
Abstract
This study aimed to explore the clinical significance and prognostic value of Fra-1 in hepatocellular carcinoma patients after curative resection. Fra-1 expression was investigated using a combination of techniques: immunohistochemistry for 66 samples of hepatocellular carcinoma and quantitative real-time polymerase chain reaction and western blotting assays for 19 matched hepatocellular carcinoma specimens. Fra-1 was present in 38 of 66 (57.6%) tumor tissues, with intense staining in the nuclei. There was also positive staining in 14 of 66 (21.2%) adjacent peritumoral tissues, with weak staining in the cytoplasm. Quantitative real-time polymerase chain reaction and western blotting assays confirmed higher expression of Fra-1 messenger RNA and Fra-1 protein in tumor tissues than adjacent non-tumor tissues for 19 hepatocellular carcinoma samples (p < 0.001). Positive expression of Fra-1 was significantly related to vascular invasion and serum alpha-fetoprotein. Kaplan–Meier survival analysis found that overexpressed Fra-1 was correlated with poor overall survival and disease-free survival. Multivariate analysis identified Fra-1 as an independent prognostic factor. Fra-1 may be involved in the progress of hepatocellular carcinoma and could be a promising molecular candidate in the diagnosis and treatment of hepatocellular carcinoma.
Introduction
Hepatocellular carcinoma (HCC) is the third leading cause of death from cancer worldwide. 1 It is attributed to several genetic mutations and epigenetic alterations. Hepatitis B virus (HBV) infections, alcohol, aflatoxins, obesity, and diabetes are other risk factors of HCC. 2 Despite the fact that there are many new methods used in the diagnosis and treatment of HCC, the prognosis remains poor because of high recurrence rates and metastasis. 3 An important reason for the poor prognosis is due to lack of a precise HCC prognostic grading system and an effective early HCC screening method. 4 Therefore, evaluation of potential biomarkers in the diagnosis and treatment of HCC is justified.
Activator protein-1 (AP-1) which consists of Jun (c-jun, JunB, and JunD) and Fos proteins (c-fos, fosB, fra-1, and fra-2) is a dimeric transcription factor. Each Fos-related protein can form a heterodimer with Jun family proteins and bind to very similar or identical AP-1 sites. The formation of heterodimer and DNA-binding activity which are prerequisites for transforming activity are specified by bZIP motif of Fos proteins. 5 FosB can mediate transcription in vitro, while FosB2, a C-terminally truncated form of FosB, cannot, although both proteins can mediate DNA binding with Jun proteins, which shows that the transcriptional activation domain is in the C terminus of FosB. 6 These molecules are involved in multiple physiological processes, including stress responses, cell proliferation, differentiation, inflammation, and cancer development.7,8 Fosl1 (Fra-1 gene) was initially described as an immediate early gene that is induced by serum and related to the c-Fos. 9 Fra-1 has been proposed to inhibit the activity of AP-1 due to the lack of a C-terminal transactivation domain (TAD). 10 Kovary and Bravo 11 reported that when serum-starved fibroblasts were stimulated by serum, the abundant Fos proteins were c-Fos and FosB at early times whereas Fra-1 and Fra-2 at later times. This shift might help to limit the duration of the AP-1 response. Bergers et al. 12 showed that Fra-1 could transform rat fibroblasts in vitro and contribute to tumor development in athymic mice. Recent studies showed that Fra-1 played a critical role in cancers involving the lung, 13 colon, 14 breast, 15 thyroid, 16 and other organs, suggesting that it may contribute to the development and progression of several malignant tumor types. In this context, we analyzed the expression of Fra-1 by immunohistochemistry, quantitative real-time polymerase chain reaction (qRT-PCR), and western blotting to assess its clinical significance in HCC.
Materials and methods
Tissue samples
Tissue specimens for immunohistochemistry analysis were collected from 66 patients with HCC who received curative surgery between 2008 and 2011 at the Department of Hepatic Surgery at Affiliated Anhui Provincial Hospital of Anhui Medical University, Hefei, China. These patients consisted of 59 males and 7 females, whose mean age was 55 years (range: 20−80 years). Clinicopathological parameters of all the patients such as gender, age, serum alpha-fetoprotein (sAFP), hepatitis B surface antigen (HBsAg), tumor size, vascular invasion, tumor capsule, status of metastasis, tumor–node–metastasis (TNM) staging, Edmondson classification system, cirrhosis, and Barcelona Clinic Liver Cancer (BCLC) grading system were gathered retrospectively. In addition, 19 pairs of snap-frozen tumor and adjacent peritumoral tissues (at least 2 cm away from tumor margin) of HCC were obtained for Fra-1 analysis by western blotting and qRT-PCR techniques. No anti-cancer treatment was carried out for the patients. Detailed information of the patients is displayed in Table 1. All patients were followed up until May 2016. Median follow-up was 21 months (range: 2–60 months). Overall survival (OS) time was defined as the interval between the date of surgery and the date of death or the last visit. Disease-free survival (DFS) time was defined as the time from the date of surgery to the date of recurrence which was diagnosed definitely for the first time. Informed consents were signed by all patients before surgery, and the study protocol was also approved by the Human Research Ethics Committee of Anhui Medical University.
Correlations between Fra-1 expression and clinicopathological characteristics of HCC (N = 66).

HCC: hepatocellular carcinoma; sAFP: serum alpha-fetoprotein; HbsAg: hepatitis B surface antigen; TNM: tumor–node–metastasis; BCLC: Barcelona Clinic Liver Cancer.
Macroscopic or microscopic tumor thrombus of HCC.
Immunohistochemistry
For the paraffin-embedded samples, 5–6-µm-thick sections were deparaffinized, rehydrated, and then placed in a solution of 0.3% hydrogen peroxide for 10 min. Sections were incubated with normal goat serum. Slides were incubated at 4°C overnight with rabbit anti-human Fra-1 antibody (dilution: 1:100; Abcam, USA). Next, the slides were incubated with horseradish peroxidase (HRP)-conjugated secondary antibody (Zhongshan Golden Bridge Biotechnology, China) for 30 min. Then, the slides were incubated with 3,3′-diaminobenzidine, counterstained with hematoxylin, and dehydrated, followed by coverslip mounting. For a negative control, phosphate-buffered saline (PBS) was used instead of the primary antibody.
Scoring was assessed by two professional pathologists. Both the staining intensity and the ratio of positively stained cells were considered. The assessment was based on a previously reported method. 17 Analysis was as follows for staining intensity—no staining: 0 points, weak staining: 1 point, moderate staining: 2 points, and strong staining: 3 points. For the proportion of positive cells, scoring was as follows—≤10% of cells stained: 0 points, 11%–25% of cells stained: 1 point, 26%–50% of cells stained: 2 points, 51%–75% of cells stained: 3 points, and >76% of cells stained: 4 points. The valid score was obtained from the multiplication of density and intensity analysis, which was then classified into two sets for further assessment: negative set (0–2 points) and positive set (3–12 points).
qRT-PCR analysis
RNA extraction was performed using TRIzol reagent (Invitrogen, USA) for the 19 pairs of snap-frozen tumor tissues and adjacent peritumoral tissues, and DNA was synthesized with the Revert Aid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA). qRT-PCR was carried out with the PikoReal Real-Time PCR System (Thermo Fisher Scientific). β-actin was used as the internal control for the detection of complementary DNA (cDNA) tested. The PCR amplification was performed for 40 cycles (95°C for 5 s, 60°C for 30 s, and 72°C for 30 s). The specific primers for human Fra-1 messenger RNA (mRNA) were as follows: 5′-AGCTTTGAGAAGCCTTCAGC-3′ and 5′-CCAAATACTCTCCCATTGGC-3′. The β-actin-specific primers were as follows: 5′-GGGAAATCGTGCGTGACATTAAGG-3′ and 5′-CAGGAAGGAAGGCTGGAAGAGTG-3′. The relative level of Fra-1 mRNA was evaluated by comparing with the β-actin level for each sample with the 2−ΔΔCt method. 18
Western blot analysis
Paired snap-frozen tumor and adjacent peritumoral tissues were lysed with radioimmunoprecipitation assay (RIPA) buffer, and the protein concentrations were measured using the bicinchoninic acid (BCA) protein assay. Extracts containing equal protein amounts were separated on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and continuously transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, USA). The membranes were incubated with the rabbit-anti-human primary Fra-1 antibody (dilution: 1:1000; Abcam) and mouse-anti-human β-actin as a control (Zhongshan Golden Bridge Biotechnology) for overnight at 4°C after blocking with 5% non-fat milk. The membranes were then incubated at 37°C with HRP-conjugated secondary antibody for 2 h. Finally, the blots were visualized with an enhanced chemiluminescence (ECL; Pierce, USA) detection method.
Statistical analysis
SPSS 21.0 (SPSS Inc., USA) was used for statistical analysis. Continuous data were expressed as mean ± standard deviation. Statistical significance was determined with the independent Student’s t test and chi-square test. Survival analysis was performed with the Kaplan–Meier method, and the log-rank test was used to assess the differences of the survival rates. Univariate and multivariate analyses were presented by the Cox proportional hazards model. Significance was set at p < 0.05.
Results
Immunohistochemical analysis of Fra-1 expression
The positive staining was mainly detected in the nuclei of HCC tumor tissues and the cytoplasm of adjacent peritumoral tissues (Figure 1). Fra-1 positive expression was detected in 57.6% (38/66) of tumor tissues and 21.2% (14/66) of peritumoral tissues. Clinicopathological studies indicated that the expression level of Fra-1 was associated with sAFP (p = 0.014) and vascular invasion (p = 0.002). The analysis did not reveal any significant relationship to the other parameters, as detailed in Table 1.

Immunohistochemical staining of Fra-1 in tumor tissues and adjacent peritumoral tissues of HCC. (a1 and a2) High expression of Fra-1 in tumor tissues. (b1 and b2) Negative expression of Fra-1 in tumor tissues. (c1 and c2) Little trace expression of Fra-1 in peritumoral tissues. (d1 and d2) Negative expression of Fra-1 in peritumoral tissues (a1, b1, c1, and d1—original magnification: 200×; a2, b2, c2, and d2—original magnification: 400×).
qRT-PCR analysis of Fra-1 mRNA expression
To quantify the level of Fra-1 mRNA expression, 19 pairs of tumor and adjacent peritumoral samples were analyzed using a semiquantitative qRT-PCR assay. The median expression level of Fra-1 mRNA was significantly higher in tumor tissues than that of adjacent peritumoral tissues (5.45 ± 1.85 vs 1.65 ± 0.30, respectively; p < 0.001), as displayed in Figure 2.

qRT-PCR analysis of Fra-1 mRNA expression in 19 pairs of HCC tissues and adjacent non-cancerous tissues (***p < 0.001).
Western blot analysis of Fra-1 expression
Western blotting was carried out to examine the protein expression level of Fra-1 in the same 19 paired HCC samples. As shown in Figure 3, Fra-1 protein levels in tumor tissues and matched peritumoral tissues were 1.39 ± 0.38 and 0.77 ± 0.29, respectively (p < 0.001), which was consistent with the result of the qRT-PCR analysis.
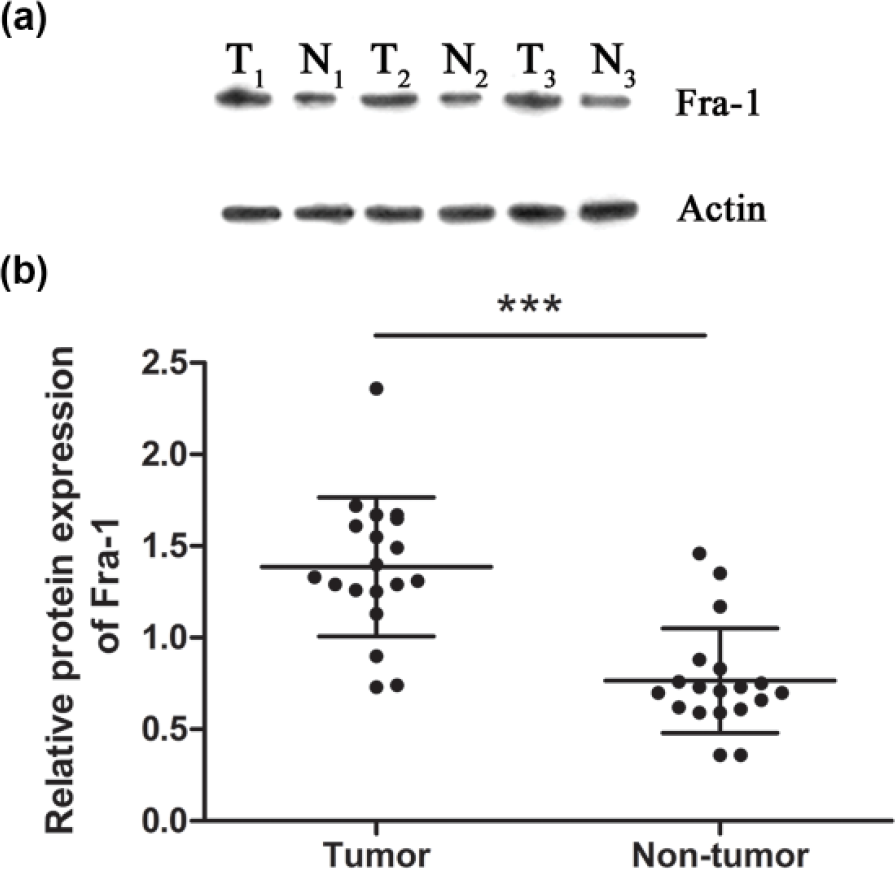
Western blotting analysis of Fra-1 expression in 19 pairs of HCC tissues and adjacent non-cancerous tissues. (a) Fra-1 protein level in the three representative pairs of HCC tissues (T) and adjacent non-cancerous tissues (N). The Fra-1 protein level was normalized to β-actin. (b) The statistical analysis of Fra-1 protein levels in 19 pairs of HCC tissues and adjacent non-cancerous tissues (***p < 0.001).
Clinical significance of Fra-1 in the prognosis of HCC
The Kaplan–Meier survival analysis was used to investigate the association between Fra-1 positive expression and patient survival. Survival curves were compared using the log-rank test. Patients with positive Fra-1 expression had a poorer OS time (25.8 months, 95% confidence interval (CI): 20.5–31.1) than that (42.6 months, 95% CI: 35.3–50.0; p = 0.001) of patients with negative Fra-1 expression. Patients with positive Fra-1 expression also had a poorer DFS time (12.4 months, 95% CI: 7.3–17.6 vs 28.1 months, 95% CI: 19.8–36.4, respectively; p = 0.001) in comparison with patients with negative Fra-1 expression (Figure 4). Univariate analysis showed that vascular invasion, tumor capsule, status of metastasis, TNM stage, BCLC stage, and Fra-1 expression had prognostic influence on OS, while tumor size, vascular invasion, status of metastasis, TNM stage, BCLC stage, and Fra-1 expression were also significantly associated with DFS. Furthermore, multivariate survival analysis showed that only positive Fra-1 expression, capsular invasion, and status of metastasis were confirmed as independent predictors of poor prognosis for both OS and DFS of HCC patients (Tables 2 and 3).

Kaplan–Meier survival analysis of Fra-1 expression in 66 HCC patients after curative resection. (a) Positive expression of Fra-1 was related to worse OS rates compared with the negative expression (p = 0.001). (b) Positive expression of Fra-1 was associated with unfavorable DFS rates compared with the negative expression (p = 0.001).
Univariate Cox regression analysis of patients’ OS and DFS.

OS: overall survival; DFS: disease-free survival; HR: hazard ratio; CI: confidence interval; sAFP: serum alpha-fetoprotein; HbsAg: hepatitis B surface antigen; TNM: tumor–node–metastasis; BCLC: Barcelona Clinic Liver Cancer.
Multivariate Cox regression analysis of patients’ OS and DFS.

OS: overall survival; DFS: disease-free survival; HR: hazard ratio; CI: confidence interval; TNM: tumor–node–metastasis; BCLC: Barcelona Clinic Liver Cancer.
Discussion
In this report, we demonstrated that Fra-1 expression was obviously higher in tumor tissues compared with adjacent peritumoral tissues. Cox regression analysis showed that Fra-1 was an independent prognostic factor.
Fra-1 is upregulated in several types of malignant tumors, including lung, 13 colon, 14 breast, 15 and thyroid 16 cancers. Zhong et al. 13 showed that both mRNA and protein expression levels of Fra-1 were higher in lung cancer tissues compared with the adjacent non-tumor tissues. Similarly, our study validated that in comparison with the adjacent peritumoral tissues, both transcriptional and protein expression of Fra-1 in HCC tissues were upregulated. Neoplastic breast tissues are all nuclear immunoreactive for Fra-1 and benign neoplastic breast tissues are predominantly nuclear immunoreactive, while all breast carcinomas are nuclear and cytoplasmic immunoreactive. 19 In our study, the result that positive expression was mainly detected in the nuclei of HCC tumor tissues and the cytoplasm of adjacent peritumoral tissues was consistent with previous research where immunohistochemical analysis showed that c-Fos, FosB, and Fra-1 were mainly detected in the nuclei in cancer cells and only in the cytoplasm in adjacent non-cancer cells of HCC. 20 Adiseshaiah et al. 21 demonstrated that Fra-1 promoted the formation and metastasis of tumor by A549 cells (human type-II–like alveolar malignant cells) but failed to enhance tumor formation by BEAS-2B (a non-malignant bronchial epithelial cell line) in athymic nude mice, and further showed that Fra-1 could promote motility and invasion of lung epithelial cells in vitro but was insufficient for the formation of tumor. Fra-1 can also inactivate β1-integrin and inhibit the activity of RhoA to promote cell motility and invasiveness of colon carcinoma cells. 22 Ramos-Nino et al. 23 reported that Fra-1 was correlated with levels of CD44 (a cellular adhesion molecule) expression which were higher in simian virus 40 negative malignant mesothelioma (MM) cells, both Fra-1 and CD44 expression was associated with cell migration in MM cells. They confirmed that Fra-1 could modulate the expression of CD44 which might govern cell migration of the selected MMs. Epithelial–mesenchymal transition (EMT) is a fundamental biological process that regulates morphogenesis, cancer invasion, and metastasis in most organisms. 24 Fra-1 can monitor Ha-RAS-induced EMT of human colon cancer cells by regulating the expression of vimentin (a member of the intermediate filament family) in association with the invasion and migration of colon cancer cells. 25 Fra-1 directly induces the activity of the matrix metalloproteinase 1 (MMP-1) and MMP-9 promoter and regulates several genes associated with invasion and angiogenesis in breast cancer progression. 26 A mounting body of evidence suggests that correlations between tumor cells and tumor-associated macrophages (TAMs) in the tumor microenvironment (TME) are pivotal in promoting invasion and metastasis of tumor cells.27,28 The interaction between Fra-1 and the interleukin 6 (IL-6)/Janus kinase (JAK)/signal transducer and activator of transcription (STAT3) signaling pathway regulates the switch from M1 with tumoricidal capacity to M2 with tumor-promoting capacity in breast tumor tissues. 15 The switch plays an important role in the increased release of vascular endothelial growth factor (VEGF) and MMP-9 from these cells, which could promote invasion and progression of tumor cells. Desmet et al. 29 revealed that Fra-1 was a polyphenic regulator of the invasion of breast cancer cells; it contributed to both early (EMT) and later (filopodia formation) events and also promoted the establishment of distant metastases (extravasation and anchorage-independent growth) of breast cancer. Our data suggested that overexpression of Fra-1 was associated with vascular invasion and sAFP of HCC. We suggest that the overexpression of Fra-1 in HCC may upregulate the heterodimer of AP-1 which can bind to the AP-1 site on the promoter of tumor progression–related genes, such as MMPs, CD44, VEGF, and vimentin to stimulate the transcription of tumor-associated proteins. These proteins may promote the vascular invasion of HCC.30–32 Likewise, we speculate that the abnormal expression of Fra-1 promotes the transcription of sAFP 33 and the establishment of EMT 34 and TAM 35 in HCC.
Fra-1 was validated to be a significant prognostic predictor of patient outcome, independent of pathological staging in colorectal cancer 14 and non-small-cell lung cancer (NSCLC). 17 Others have previously shown that Fra-1 regulates high mobility group protein 1 (HMGA1) expression by binding to the promoter region of HMGA1 gene, and a multivariate analysis shows the positive expression of HMGA1 to be an independent prognostic factor. 36 Fra-1 can also upregulate several adhesion-related proteins (CD44 and integrin α5) and increase adhesion of Fra-1-overexpressing MCF7 cells, and the overexpression of Fra-1 contributes to higher rate of lung metastasis and shorter OS in estrogen receptor (ER)-positive breast cancer patients. 37 In our study, survival analysis showed that patients with positive Fra-1 expression had a poor prognosis. Cox regression analysis indicated that Fra-1 was an independent prognostic factor for HCC patients. We speculate that the abnormal expression of Fra-1 in HCC may promote the formation of an invasion phenotype by modulating the mobility and adhesion of tumor cells, resulting in poor prognosis.38,39
Despite the often abnormal expression of Fra-1, the molecular mechanisms which regulate its accumulation in tumor cells remain unclear. Fra-1 accumulation is regulated by the transcription of the fosl1 and post-translational modifications. Transcription of fosl1 is induced by various growth factors and cytokines.40,41 The primary modification of Fra-1 is phosphorylation, which has been validated to affect both its transcriptional activity and stability. A recent study indicated that the degradation of Fra-1 was regulated by two different mechanisms including association with TATA-box-binding protein 1 (TBP-1; 19S proteasomal subunit) and a C-terminal degron which was regulated by RAS–ERK (extracellular signal–regulated kinase) and that TBP-1 may act as a tumor suppressor to reduce Fra-1 levels in tumor cells. 42 Consequently, Fra-1 may be considered as a potential therapeutic target for HCC.
In conclusion, specific expression of Fra-1 was observed in the nuclei of HCC tumor tissues and the cytoplasm of adjacent peritumoral tissues. Its detection with other biomarkers may be used as useful tools in the diagnosis of HCC. It may also be a novel prognostic biomarker and a potential therapeutic target for HCC. The exact mechanisms by which Fra-1 promotes HCC progression remain unclear and deserve further investigation.
Footnotes
Acknowledgements
The authors thank all the experts and donors involved in this research. X.Q.G. wrote the manuscript; Y.S.G. and Q.H.S. revised the paper; and H.X.M. collected the data.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
The study protocol was approved by the Human Research Ethics Committee of Anhui Medical University.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research was supported by the Integrated Projects of Commonwealth Technological Applied Research of Anhui Province (No. 1604f0804011).