
Abstract
Osteosarcoma is one of the most lethal malignancies, and the prognosis remains dismal due to the paucity of effective therapeutic targets. Bmi-1 and TRIM-14 are associated with the initiation and progression of osteosarcoma, which could promote angiogenesis, invasion, and apoptotic resistance in bone cancer tissue. In this study, we constructed a bispecific antibody of BsAbBmi/TRIM targeting Bmi-1 and TRIM-14 and investigated the therapeutic value in bone carcinoma cells and xenograft mice. Our results showed that Bmi-1 and TRIM-14 expression levels were markedly upregulated correlated with nuclear factor-κB nuclear translocation in bone cancer cells and clinical carcinoma tissues. Results have demonstrated that overexpression of Bmi-1 and TRIM-14 promoted growth, proliferation, aggressiveness, and apoptosis resistance of osteosarcoma cells. BsAbBmi/TRIM administration significantly inhibited nuclear factor-κB expression derived by matrix metalloproteinase–9 promoter. BsAbBmi/TRIM administration inhibited growth of osteosarcoma cells and downregulated Bmi-1 and TRIM-14 expression levels. Data also demonstrated that migration and invasion of osteosarcoma cells were also inhibited by BsAbBmi/TRIM. In addition, results illustrated that BsAbBmi/TRIM inhibited tumor growth and tumorigenicity by blockaded sensor expression in nuclear factor-κB signal pathway. Furthermore, in vivo study showed that BsAbBmi/TRIM treatment markedly inhibited the tumorigenicity and growth of osteosarcoma cells compared to either AbBmi-1 or AbTRIM-14 treatment. Notably, survival of xenograft mice was prolonged by BsAbBmi/TRIM treatment compared to either AbBmi-1 or AbTRIM-14 treatment. In conclusion, these results provided new evidence that BsAbBmi/TRIM inhibited the progression of osteosarcoma, which suggest that BsAbBmi/TRIM may be a novel anti-cancer agent for osteosarcoma therapy.
Introduction
Bone cancer occurs in skeleton, and its affiliates and tumorigenesis are still remained unclear. 1 Malignant bone cancer is a typical systemic malignant disease, and the common symptoms are bone pain, swelling, and fatigue in clinical patients.2,3 In recent years, more and more strategies have been proposed; however, the overall survival rate of osteosarcoma patients required little improvement.4,5 In addition, apoptosis resistance for osteosarcoma cells contributes to the difficulty for cancer patients.6–8 Furthermore, researches have revealed that apoptotic resistance has become the greatest challenge in cancer therapy due to fierce resistance of tumor cells through many kinds of metabolisms.9,10 Therefore, exploring more efficient target therapies is required to improve apoptosis resistance for osteosarcoma therapy.
B-cell-specific Moloney murine leukemia virus integration site 1 protein (Bmi-1) plays an essential role in tumor growth via forming complexes with multiple other polycomb group (PcG) family members. Bmi-1 expression is associated with dysplastic cell transformation during carcinogenesis, which is required for cancer cell replication and survival. 11 Bmi-1 is also identified as transcription factor that presents prognostic roles in several malignancies.12,13 Previous study has indicated that Bmi-1 acts as an oncogene and plays important role in the development and progression of cancer. 14 Additionally, research has found that overexpression of Bmi-1 could promote apoptosis resistance of cancer cells via activating nuclear factor-κB (NF-κB) signaling pathway. 15 In contrast, Bmi-1 knockdown can induce cell-cycle arrest and upregulate the tumor-inhibiting gene expression including HOXC13, p16INK4a, and HOXA9. 16
The superfamily of tripartite motif-containing (TRIM) protein family includes more than 60 kinds of TRIM protein that is evolutionarily conserved with the most striking feature of the highly conserved order of domains in the RING–B-box–coiled-coil (RBCC) motif. 17 Tripartite motif containing-14 (TRIM-14) is an important member of TRIM family protein, which could promote proliferation, angiogenesis, growth, invasion, and increased apoptosis resistance in carcinoma cells. 18 In addition, TRIM-14 has been found to have higher expression in monkey lymphomas caused by HIV and simian immune deficiency (SIV). 19 Furthermore, gene expression of TRIM-14 also plays a mediator role in immune responses and gene transcription by regulating NF-κB signaling pathway. 18 Therefore, these reports suggest that TRIM-14 upregulation may be associated with tumorigenesis in the progression of human cancer.
Aberrant NF-κB activation is observed in majority of human cancer.20,21 Evidences have suggested that poor survival rate and insufficient outcomes of bone cancer are associated with aberrant expression of NF-κB signal pathway.22,23 The indicator of NF-κB activation p65 has also been demonstrated as extremely active in clinical specimens of bone cancer. 23 Aberrant expression of NF-κB involves in apoptosis resistance induced by chemotherapy and enhances tumors cell survival, proliferation, survival, invasion, and angiogenesis. 24 Therefore, discovering novel molecules that regulate aberrant activation of NF-κB may be beneficial for the treatment of bone cancer.
In this study, a bispecific antibody of BsAbBmi/TRIM targeting Bim-1 and TRIM-14 was constructed and its potential in cancer therapy was investigated. We found that BsAbBmi/TRIM inhibited the migration and invasion of osteosarcoma cells. Our data demonstrated that BsAbBmi/TRIM inhibiting the invasive phenotype was mechanistically associated with inactivation of NF-κB, which subsequently inhibited expression and activity of MMP-9 in osteosarcoma cells. Our findings provided new evidences that BsAbBmi/TRIM may be a novel anti-cancer agent for bone cancer therapy.
Materials and methods
Animal experiments
Specific pathogen-free (SPF) BALB/c nude (6–8 weeks old) mice were purchased from Harbin Veterinary Research Institute (Harbin, China). All animals were fed under pathogen-free conditions. A volume of 1 × 106 U-2OS cells were injected subcutaneously into dorsum of BALB/c nude mice in a total volume of 200 µL. The detailed procedures were carried out according to previous report. 25 Mice were received treatments of phosphate-buffered saline (PBS), AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM (10 mg/kg). The treatments were continued five times at intervals of every 2 days. Tumor diameters were recorded once every 2 days, and tumor volumes were calculated using the following formula: 0.52 × smallest diameter 2 × largest diameter.
Statistical analysis
Statistical tests for data analysis included Fisher’s exact test, log-rank test, chi-square test, and Student’s two-tailed t-test. Multivariate statistical analysis was performed using a Cox regression model. Statistical analyses were performed using the SPSS 19.0. Data represent mean and standard error of mean (SEM); p < 0.05 was considered statistically significant.
Results
Bmi-1 and TRIM-14 induced the aggressive phenotype in osteosarcoma cells in vitro
In order to investigate the effect of Bmi-1 and TRIM-14 on the aggressiveness of osteosarcoma cells, bone carcinoma U-2OS cells stably overexpressing Bmi-1 or TRIM-14 were established (Figure 1(a)). The data in Figure 1(b) showed that ectopic expression of Bmi-1 and TRIM-14 significantly elevated the migration of osteosarcoma cells determined by wound-healing assays. Migration and invasion assays revealed that Bmi-1 and TRIM-14 overexpression markedly increased migration of U-2OS cells compared to control (Figure 1(c) and (d)). Notably, we found that overexpression of Bmi-1 and TRIM-14 displayed cellular morphologies typical of a highly invasive phenotype and presented increased numbers of outward projections analyzed by the three-dimensional (3D) spheroid invasion assay (Figure 1(e)). In addition, our results in Figure 1(f) demonstrated that Bmi-1 and TRIM-14 expression levels were higher in patients with bone carcinoma. In addition, furthermore, the data in Figure 1(g) showed that Bmi-1 and TRIM-14 hematoxylin–eosin (H&E) staining for primary bone carcinoma from patients with stages I–IV were significantly higher than noncancerous tissues analyzed by quantitative analysis (p < 0.01). Moreover, univariate and multivariate analyses indicated that Bmi-1 and TRIM-14 expression was an independent prognostic factor in osteosarcoma (Supplementary Table 2). Importantly, we analyzed the relationship between Bmi-1 and TRIM-14 expression and survival of bone carcinoma patients. Our data presented that lower Bmi-1 and TRIM-14 expression level had predicted longer survival for patients with osteosarcoma (p < 0.01, Figure 1(h)). We also observed that BsAbBmi/TRIM could enter into the intracellular of U-2OS cells determined by immunofluorescence (Supplementary Figure S1). Collectively, our findings suggest that overexpression of Bmi-1 and TRIM-14 promoted the migration and invasiveness of osteosarcoma cells in vitro and presented potential association between Bmi-1 and TRIM-14 upregulation and osteosarcoma progression.

Bmi-1 and TRIM-14 expression in bone cancer cells and clinical tissues. (a) Expression levels of Bmi-1 and TRIM-14 in bone cancer cells. (b) Wound-healing assay analyzed the function of Bmi-1 and TRIM-14 for bone carcinoma U-2OS cells. (c) Representative micrographs and quantification of cell migration in the Transwell migration assay. (d) Representative micrographs and quantification of cell invasion in the Transwell matrix penetration assay. (e) Representative micrographs from the three-dimensional spheroid invasion assay on the fourth day after cells were recorded. (f) Statistically quantified between normal bone tissues and clinical stages I–IV bone cancer analyzed by average MOD of Bmi-1 and TRIM-14 staining. (g) Bmi-1 and TRIM-14 protein expression levels in human clinical stage I–IV bone carcinoma tissues compared to normal tissues determined by IHC staining. (h) Survival rate of bone cancer patients suffered from low and high Bmi-1 and TRIM-14 expression levels analyzed by Kaplan–Meier survival curves. Error bars represent the mean and SEM of three independent experiments (**p < 0.01).
Bmi-1 and TRIM-14 induced expression of MMP-9 through activation of the NF-κB pathway
To understand the mechanism of overexpression of Bmi-1 and TRIM-14 which promoted the migration and invasion of osteosarcoma cells, we investigated the expression and activity of MMP-9 in U-2OS cells stably overexpressing Bmi-1 and TRIM-14. The results in Figure 2(a) showed that MMP-9 messenger RNA (mRNA) expression was upregulated in Bmi-1 and TRIM-14-overexpressing cells compared to that in control cells. We confirmed that overexpression of Bmi-1 increased MMP-9 activity in osteosarcoma cells (Figure 2(b)). To establish cell model in which endogenous Bmi-1 and TRIM-14 was neutralized, AbBmi-1, AbTRIM-14 and BsAbBmi/TRIM were used to analyze the efficacy of target therapy. Western blotting showed that Bmi-1 and TRIM-14 protein expression was downregulated in osteosarcoma cells after treatment of AbBmi-1, AbTRIM-14 and BsAbBmi/TRIM (Figure 2(c)).

Downregulation of Bmi-1 and TRIM-14 expression level inhibited tumor cells activities. (a) RT-qPCR analyzed MMP-9 mRNA expression levels in Bmi-1 and TRIM-14-overexpressing cells. (b) ELISA for secreted MMP-9 protein expression in U-2OS cell supernatants. (c) Bmi-1 and TRIM-14 expression was inhibited by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM, respectively. (d) Analysis of the efficacy of AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM for U-2OS cells determined by wound-healing assay. (e) Migration of U-2OS cells was inhibited by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM. (f) Invasion of U-2OS cells was inhibited by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM. (g) MMP-9 mRNA expression level was suppressed by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM. (h) MMP-9 protein activity was inhibited by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM. Error bars represent the mean and SEM of three independent experiments (**p < 0.01).
In addition, neutralizing Bmi-1 and TRIM-14 expression dramatically reduced the aggressiveness and spheroid morphology (Figure 2(d)–(f)). Furthermore, neutralizing Bmi-1 and TRIM-14 expression markedly downregulated MMP-9 expression and activity in osteosarcoma compared to control cells (Figure 2(g) and (h)). We performed quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis to quantify the expression levels of selected NF-κB target genes, including CCND1, BcL-XL, tumor necrosis factor (TNF)-α, vascular endothelial growth factor (VEGF)-C, and MYC, in Bmi-1 and TRIM-14-overexpressing, neutralizing osteosarcoma cells (Supplementary Figure S2). Data indicated that AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM inhibited normal cells and convert to cancerous cells by decreasing expression levels of Bmi-1 and/or TRIM-14 (Supplementary Figure S3). Collectively, experimental data indicated that neutralizing Bmi-1 and TRIM-14 expression promoted the transactivation activity of the NF-κB binding site present in the MMP-9 promoter and inhibited the aggressiveness of osteosarcoma cells.
Bmi-1 and TRIM-14 induced aggressiveness in bone carcinoma cells through the NF-κB/MMP-9 pathway
Next, we examined whether Bmi-1 and TRIM-14 increased the aggressiveness of osteosarcoma cells via activation of the NF-κB signaling pathway. The increased migratory and invasive ability of Bmi-1-overexpressing osteosarcoma cells was dramatically reversed by treatment with NF-κB inhibitor (JSH-23), and these effects were accompanied by a reduction in MMP-9 expression and activity (Figure 3(a)–(d)). We found that expression of AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM in osteosarcoma cells markedly decreased the growth rate and augmented the anchorage-independent growth ability in these cells (Figure 3(e) and (f)). Meanwhile, the ability of osteosarcoma cells to induce human umbilical vein endothelial cells (HUVEC) tube formation and chorioallantoic membrane (CAM) neovascularization and invasive ability of osteosarcoma cells were significantly decreased in the osteosarcoma cells that were neutralized by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM (Figure 3(g) and (h)). Furthermore, AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM treatment inhibited the resistance of osteosarcoma cells to apoptosis induced by treatment with the chemotherapeutic agent cisplatin (Figure 3(i)). Western blotting assays presented that the levels of p65, p84, IKK-β, and IκBα were obviously decreased in Bmi-1 and TRIM-14 overexpressing osteosarcoma cells, while the levels were downregulated after treatment with AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM (Figure 3(j)). Furthermore, BsAbBmi/TRIM treatment canceled the stimulatory effect of Bmi-1 and TRIM-14 on NF-κB phosphorylation activation and markedly canceled the enhanced effects of Bmi-1 and TRIM-14 on bone carcinoma cell aggressiveness (Figure 3(k) and (l)). Collectively, these results indicated that the BsAbBmi/TRIM inhibited aggressive phenotype in osteosarcoma cells, leading to the downregulation of the NF-κB target gene MMP-9 through activation of the NF-κB signaling pathway.

Bmi-1 and TRIM-14 regulated bone tumor cell growth through the NF-κB signaling pathway. (a) U-2OS cell migration in the Transwell assay in cells treated by NF-κB inhibitor or MMP-9 inhibitor. (b) Quantification of U-2OS cell invasion in the Transwell assay in cells treated by NF-κB inhibitor or MMP-9 inhibitor. (c) Representative micrographs of the 3D spheroid invasion assay analyzed the efficacy of NF-κB inhibitor or MMP-9 inhibitor. (d) ELISA analysis of MMP-9 secretion after treatment with NF-κB inhibitor or MMP-9 inhibitor. (e) AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM inhibited U-2OS cell growth. (f) Representative micrographs of BrdU-positive signaling in the cells treated with AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM. (g) Representative images and quantification of HUVECs cultured on Matrigel-coated plates with conditioned medium in AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM and control cells. (h) Representative images of the CAM blood vessels stimulated with conditioned medium from indicated cells. (i) Apoptosis resistance of bone cancer cells induced by cisplatin after treatment with indicated agents analyzed flow cytometry. (j) Western blotting analysis of p65, p84, IKK-β, and IκBα expression levels in bone cancer cells treated by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM. (k) Analysis of phosphorylation level of NF-κB in bone cancer cells after treatment with indicated agents for 24 h. (l) AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM canceled the enhanced effects of Bmi-1 and TRIM-14 on bone carcinoma cell aggressiveness. Error bars represent SD of three independent experiments (**p < 0.01).
Targeting of Bim-1 and TRIM-14 expression showed benefits for bone cancer therapy in vivo
Immunohistochemistry (IHC) analysis of 127 cases of clinical human osteosarcoma specimens revealed that upregulation of Bmi-1 and TRIM-14 was associated with upregulation of MMP-9 and nuclear localization of NF-κB (Supplementary Figure S4). To further evaluate the effects of Bmi-1 and TRIM-14 on bone carcinoma cell growth, the biological effects of AbBmi-1, AbTRIM-14 and BsAbBmi/TRIM on bone carcinoma growth and metastasis were further analyzed by establishing osteosarcoma xenograft mice tumor model. As shown in Figure 4(a)–(c), tumors formed by AbBmi-1, AbTRIM-14 and BsAbBmi/TRIM-treated cells exhibited a smaller size and mass than tumors formed by the control cells. The results in Figure 4(d) revealed that AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM decreased percentages of Ki67-positive cells, greater microvascular density (MVD), and fewer TUNEL-positive cells, whereas PBS-treated tumors displayed higher Ki67 proliferation index and MVD and a higher percentage of TUNEL-positive apoptotic cells. Notably, our results also exhibited that NF-κB luciferase activity was upregulated by Bmi-1 and TRIM-14 and downregulated by AbBmi-1, AbTRIM-14, and BsAbBmi/TRIM (Supplementary Figure S5). In addition, we also showed that the N-ethylmaleimide sensitive factor (NSF) was significantly decreased after BsAbBmi/TRIM treatment compared to control groups (Figure 4(e)). Furthermore, BsAbBmi/TRIM treatment significantly elevated paw withdrawal mechanical threshold (PWMT), while BsAbBmi/TRIM-treated mice exhibited a decreasing PWMT compared to PBS group ( Figure 4(f)). Moreover, the NF-κB-activating effect of Bmi-1 and TRIM-14 was examined in osteosarcoma clinical tissues. Data also suggested that activation of NF-κB was essential for the highly invasive phenotype induced by Bmi-1 and TRIM-14 in osteosarcoma tumors. Moreover, experimental data showed that neither overexpressing nor neutralizing Bmi-1 and TRIM-14 had any an effect on the mRNA and protein levels of MMP-2 or MMP-7 (Supplementary Figure S6). Moreover, we observed that MMP-9 and NF-κB expression were downregulated after treatment with target-therapy agents (Figure 4(g)). Importantly, BsAbBmi/TRIM treatment significantly prolonged survival of xenograft mice (Figure 4(f)). Collectively, results further supported the notions that Bmi-1 and TRIM-14 induced MMP-9 expression, which suggested that BsAbBmi/TRIM inhibited osteosarcoma progression, involved in relaxation of bone cancer pain, and may be a potential anti-cancer agent.
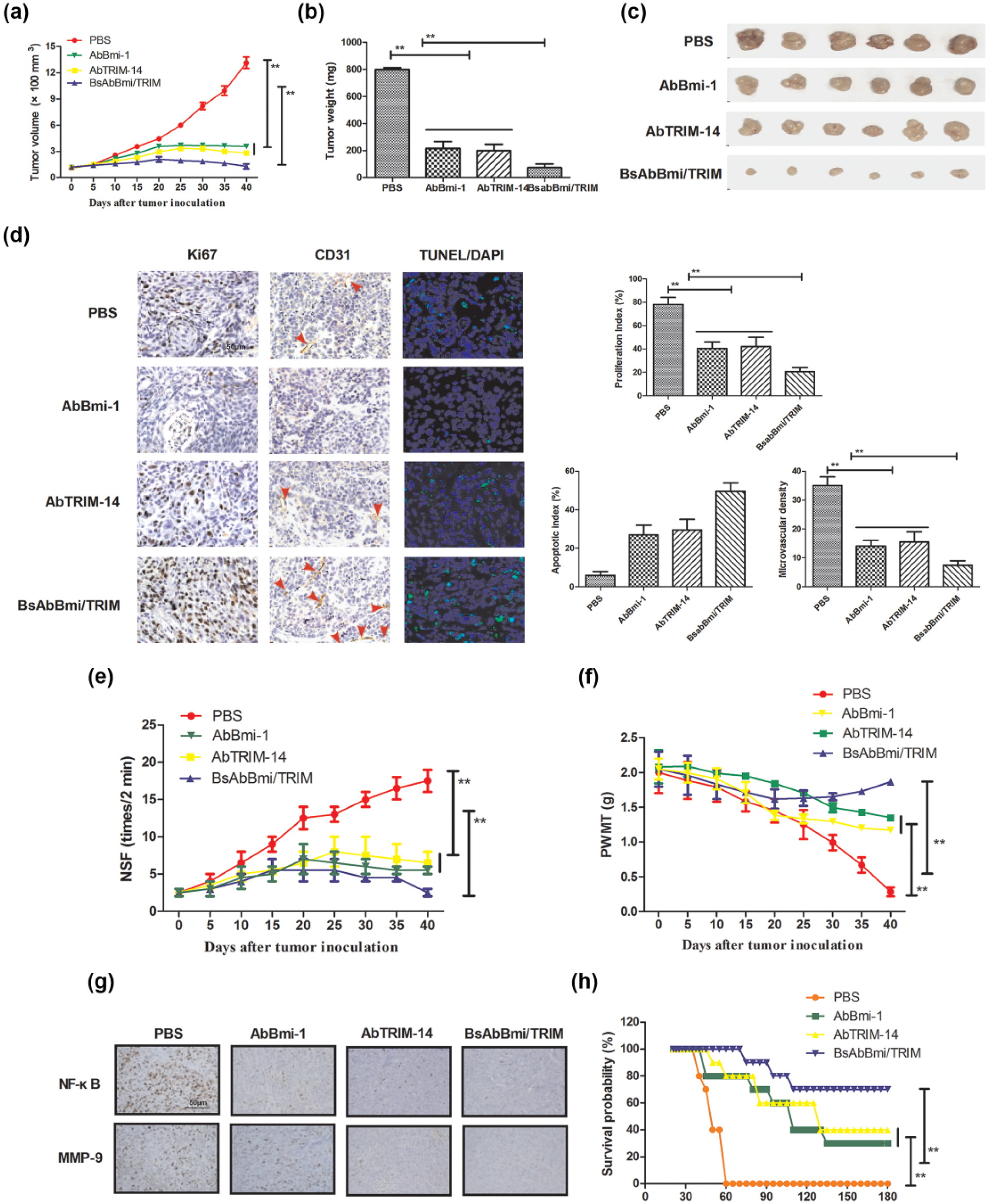
Inhibition of Bmi-1 and TRIM-14 expression contributes to bone carcinoma therapy in vivo. (a) Mean of tumor volumes was measured after treatment with indicated agents. (b) Representative images of tumors from experimental mice in each group. (c) Mean of tumor weight after treatment with indicated agents. (d) Expression levels of Ki67, CD31, and TUNEL-positive cells in tumors treatment with indicated agents determined by IHC staining. (e and f) Pain-related behaviors of tumor-bearing mice after treatment with indicated agents determined by (e) NSF and (g) PWMT. (g) MMP-9 and NF-κB expressions were downregulated after treatment with indicated agents. (h) Survival of xenograft mice was prolonged after treatment with indicated agents. Error bars represent SD of three independent experiments (**p < 0.01).
Discussion
The purpose of this study was to investigate the anti-tumor efficacy of BsAbBmi/TRIM targeting of Bmi-1 and TRIM-14 for bone carcinoma therapy. Previous reports have reported that Bmi-1 and TRIM-14 could induce an aggressive phenotype in cancer progression through modulation of the NF-κB signal pathway.14,26 Therefore, understanding the role of Bmi-1 and TRIM-14 is essential to tumor treatments in human tumorigenesis and metastasis. In this study, our data have identified that Bmi-1 and TRIM-14 promoted migration and invasion of osteosarcoma cells through activating NF-κB signaling pathway. We also identified that overexpression of Bmi-1 and TRIM-14 promoted tumorigenicity and apoptosis resistance through downregulation of NF-κB signal pathway. Importantly, BsAbBmi/TRIM-treated tumors were significantly smaller and had lower tumor weights than control tumors in xenograft mice. These data elaborated that Bmi-1 and TRIM-14 may serve as potential molecular targets and BsAbBmi/TRIM will be a potential anti-cancer agent through inhibition of NF-κB signaling pathway for osteosarcoma therapy.
Apoptosis resistance is the most serious obstacle in cancer clinical treatments.27,28 Decreasing apoptosis resistance of cancer cells contributed to eradication of the residual cells in patients who undergone chemotherapy followed by tumorectomy. Currently, TRIM-14 is reported as an oncogene and can promote tumor growth, aggressiveness, and tumor angiogenesis, while knockdown of its expression inhibited tumor growth, migration, invasion, and tumor angiogenesis in human colorectal cancer cells. 29 Additionally, TRIM-14 overexpression also can predict poor survival in colorectal cancer patients. 30 In this study, we have showed that TRIM-14 could regulate tumor proliferation and tumorigenicity of bone carcinoma, whereas BsAbBmi/TRIM resulted in anti-cancer efficacy through inhibiting NF-κB signal pathway.
Bmi-1 is a member of the PcG family and frequently overexpressed in majority of human tumor cells, which have identified as a potential oncogene in the initiation of tumorigenesis. 31 In addition, Bmi-1 could suppress its targets (p14ARF and p16INK4a) and lead to tumor migration by suppressing the p14ARF/MDM2/p53 and/or p16/Rb signaling pathways. 32 Furthermore, Bmi-1 upregulation could enhance the aggressiveness of human carcinoma and regulate the epithelial–mesenchymal transition (EMT) through modulation of the phosphoinositide 3-kinase (PI3K)/Akt/GSK-3 β pathway. Our results showed that BsAbBmi/TRIM suppressed growth and aggressiveness of osteosarcoma cells by inhibiting Bmi-1 expression, suggesting that Bmi-1 may be a potential therapeutic target for osteosarcoma therapy.
Interestingly, MMP-9 transcription involves in aggressiveness of osteosarcoma in different signaling pathways. 33 MMP-2 may be involved in adriamycin-induced resistance dependent on ERK1/2 signal pathway, which may be a new method of targeted therapy for human osteosarcoma treatment. 34 The correlation between MMP-2 and NF-κB has been systematically researched in esophageal squamous cell carcinoma. 35 Also, NF-κB transcription factors could induce expression and activation of MMP-9 by interacting with binding sites and consequently promote tumor endothelial cell growth. This study indicated that BsAbBmi/TRIM significantly inhibited growth and invasion of osteosarcoma by blocking the activity of NF-κB, which was attributed to inhibition of expression levels of Bmi-1 and TRIM-14.
In conclusion, this study provided evidences of Bmi-1 and TRIM-14 overexpression in bone cancer cells and clinical bone cancer tissues. Inactivation of NF-κB signaling pathway through BsAbBmi/TRIM could inhibit growth, aggressiveness, angiogenesis, and apoptosis resistance in bone cancer in xenograft mice. These findings indicate that BsAbBmi/TRIM may serve as a promising anti-cancer agent for the treatment of osteosarcoma.
Footnotes
Acknowledgements
A.L. and A.T. have contributed equally to this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.