ASAP1 expression. 189 Aberrant expressions of eIF5 correlated with advanced tumor stage are described in Table 6.
Aberrant expression of eIF5 in human cancer.
EIF6
EIF6 is a rigid protein consisting of 245 amino acid residues. Among various factors that regulate ribosome biogenesis and translation, eIF6 has been identified to function in both processes. Translation regulation is carried out by rate-limiting nature of eIF6. It acts as a ribosomal anti-association factor in translation initiation,198,199 as it blocks the interaction between the 40S and 60S ribosomal subunits by binding to 60S ribosomes, leading to translation initiation blockade.200,201 With the cytoplasmic (70%) role of eIF6, it has also expressed in the nucleus (30%) by playing a role in maintaining biogenesis process of 60S ribosomal subunit.200,202 Reduced expression of eIF6 in human cell lines disclosed the repressive activity of miRNA, and it has a role as a regulator of translation at the post-transcriptional level and RNA stability. 203 The initiation factor eIF6 appears to participate in tumorigenesis. For long, eIF6 overexpression was thought to be a byproduct of uncontrolled growth rate by which cancers are characterized. EIF6 is overexpressed in different types of cancers: colorectal, highest level (up to 10-fold) of eIF6 is expressed, 204 malignant mesothelioma, 205 ovarian serous carcinoma, 206 acute promyelocytic leukemia, 207 head and neck carcinoma, 208 and lung metastasis. 209 Reduction in 50% expression of eIF6 led to 90% reduction in oncogene-mediated transformation by activated Ras and Myc. 210
Signaling pathways regulating translation in cancer
Signaling pathways which promote tumorigenesis possess growth factor signaling characteristic stimulated by activation of RAS signaling, 211 receptor tyrosine kinases (RTKs), 212 PI3K, 213 MAPKs,214,215 and AKT 216 signaling (Figure 3). These pathways also control translation by phosphorylation and regulate the protein synthesis factors or ribosomal proteins. mTORC1 directly and indirectly regulates various factors involved in translation initiation, such as eIF4B, eIF4E, 4E-binding proteins (4E-BP), eIF4G, ribosomal protein S6 (rpS6), and S6 kinase (S6K). 217 mTORC1 (mTOR, Raptor, and mLst8), a major regulator of translation, is negatively regulated by AKT signaling which deactivates the TSC1/2 complex. 218 On activation, mTORC1 is responsible for phosphorylation of eIF4E-binding proteins (4E-BP), which in result lead to release and activation of cap-binding protein eIF4E219,220 and leads to an increase in cap-dependent translation. 221 Various in vitro and in vivo studies have verified the clinical and biological importance of mTORC1-mediated 4E-BP phosphorylation, demonstrating that 4E-BPs are critical mediators of mTORC1 action.222–224 Hyperphosphorylated eIF4E-binding proteins’ inactivation is associated with elevated tumor grades and limits patient survival rate in breast,138,225 prostate, 136 gastric, 226 penile cancers, 227 leukemia, 228 and brain tumor. 70 Activated mTORC1 also induces global protein translation by phosphorylating the scaffold protein eIF4G and the S6K that regulates rpS6 and eIF4B translation initiation factor.229,230 EIF4G1, one of the eIF4G-related protein, is phosphorylated in an mTOR-dependent manner in response to extracellular stimuli (growth factors, serum, and insulin). Nevertheless, the importance of eIF4G1 phosphorylation is not significantly determined because no evidence has been associated with any changes in the activity of other translation initiation factors upon its phosphorylation. On the other hand, depletion of eIF4G1 has been reported with blocking of cell proliferation, decrease in cell size, and partial inhibition of translation231,232 suggesting that mTORC1-mediated eIF4G1 phosphorylation may have some effects on the promotion of cell proliferation and growth. RPS6 kinase proteins (S6K1 and S6K2) are a direct target of mTORC1 and regulate cell growth. 233 S6K1 phosphorylates and deactivates eEF2 kinase, which inhibits elongation factor eEF2, and PDCD4, leading to its proteasomal degradation. 234 PDCD4 is a tumor suppressor which inhibits both tumor promotion and tumor progression of carcinogenesis.235,236 Tumorigenesis is suppressed when PDCD4 binds with eIF4A by blocking RNA helicase eIF4A activity and hence inhibits translation of a set of mRNAs that require eIF4A activity. 237 Downregulated expression of PDCD4 is associated with numerous cancers, such as colorectal,238,239 renal adenocarcinoma, 240 pancreatic, 241 breast, 242 ovarian, 243 esophagus, 244 lung, 245 gastric, 246 hepatocellular carcinoma, 247 suggesting that PDCD4 is a significant tumor suppressor for many types of cancers. MTOR pathways also induce ribosomal biogenesis by regulating polymerases (Pol I and III) that generate ribosomal RNAs and by influencing ribosomal RNA (rRNA) processing.248,249 In tumorigenesis, mTORC1 can also be activated by MAPKs that act downstream of hyper-activated RTKs and RAS pathways. Extracellular signal–regulated kinases (ERKs) can directly phosphorylate and inhibit tuberin (TSC2), which further phosphorylate and irregulate TSC2 function 250 and additionally stimulate mTORC1 activity via direct raptor phosphorylation.251,252 AMP-activated protein kinase (AMPK) also inhibits mTORC1 activation by inducing mTORC1 negative regulator TSC1/TSC2 complex, a GAP. 253 mTORC2 (Rictor, Protor, mLst8, and Sin1) 220 insulin-stimulated activation through PI3K pathway is associated with ribosomes, as ribosomes are required for mTORC2 activation. mTORC2 controls various members of AGC subfamily of kinases such as Akt, SGK1, and protein kinase C alpha (PKC-alpha) by phosphorylation of Akt and AGC at hydrophobic motifs.254,255 Consequently, all these complex network-dependent manners indicate that signaling networks influence tumorigenesis by their effect on various important steps of translation in highly proliferating cancer cells.
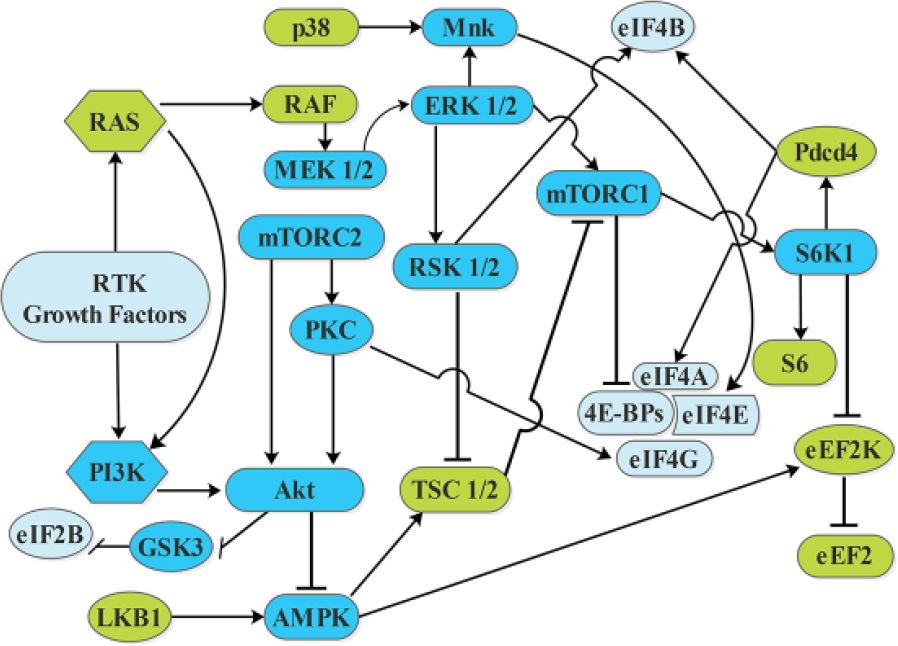
Regulatory networks involved in translation initiation and inhibition points. Arrows show activation and bars show inhibition.
Targeting dysregulated eIFs as cancer treatment
Current cancer research majorly unveils two vulnerabilities to which cancerous cells have become dependent on their stability and growth, that is, the gene networks (PI3K/AKT/mTOR pathway) and translation regulators. In the past decade, blocking of dysregulated signaling pathways by kinase inhibitors has shown great significant for cancer treatment. Figure 3 demonstrates the regulatory networks involved in translation initiation and inhibition points.
Current therapeutic strategies aimed at targeting translation initiation components as an anti-neoplastic approach focus on the rate-limiting eIF4F complex (eIF4E, eIF4A, and eIF4G), involved in the ribosome recruitment phase of translation initiation. The function of the eIF4F complex is to recruit small ribosomal subunits and related factors (43S PICs) to the 5′ end of mRNA. EIF4F complex–based approaches at therapeutic levels might include the following: (1) suppressing eIF4E activity, (2) targeting eIF4E phosphorylation, (3) intervening the eIF4G–eIF4E interaction, and (4) targeting the eIF4A helicase activity.
EIF4E is the least abundant initiation factor making cap-dependent initiation, the rate-limiting step of translation. 256 EIF4E suppression at the organism level is appeared to be well tolerated 257 and has also been modeled in the mouse exhibiting significantly delayed sporadic tumor onset. 258 Suppressing the eIF4E expression is an efficacious approach to reducing eIF4F complex levels, eIF4E is the least abundant (0.05 copies per ribosome) of all translation factors. 256 Approximately, 80% knockdown of eIF4E has been obtained using antisense oligonucleotides (4EASOs), while the synthesis of global protein is slightly affected (~20%). As a result, suppressed tumor growth in breast and prostate xenograft models has been reported by apoptosis. Additionally, the endothelial cell tube formation has also been inhibited by 4EASOs, suggesting a possible anti-angiogenic activity. 257 MAPK-interacting kinases, MNK1 and MNK2, phosphorylate the eIF4E on a single residue, Ser209. Experimental results revealed complete loss of eIF4E phosphorylation in the absence of MNK1 and MNK2 in the mouse model and have no observable phenotype, 259 although eIF4E phosphorylation appears essential for oncogenic activity. 177 These results attracted the scientists to develop MNK inhibitors as potential anti-neoplastic agents. To support this evidence that the effect of MNK inhibition on tumorigenesis is due to loss of eIF4E phosphorylation, mice were genetically modified with a nonphosphorylatable allele of eIF4E, Ser209Ala. Results demonstrated that the cells from these mice are comparatively more resistant to RAS-based oncogenicity than their wild-type counterparts. When crossed with mice, in which phosphatase and tensin homolog (PTEN) was deleted in the prostate, the development of invasive prostate neoplasia was significantly reduced in mice harboring the Ser209Ala allele. 177 The eIF4E-binding site on eIF4G has been well studied and consists of a Y(X)4LF motif (where X is any amino acid and F is hydrophobic). 4EGI-1 has been reported to inhibit eIF4E/eIF4G interaction 260 but it enhances eIF4E/4E-BP1 association. 4EGI-1 also effectively induces apoptosis in multiple myeloma cells through an NOXA-dependent induction261,262 and inhibits the growth of breast and melanoma cancer xenografts without showing any evident toxicity. 148 The 4EGI-1’s mode of action is reported as cap-dependent and cap-independent manner. 263 The RNA helicase eIF4A is the enzymatic subunit of the eIF4F complex. Three natural occurring products, hippuristanol (inhibits eIF4A–RNA interaction), 264 silvestrol (dimerization inducer, depletes eIF4A from the eIF4F complex), 265 and pateamine A (stimulates binding of eIF4A to RNA in a non-sequence–dependent manner), 266 interfere cap-dependent translation by interfering with eIF4A helicase activity and are currently being researched as chemotherapeutic agents. These natural products all perturb cap-dependent translation by interfering with eIF4A activity and are currently being explored as chemotherapeutic agents.
Future perspectives
Recent advancements have made substantial contributions in our understandings of mRNA translation in eukaryotic cells. More detailed studies of all these factors will provide better insight to understand the researchers that how the translation process is reprogrammed in malignancies and could provide important clues to deal cancer at therapeutic grounds. It is essential to relate future thermodynamic and kinetic frameworks to the translation process in cells so that a complete picture of translation can be achieved in normal and diseased states. Furthermore, advanced technologies such as single molecule approaches, cryoelectron microscopy, and biochemical mapping can anticipate deeper studies of binding sites of initiation factors on ribosomal subunits. Recently, high-resolution structures of the eukaryotic ribosome started to provide much needed structural information on which future genetic and biochemical experiments can be based. Ultimately, we can hope that a better understanding of the reprogrammed translation mechanism will provide new insights into possible targets for both diagnostics and therapeutics for different stages of cancer.
Footnotes
Acknowledgements
The authors would like to extend their gratitude to Miss Adeeba Mahmood for her help revising English language of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work is supported by the National Natural Science Foundation of China (grant no. 81172516).