
Abstract
PTEN is the second most frequently mutated tumor suppresser gene in cancers after p53. Genetic and epigenetic alterations in the PTEN gene and its regulatory regions have been reported in various studies. PTEN is a crucial downregulator of the pro-survival phosphoinositide 3-kinase/Akt/mammalian target of rapamycin pathway and also suppresses insulin signaling. Failure to regulate these pathways leads to increase in cell proliferation and migration which in turn promotes tumorigenesis. PTEN underexpression is mediated by a variety of cytokines and stress kinases which seem to collectively induce the RAS/mitogen-activated protein kinase (MAPK)/extracellular signal–regulated kinase (ERK) pathway. In the context of hepatocellular carcinoma, reduced expression of PTEN is seen in nearly half of the cases on average. In some cases, PTEN has been observed to be either mutated or methylated which can also lead to reduced expression or in some cases, complete loss of expression. On the cellular level, PTEN is also a target in the pathogenic pathway of hepatitis C virus core protein and hepatitis B virus X protein. These viruses appear to alter PTEN regulation and pro-apoptotic ability to enhance the process of tumor formation. In perspective of the crucial role PTEN plays in balancing proliferation and apoptosis, we propose PTEN as a valuable marker in the diagnosis, assessment of tumor grade, and disease stage in hepatocellular carcinoma patients.
Introduction
Hepatocellular carcinoma (HCC), a malicious tumor of the liver originating from hepatocytes, is the most recurrent type of primary liver cancer. Globally, it is the second primary cause of cancer mortality and the fifth most common type of cancer. 1 The incidence of HCC is not equally distributed throughout the world. Globally, Africa and Asia have higher incidence; owing to China’s large population, it carries about 50% of all cases of HCC. North and South America as well as Europe have low incidence of HCC compared to the Asian region. 2 Over 70% of all new cases of liver cancer are spotted in Asia. 3 This catastrophic cancer is more prevalent in male population compared to females. 4
The biomarkers for HCC diagnosis and staging that are currently in use include alpha-fetoprotein (AFP), des-gamma-carboxy prothrombin (DCP), and lens culinaris agglutinin-reactive AFP (AFP-L3). 5 Compared to AFP-only detection, DCP and AFP-L3 allow detection of 10%–20% more positive cases. 6 Each of these methods has its limitations regarding early diagnosis, sensitivity, and specificity with regard to HCC diagnosis.6,7
A number of tumor suppresser genes and oncogenes are involved in the progression of HCC8,9 which have the potential to be used as biomarkers for early detection of HCC and staging of disease. In this review, we discuss PTEN (phosphatase and tensin homolog deleted on chromosome 10) as a potential biomarker for HCC diagnosis. We focus on expression and regulation of PTEN in the context of HCC. PTEN is a frequently downregulated tumor suppresser gene in cancers and is a crucial mediator of oncogenic pathways.
Etiology of HCC
The major etiological agents responsible for inducing HCC are aflatoxins B1, alcohol, alcoholic and biliary cirrhosis, chronic liver injury, hepatitis C virus (HCV), hepatitis B virus (HBV), and hemochromatosis (Figure 1). 10 Glycogen storage diseases, autoimmune hepatitis (AIH), alpha-1 antitrypsin deficiency, and Wilson’s disease are among the less common risk factors for induction of HCC. 11 The primary means of prevention from virus-induced HCC are vaccination and antiviral treatment. Dietary modifications, weight loss, and tobacco/alcohol cessation remain the secondary means of prevention. Treatment with interferon alpha (IFN-α) has been shown to reduce the risk of virus-induced HCC.12,13 In addition, there are new therapies that are now available for the treatment of HCV. 14 Radiofrequency ablation, tumor resection, and liver transplantation are the major curative options available for HCC. 11

Risk factors for hepatocellular carcinoma. The major etiological agents for HCC are HCV, HBV, hemochromatosis, chronic liver injury, alfatoxin B1, and alcohol and chronic liver disease.
The key contributors of the high occurrence of HCC in Africa and South East Asia are viral infections, that is, HCV and HBV. Nearly 80% of all HCC cases are associated with a HBV or HCV infection. 15 HCV infection is more strongly linked with liver insulin resistance and causes steatosis and fibrosis, 16 and there have been studies that found that 40%–80% of HCC cases have a positive HCV infection as the etiological factor. 17 In total, 3% of the world population is infected with HCV, with total infections estimated to be around 200 million. 18
HCC development that is due to a viral infection is a long process involving a variety of steps that may span over 20–40 years. The five major steps are establishment of chronic viral infection, persistent hepatic inflammation, progressive liver fibrosis, cirrhosis of liver, and finally oncogenesis.19–21 Each stage of HCV-induced hepatocarcinogenesis is a possible target for curative intervention. 19
In case of HBV, there have been studies that report that nearly 50% of all HCC cases are attributable to a chronic HBV infection.22–24 HBV can cause both acute and chronic liver disease. Patients who have acquired HBV have 100-fold more chance of developing chronic liver disease. 25 HBV, a Hepadnaviridae family virus, is a partially double-stranded DNA virus. HBV causes HCC by both direct and indirect pathways. HBV causes hepatocyte injury and chronic necro-inflammation, with fibrosis and cirrhosis. It can also lead to HCC without causing cirrhosis. It is able to integrate its DNA into the host cells and act as a mutagenic agent, thus increasing genomic instability. 22
Pathogenesis of HCC
Like all the other cancers, HCC develops as a result of genetic mutations, leading to an unusually high level of replication at the cellular level.26,27 It causes the cell to avoid the process of apoptosis. In these types of cells, the damaged DNA continues to multiply and increase in number with the mutated DNA copies. As these cancerous cells reproduce, they start to form tumors which may be metastatic or benign. 28
Like all the other types of human cancers, a variety of oncogenic genes are involved during hepatocarcinogenesis.9,29 These genetic changes are associated with tumor suppressor genes, oncogenes, reactivation of developmental pathways, and growth factors and their receptors.30,31 Telomerase activation is also a general feature of HCC and is closely related to immortalization which may be the common result of variations in cancer-related genes as depicted in Figure 2.32,33

Molecular alterations occurring in HCC. Many genes are altered in hepatocellular carcinoma. Some of the commonly altered genes are PTEN, p53, IGF-2, TGF-α, and Myc.
PTEN
PTEN, is one of the most important tumor suppressors in mammals along with p53, Ink4a, and Arf.34,35 It is located on the long arm of chromosome 10 at position 23.3, more specifically from base pair 87,863,438 to base pair 87,971,930. It is located at the 10q23 locus 36 and is sometimes also known as MMAC (mutated in multiple advanced cancers); it was simultaneously discovered by two separate group of researchers almost at the same time.36,37 This locus is highly susceptible to mutations related to human cancers38–41 and also undergoes changes in promoter activity due to methylation which leads to loss of expression or reduced expression in cancer cell lines.41–44 In accordance with its role as a tumor suppressor, PTEN has the ability to induce cell death via apoptosis45,46 and also mediates interactions with extracellular matrix which result in reduced cell migration and inhibition of focal adhesion formation. 47
The gene that encodes PTEN is non-redundant and is expressed in all eukaryotic cells. Various mouse models have been established to study the function of PTEN in vivo. The major role of PTEN is the downregulation of phosphoinositide 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) pathway.46,48 PTEN-mediated suppression of Akt pathway results in increase in longevity, downregulation of insulin signaling, and tumor suppression. 49 PTEN also appears to be essential for embryonic development in mouse models. 50 Some of the PTEN regulators are Ras, early growth response protein 1 (EGR-1), ROS, and nuclear factor kappa B (NF-κB).51,52
Role of PTEN in the regulation of PI3k/Akt pathway
PTEN has its own distinctive growth-regulatory functions based on whether it is in the cytoplasm or nucleus. In the cytoplasm, PTEN has a built-in lipid phosphatase function which is responsible for the negative regulation of the cytoplasmic PI3K/Akt pathway, whereas in the nucleus, PTEN has Akt-independent growth activities. 53
As a dual action lipid and protein phosphatase, function of PTEN in the cells is non-redundant and therefore crucial. Its basic and most important role is in the dephosphorylation of the pro-survival P13K/Akt/mTOR pathway.54,55 PI3K is a lipid kinase responsible for generating phosphatidylinositol (3,4,5)-triphosphate (PIP3 or (PI(3,4,5)P3)) from phosphatidylinositol 4,5-triphosphate (PIP2 or PI(4,5)P2). 56 The next step in this pathway is the recruitment of Akt, also called protein kinase B (PKB), by PIP3 to the cell membrane where it is phosphorylated by phosphoinositide-dependent kinase 1 (PDK1).57,58 Akt undergoes a second phosphorylation step which is catalyzed by mTOR complex 2 (mTORC2). 59 The fully functional Akt can now travel into the nucleus and activate mTORC1 which further triggers a myriad of downstream pathways that are involved in cell growth, angiogenesis, metabolism, and proliferation. 60
PI3K/Akt/mTOR pathway is a complex web which controls its own activation by various fail safe methods. Inhibiting the effects of mTOR via rapamycin is not able to shut down the progression of P3K/Akt pathway alone. In fact, this approach leads to cancellation of a negative feedback loop mediated by Forkhead box protein O1 (FOXO)-induced insulin receptor substrate 1 (IRS-1) that controls PI3K/Akt pathway; when blocked by rapamycin, it leads to uninhibited upregulation of the P13K/Akt pathway.61,62
PTEN has emerged as a promising tumor suppresser gene and key downregulator of this pro-apoptotic pathway. It is reported to be the second most mutated gene in cancers after p53. 54 Studies conducted on PTEN in tumor cell lines have shown that activation of this phosphatase inhibits cell migration and causes cell cycle arrest. PTEN hydrolyzes the PIP3 back to PIP2, thereby inhibiting the action of PIP3 and downstream kinases.46,63 Downregulation of PTEN, especially at advanced stages of HCC, is considered to be the key process that causes dysregulation of the crucial mTORC pathway. 64 Downregulation of PTEN is associated with dephosphorylation of Akt and mTOR and therefore leads to consistent activation of the PI3K/Akt/mTOR pathway which leads to oncogenesis.
Other cellular functions of PTEN
PTEN also plays a role in insulin signaling pathways which has been proved in some cancer models. A study showed that PTEN downregulates the phosphorylation of IRS-1 and blocks the formation of IRS-1/Grb2/Sos complex, thereby reducing insulin stimulated cell growth. 65 In breast cancer cell lines with constitutive expression of PTEN, phosphorylation of extracellular signal–regulated kinase/mitogen-activated protein kinase kinase (ERK/MEK) was inhibited when the cells were exposed to growth factors such as insulin or insulin-like growth factor 1 (IGF-1). Weng et al. 65 therefore suggested that PTEN mediates suppression of insulin signaling by indirectly inhibiting mitogen-activated protein kinase (MAPK) phosphorylation via blocking phosphorylation of IRS-1 and associated receptor complex.
Other than its potent inhibition of PI3K/Akt/mTOR pathway, PTEN has been implicated in the downregulation of separate downstream pathways as well. A study by Vivanco et al. 66 found that in PTEN knockout mice the c-Jun N-terminal kinase (JNK) was consistently upregulated and this was unrelated to the activation of Akt. This study found that JNK was upregulated in a PIP3-dependent manner involving Rac1 and Mlk3. The proposed pathways that mediate the increased expression of JNK are phosphatases, dual specificity protein phosphatase 10 (DUSP10), DUSP4, and DUSP8, that are needed to terminate JNK signaling and were observed to be downregulated in PTEN−/− cell lines. 66
Matrix metalloproteinase 9 (MMP-9), a zinc metalloproteinase involved in the regulation of extracellular matrix structure, has been shown as a downstream target of PTEN in brain tumor models. In glioma cell lines, stable expression of PTEN was able to reduce the levels of hyaluronic acid–induced production of MMP-9 and MMP-2. This led to the possibility of using PTEN phosphatase activity to reduce tumor invasion. 67
Furthermore, it has been shown in a study on adenocarcinoma cell lines that the pro-survival RAS–MAPK signaling brings about PTEN suppression by transforming growth factor beta (TGF-β) signaling. This causes increased cell proliferation through the transcriptional downregulation of PTEN by TGF-β. 68 Other factors that have been implicated in mediating suppression of PTEN via the RAS/RAF/MEK/ERK pathway are c-Jun 69 and binding of NF-κB to PTEN promoter by MEK kinase 4 (MEKK4) and JNK. 70
PTEN is at the nexus of the key pathways which regulate growth, survival, and proliferation. Alteration of these key signaling components play a central role in promoting tumorigenesis. 48 The deregulated expression/activity of PTEN in liver cells contributes to the occurrence of non-alcoholic fatty liver disease (NAFLD) and HCC. Liver-specific PTEN knockout mice spontaneously develop NAFLD and HCC.71,72 Along with other cancers, PTEN is often mutated or biochemically modified and therefore has low expression during HCC.29,73–75 Moreover, dysregulation of PTEN expression/activity strongly promotes uncontrolled cell proliferation and migration in HCC cell lines. 76
PTEN in progression of HCC: HCV core and PTEN
It has been observed that PTEN expression and function is altered in HCV infections which may contribute to the progression of steatosis in case of chronic HCV patients. 77 HCV core protein is thought to be a significant determinant in mediating pathological effects of HCV infection 78 and in creating the pathology of oxidative stress. 79 In a previous study conducted on transgenic mice, HCV core gene was reported to have a major role in development of HCC. HCV core protein can alter tumor suppressive role of retinoblastoma protein (Rb) by downregulating its expression. 80
A study conducted by Zhang et al. 81 reported the downregulation of PTEN by HCV core protein at transcriptional level which resulted in activation of downstream Akt gene. They found that the core protein suppressed PTEN expression by activating the NF-κB pathway which directly interfered with PTEN transcription by interacting with its promoter. The HepG2 cells expressing HCV core protein showed increased growth and survival and had accumulation of cells in G2/M phase. These findings were similar to the PTEN deficient HCC cells 82 signifying a potential role of HCV core in the development of cancer by inhibiting PTEN expression and activating the expression of Akt. The same study also showed that PTEN expression was revived by inhibiting HCV core protein at messenger RNA (mRNA) and protein level. 82
Previously, it has been observed that PTEN causes cell cycle arrest in the G1 phase by reducing the localization of cyclin D1 in the nucleus. Just as the levels of cyclin D1 are reduced in nucleus, the phosphorylation of Rb is also downregulated. In the phosphorylated form Rb is unable to arrest cell cycle progression. 83
HCV core has also been shown to dysregulate PTEN expression by NF-κB activation, and it also increases level of pAkt suggesting a positive feedback loop where upregulation of Akt promotes NF-κB activation. This feedback loop mechanism might enhance the activity of HCV core in tumor growth and expansion. 81 During HCV infection, HCV core protein can downregulate PTEN by activating NF-κB. As PTEN is involved in negative regulation of PI3/Akt pathway which controls cell survival and growth, inhibition of PTEN leads to marked increase in Akt signaling. Akt further activates inhibitor of kappa B (IκΒ) kinase (IKK) which phosphorylates IκB and results in dissociation of NF-κB into the nucleus. 84
Other studies also conform with this model; NF-κB binding to PTEN promoter reduces its expression and induced MAPK signaling thus promoting cell survival. 70 The major pathways that converge in HCV core and PTEN signaling are summarized in Figure 3.

Interplay between HCV core protein and PTEN. (1) HCV core protein suppresses the production of Rb protein. 80 (2) HCV core activates the NF-κB pathway that suppresses PTEN transcription.70,81,82 (3) HCV core indirectly activates pAkt,81,82 (4) which subsequently activates NF-κB pathway. 84 (5) These changes converge in producing an increase in G2/M phase.81,82 (6) PTEN transcription brings G1 arrest which is accompanied by reduced accumulation of cyclin D1 in the nucleus. 83 (7) In normal conditions, Rb protein acts as a tumor suppresser and helps PTEN in causing G1 arrest. 83
PTEN in progression of HCC: HBx and PTEN
Direct involvement of HBx protein has been implicated in the progression of HCC 85 and also in the inhibition of the PTEN expression. HBx-induced overexpression of MMP-9 has been established in the progression of HCC. A study conducted in 2003 found that in HBx-transfected cells, HBx directly disrupted p53-mediated transcription of PTEN. 86 Further studies by the same group found that HBx-transfected cells had increased expression of MMP-9 due to the upregulation of activator protein 1 (AP-1) and NF-κB in these cells. Both these transcription factors are regulated via the ERK1/2 and PI3K pathway, which are strongly involved in increasing cell invasion in HCC. 87
It was observed that in 36 HCC samples, the mRNA levels of PTEN and MMP-9 were inversely correlated. Whereas microarray studies of 200 HCC samples observed that loss of PTEN expression was directly related to tumor grade, metastasis, and vascular invasion. 88
In Huh7 and HepG2 cell lines, overexpression of microRNA 21 (miR-21) was observed when increased expression of HBx protein was induced. MicroRNAs (miRNAs) are endogenously expressed segments of RNA that can have more than one downstream target. With a three to fourfold increase in HBx mRNA expression, a simultaneous 36-fold increase in miR-21 expression was witnessed which also led to increase in cell proliferation. Predictably, this led to a decrease of expression in the miR-21 targets, PTEN, and programmed cell death protein 4 (PDCD4). 88
HBx also mediates the insulin signaling pathway by upregulating the expression of IGF-2 via protein kinase C (PKC) or MAPK signaling. By increasing phosphorylation of transcription factor Sp-1, IGF-2 protein expression is increased by HBx. 89 Furthermore, this is accomplished by the involvement of PKC and MAPK/ERK networks as downregulation of either of these had a direct effect on the phosphorylation of Sp-1. 90 When the role of PTEN in HBx-induced IGF-2 signaling was explored by the same group, it was observed that wherever there was increased expression of PTEN, the levels of IGF-2 were low. 49 This is in line with the findings of Weng et al. 65 that implicated the inhibition of MAPK/ERK (MEK) pathways in inhibition of insulin signaling by PTEN. Kang-Park and Lee 49 also observed reduced DNA binding capacity and phosphorylation of Sp-1, implying that PTEN was inhibiting the PKC and MAPK pathways via its phosphatase activity, thereby limiting tumor progression. The major pathways that converge in HBx and PTEN signaling are summarized in Figure 4.
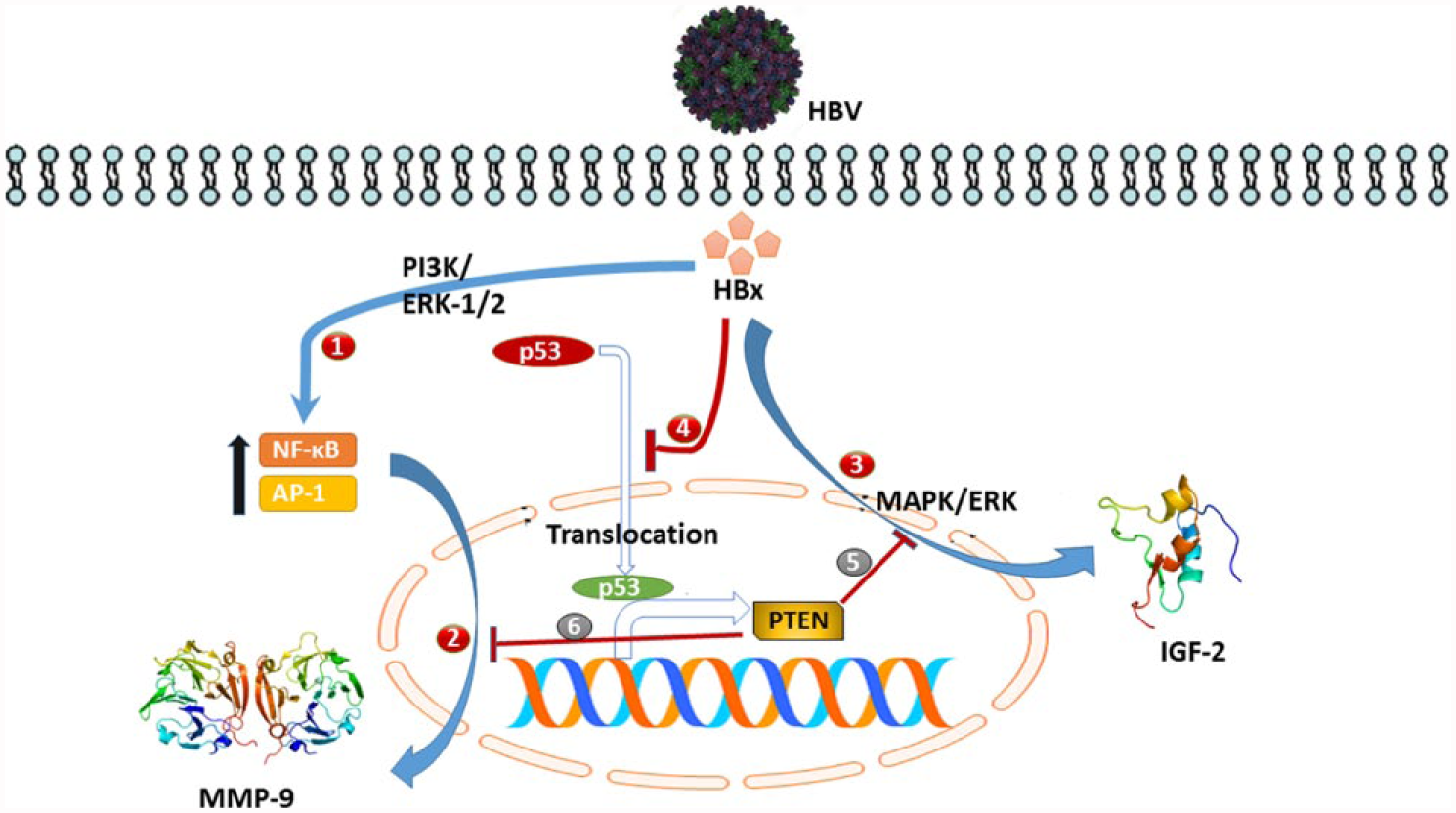
Interplay between HBx protein and PTEN. (1) HBx mediates upregulation of NF-κB and AP-1 transcription factors via the ERK1/2 and PI3K pathways. 87 (2) AP-1 and NF-κB bring about increased expression of MMP-9 protein. 87 (3) HBx upregulates the expression of IGF-2 by mediating MAPK/ERK pathway.89,90 (4) HBx inhibits p53-mediated transcription of PTEN. 86 (5) PTEN brings about inhibition of insulin signaling by downregulating the MAPK signaling.49,65 (6) PTEN indirectly inhibits expression of MMP-9. 88
Role of PTEN as prognostic marker
PTEN knockout mouse models of liver sarcoma have demonstrated that loss of PTEN activity might contribute to HCC. 91 Moreover, about 20%–30% of the patients having HCC have been reported to have allelic loss of PTEN gene.92,93
A few years after the discovery of PTEN, its link with the assessment of disease stage was established in a study by Hu et al. 94 in 2003. They found that lower PTEN expression correlated with stage of disease, tumor grade, and higher expression of AFP, a tumor marker for HCC. 94
A study on patient samples of HCC found that nearly half of the subjects had lower expression of PTEN. This lower expression was also associated with increased tumor size and lower survival rate of patients. 75 Another study conducted in China assessed the expression levels of PTEN in liver tissues and found progressively increasing loss of expression with worsening tumor grade. The study also observed that overall 50% of the HCC samples had no expression of PTEN protein. 95
Comparison between PTEN expression in resected liver tissue and surrounding parenchyma also reveals interesting findings. One study from Japan suggested PTEN as a strong prognostic marker of survival in HCC patients based on their findings. Rahman et al. 96 found that although surrounding normal liver parenchyma had strong expression of PTEN in 90% of the cases, there was low expression of the protein in 65% of the tumorous liver tissue.
Another study has also reported decrease in the protein levels of PTEN gene in more than 40% of HCC specimens which demonstrates the diagnostic significance of reduced expression of PTEN and its value as a prognostic marker for HCC. 97 Loss of PTEN expression was observed in 36% of liver tumors whereas significantly low expression compared to normal tissue was observed in the 200 samples that were under surveillance in this study. 88 This study also found that level of PTEN expression was inversely correlated with the stage of tumor and metastasis. 88
Although several studies show a decline in the transcript levels of PTEN in liver cancer patients, mutations in the PTEN gene are less frequently reported in the setting of HCC.74,98
One study found that in addition to its own downregulation, loss of PTEN expression is also related to increased expression of CD133, CD90, and epithelial cell adhesion molecule (EpCAM), all of which are markers of liver cancer stem cells (LSCSs). The researchers observed that PTEN had a strong prognostic role as a marker for HCC recurrence and survival rate in subjects. Lower expression of PTEN correlated with stronger chances of HCC recurrence; similarly, downregulation of PTEN along with upregulation in expression of CD133 and EpCAM correlated with shorter survival times. 99 The researchers suggested that PTEN along with CD133 and EpCAM could be useful tools to assess HCC prognosis and recurrence.
In 2007, miR-21 was identified as an endogenously expressed miRNA that specifically targeted PTEN. Later on, other miRNAs have also been found that target PTEN, such as miRNA-19a. 100 In malignant cholangiocytes, suppression of miR-21 was able to increase sensitivity to chemotherapeutic agent, gemcitabine. It was found that miR-21 could modulate PTEN-mediated apoptosis in tumor xenografts. 101
Higher expression of this PTEN targeting miRNA has been linked to the presentation of poorly differentiated hepatocellular tumors. 102 In another study, significantly higher expression of miR-21 was observed in malignant HCC tumors 103 and has been linked to the promotion of tumorigenesis in liver cancer cases. 104
So, it can be suggested in light of these recent studies that HCC pathogenesis is significantly associated with the reduced expression of PTEN gene, which can be modulated by either the increased expression of miRNAs or by the upregulation of PTEN inhibiting signaling pathways. Therefore, level of PTEN expression can be exploited to assess the stage of HCC and to establish a prognosis for the patients.
Conclusion
Since its discovery in 1997, PTEN has emerged as a significant and non-redundant tumor suppresser gene. Specifically in case of HCC, the loss or underexpression of PTEN is reported in over half of the study sample that has been summarized in this review. Although mutations in the PTEN gene are less frequently reported in HCC compared to other cancers, its role in assessment of tumor grade, disease stage, and patient survival has been established by many researchers. It is apparent that dysregulation of PTEN plays an important pathogenic role in the progression of HCC. It will be of interest to explore the sensitivity of PTEN as a tumor marker and compare with other available biomarkers. Further research is warranted in order to understand the regulation of PTEN expression and its downstream signaling, which will in turn help in designing better treatment strategies for HCC and other cancers.
Footnotes
Acknowledgements
A.K. and T.H. contributed equally to this work. Conception and design, acquisition of data, analysis and interpretation of data, and article drafting was done by A.K. and T.H. S.M. was the principal investigator and was involved in conception and design, analysis and interpretation of data, critical revision of the article, and final approval of the version to be published. M.S. analyzed and interpreted the data and drafted the article. Critical evaluation of the article was done by S.K.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.