
Abstract
Colorectal cancer is the third common cancer after lung and genital cancers worldwide with more than 1.2 million new cases diagnosed annually. Although extensive progress has been made in the treatment of colorectal cancer, finding novel targets for early diagnosis and effective treatment of these patients is an urgent need. Nuclear factor-erythroid 2–kelch-like ECH-associated protein 1 signaling pathway plays a key role in protecting cells from the damage of intracellular oxidative stress and extracellular oxidizing agents. Nuclear factor-erythroid 2 is a transcription factor that creates intracellular redox homeostasis via transcriptional activity and interaction with kelch-like ECH-associated protein 1. Furthermore, it contributes to survival and chemoresistance of colorectal cancer cells which is mediated by overexpression of cytoprotective and multidrug resistance genes. In this review, the dual role of nuclear factor-erythroid 2 signaling in induction of colorectal cancer cell survival and death as well as the possibility of targeting nuclear factor-erythroid 2–kelch-like ECH-associated protein 1 axis as an advanced strategy in prevention and effective treatment of colorectal cancer patients have been discussed.
Introduction
Each year, more than 1.2 million new cases are diagnosed with colorectal cancer (CRC) worldwide. Our improved knowledge of cancer biology in CRC accompanied by a number of novel proteins and development of genomic-based technologies has shown that CRC can be considered as a heterogeneous disease.1,2 Thus, there is an urgent need to apply molecular-based therapeutic approaches as well as combinations of targeted therapies and chemotherapy in CRC treatment. 3 Diagnosis and staging of CRC are still based on histological analysis of the biopsy specimen obtained from the tumor tissues. The most generally applied prognostic factors to evaluate recurrence risk and survival rate of the patients are T stage (tumor size) and N stage (lymph node involvement). Postoperative adjuvant chemotherapy is suggested for patients with stage-III colon cancer. However, the responses to this treatment are widely variable among stage-III CRC patients. Several clinical and pathological factors are measured in stage-II CRC patients, including number of lymph nodes sampled, evidence of obstacle and/or perforation, histological grade, and lymphovascular and perineural invasion. 4 CRC is the third most common cancer after lung and genital cancers 5 with various survival rate including 90% of 5-year survival for patients with localized stage of cancer and 10% for those diagnosed with a distant metastatic cancer. The risk of CRC in the general population is about 5% which increases gradually with age. 6 The general knowledge of cancer prevention and treatment is improving. Recent studies have indicated that chronic inflammation plays an important role in cancer development. About 15%–20% of cancer-related mortality worldwide, including CRC, is associated with infection and inflammation. 7 Therefore, the mechanism of CRC formation, prevention, and treatment modalities are important. Based on the previous studies, nuclear factor-erythroid 2 (Nrf2) plays a critical role in tumorigenesis and cancer progression via regulating the expression of downstream genes involved in chemoresistance. The aim of this review is to define the impact of Nrf2-dependent signaling in initiation and progression of CRC. The potential possibility of considering Nrf2-antioxidant response elements (AREs) signaling pathway as an advanced target for treatment of inflammatory diseases of the colorectal and prevention of its spreading toward malignancy has also been discussed.8–11
Epidemiology of CRC
Accumulation of both genetic and epigenetic deregulation results in genome alterations. 12 Both of the mutations and epigenetic modifications have been initially considered as two separate mechanisms contributing to carcinogenesis. However, current evidence has revealed interference between gene mutations and epigenetic variations in cancer formation. Therefore, it has been suggested that gene mutations lead to distribution of several epigenetic patterns as well as mutagenesis and genome instability which can be mediated by epigenetic modifications.13,14 For instance, whole exon sequencing of thousands of human cancer samples has shown unanticipated mutations in genes involved in epigenetic mechanisms which have the potential to disrupt DNA methylation patterns, histone modifications, and nucleosome positioning 14 One of the most investigated epigenetic marks in CRC is DNA methylation.15,16 Methylation of CpG islands in the promoter region of a gene can induce modification of chromatin and impedes the availability of the transcriptional device which consequently results in alteration of gene expression. Promoter hypermethylation and demethylation are usually related to gene silencing and gene expression, respectively. Given the fact that numerous genes undergo epigenetic changes in cancer, the role of epigenetic alterations and DNA methylation should be taken into consideration for future approaches of cancer diagnosis, prognosis, and anticipation of therapies. 17 In 1983, Feinberg and Vogelstein 18 have reported the DNA hypomethylation and depletion of overall 5-methylcytosine add-up in CRC tissues, which fundamentally observed at CpG dinucleotides in repetitive sequences are age-dependant and associated with early stages of carcinogenesis. It has also been shown that global DNA hypomethylation, hypermethylation, and transcriptional silencing of tumor-suppressor or expressive genes widely affect DNA repair proteins in CRC tissues. 19 Previous studies have shown that gene methylation occurs in a multi-step process that triggers the conversion of normal colonic epithelium to adenocarcinoma. It has been observed that some of the hypermethylated genes are involved in the transmission of normal colon epithelium to aberrant crypt foci. Furthermore, other methylated genes are participated in the transmission of an aberrant crypt focus to polyp/adenoma or CRC tumorigenesis. Some studies on CRC diagnosis introduced methylation analysis of several hypermethylated genes from plasma or fecal samples, including MutL homolog 1 (MLH1), adenomatous polyposis coli (APC), and Septin9 as valuable non-invasive biomarkers.20,21 Methylation of important genes, such as those involved in the oxidative stress hemostatic pathway, is associated with low survival rates for CRC patients. Therefore, epigenetic biomarkers have prognostic value for CRC (Figure 1). 22

The procedures of constant accumulation of Nrf2 in nucleus of colorectal cancer cells. The effect of Keap1-Nrf2 complex mutations and epigenetic modifications including DNA methylation of Keap1 in colorectal cancer development.
The Nrf2-Keap1-ARE signaling pathways
Kelch-like ECH-associated protein 1 (Keap1) protein acts through its Neh2 Glu-Thr-Gly-Glu (ETGE) and asp-leu-Gly (DLG) motifs. Both of these motifs bind to similar sites on the underside of the Keap1 kelch motif. Keap1 is rich in cysteine with 27 cysteine residues in human protein. Basic residues which are set closely with some of these cysteines can be therefore suitable targets for electrophiles and oxidants. 23 The cysteine residues are modified by electrophiles which are known as the cysteine code. The cysteine code hypothesis suggests that structurally different Nrf2 agents affect different Keap1 cysteines. Inhibition and polyubiquitination of Nrf2 which are mediated by cysteine alteration result in conformational changes in Keap1 which disturb the interaction between Nrf2 DLG and Keap1 kelch domains. Cys151, Cys273, and Cys288 play important roles in regulation of Nrf2 activity; therefore, suppression of Nrf2 mediated by Cys273, Cys288, and Cys151 is necessary for activation of Nrf2 by inducers.24,25 The cysteine residues of Keap1 are key players in encoding the sensor mechanism. These residues in Keap1 (C151, C273, and C288) are recognized by both in vitro alkylation and in vivo site-directed mutagenesis assays.26,27 The direct Nrf2 inducer is alkylate C151, which appears to be the major site.28,29 Nrf2 has six conserved domains which are homologous to Neh1–6 of the chicken, mouse, and human. The Neh1 domain has a cap ‘n’ collar (CNC)-type basic leucine zipper which is required for DNA binding and dimerization with other transcription factors. In addition, this domain acts as a nuclear localization signal (NLS). 30 Neh2 domain binds to the Kelch domain of Keap1 through ubiquitin conjugation via seven lysine residues and acts as a negative regulator of Nrf2 and finally leads to its degradation.31,32 Neh3 is necessary for transcriptional activation along with the coactivator chromodomain helicase DNA binding protein 6 (CHD6). Nevertheless, the exact role of CHD6 is yet to be fully understood. 33 Neh4 and Neh5 act synergically with two independent transactivation domains which interact with the cyclic adenosine monophosphate (cAMP)-responsive element binding protein (CREB or CBP). 33 Nrf2 translocates to the nucleus in order to form a heterodimer with small Maf protein and activate transcription of several different types of genes by binding to the ARE sequence 34 (Figure 2).
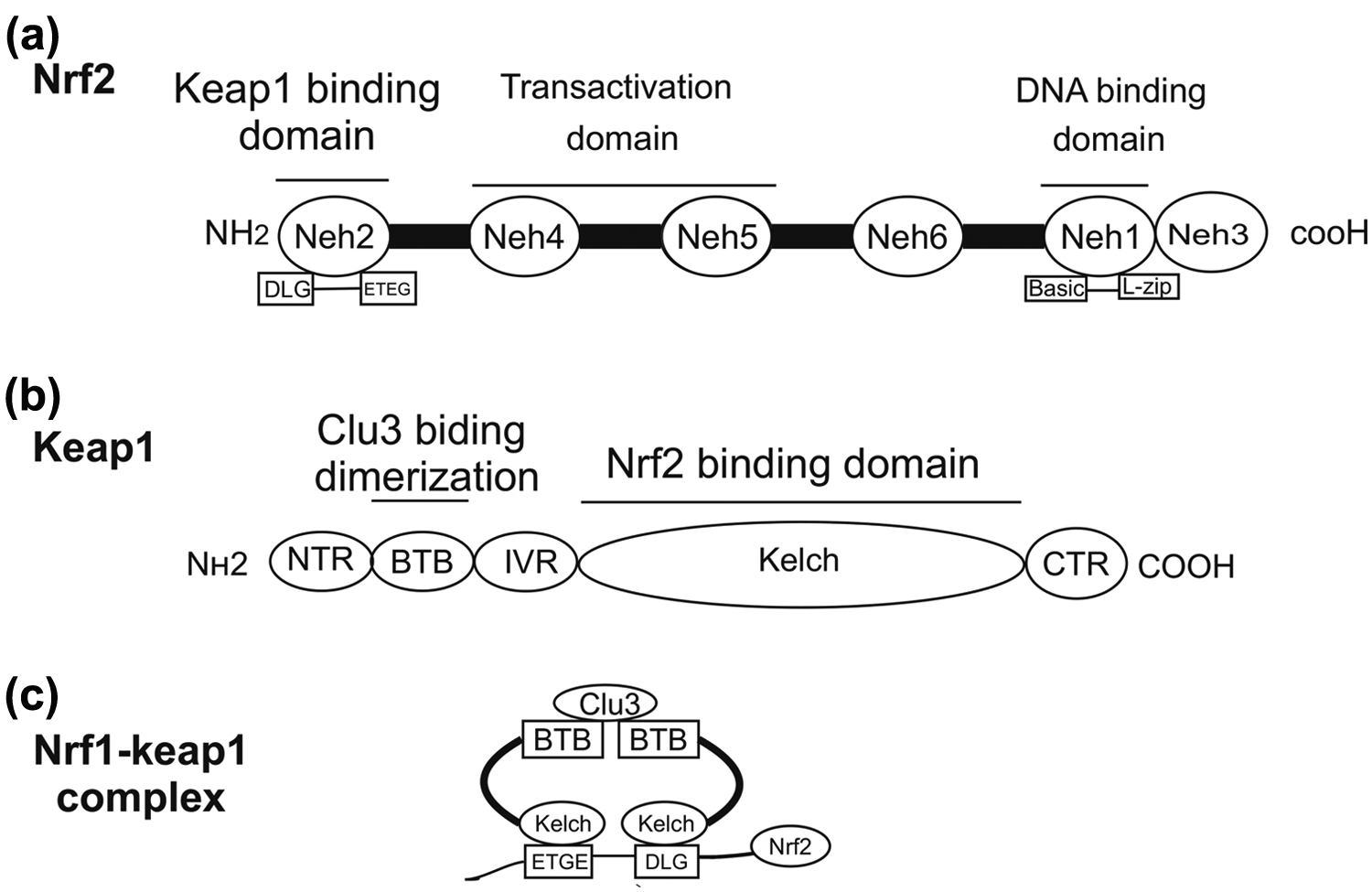
Structures of (a) Nrf2 and (b) Keap1 and (c) interaction between Nrf2 (ETGE domain) and Keap1 (kelch domain).
Nrf2-Keap1 signaling pathway functions partially through cross talk with important signaling pathways involved in tumor progression and chemoresistance. Nuclear factor-κB (NF-κB) is a family of transcription factors that includes p65, RelB, c-rel, p50, and p52, which mediates immune responses to oxidative stress and inflammation followed by tumor initiation and progression.35,36 NF-κBp65 negatively regulates Nrf2 signaling in which p65 inhibits Nrf2- and ARE-dependent gene transcriptions. 37
Notch1 is a transmembrane receptor of Notch family that promotes Notch-signaling by activating Rbpjκ transcription factor. This molecule is highly conserved in the Nrf2 promoter regions ARE. 38 Notch-signaling pathway is involved in cell fate decisions including cell survival, differentiation, and apoptosis. The gene regulatory region of the major Notch1 transcript has been identified to be a functional ARE through which Nrf2 can regulate Notch1 gene expression (Figure 2). 39
The dual role of Nrf2 in CRC
Activation and different expression levels of the molecules involved in Nrf2 pathway have been observed in tumor and normal colorectal tissues. 40 These tissues may be auto-regulated in response to different amount of Nrf2-Keap1 or Nrf2-Bach1 chain. Cancer cells can abduct the Nrf2 pathway to protect themselves from the cellular stress related to enhanced proliferation, hypoxic environment, and deregulated protein synthesis. 41 The difference in the expression profile of Nrf2 pathway between the tumor and normal tissues may suggest a novel therapeutic approach against CRC. 42
Although Nrf2 prevents tumor initiation, progression, and cancer metastasis by omitting carcinogens, reactive oxygen species (ROS), and other DNA-damaging factors, accumulation of DNA damages can result in Nrf2 hyperactivity, which in turn enhances the rate of survival of cancer cells when exposed to high levels of endogenous ROS and to escape apoptosis.25,43,44 In a mutant cell, persistent increase in Nrf2 leads to upregulated expression of downstream genes which consequently induce metabolic reprogramming and improved cell proliferation rate. High Nrf2 levels in various cancer cells are relevant to poor prognosis, radio- and chemoresistance, and aggressive proliferation of cancer cells. 40 Activation of Nrf2 pathway mediates cell protection in the early stages of tumorigenesis; however, it plays a destructive role in later stages. Therefore, elevation of Nrf2 pathway activity is an efficient approach for cancer prevention, and Nrf2 inhibition is advantageous for treatment of early-stage cancer.41,45 Nrf2 protects normal cells from being transformed into cancer cells as well as protects cancer cells from cellular stress. This dual role of Nrf2 augments the survival rate of cancer cell and enhances cancer progression (Figure 3). 46

Dual role of Nrf2 in normal and cancer cells. (a) Cancer prevention mediated by Nrf2 activation in normal cell. (b) While in cancer cell anticancer therapy is mediated by Nrf2 inhibitors.
Inflammation and oxidative stress in carcinogenesis of CRC
Chronic inflammation is a major factor involved in CRC. Inflammation can transform a normal tissue into a cancer tissue.40,47 Various types of activated inflammatory-immune cells, including mast cells, neutrophils, leukocytes, macrophages, monocytes, eosinophils, dendritic cells, phagocytes, and natural killer cells, are produced in an inflamed tissue. These cells create ROS and/or reactive nitrogen species (RNS) which can consequently cause DNA damage and trigger carcinogenesis by activation of oncogenes and/or inhibition of tumor-suppressor genes. 48
Tumor-suppressor function of Nrf2
In response to cellular stress and premalignant or chemopreventive compounds, Nrf2 dissociates from its repressor (Keap1) prior to its translocation to the nucleus in order to bind to ARE promoter of genes that are involved in cellular homeostasis and mediate its effect by transcriptional activity. The molecular structure and physiological regulation of this complex have been vastly investigated in detail.25,49,50 The Nrf2-Keap1 pathway is necessary to maintain homeostasis since adaptive responses to endogenous and exogenous stresses are essential for cell survival. Interaction between Nrf2 and Keap1 in normal condition is stable at low concentrations, which regulates cellular homeostasis, while at high concentrations, it mediates cellular defense against oxidative stress, including ROS, such as hydrogen peroxide (H2O2) and RNS, for instance, nitric oxide (NO).40,51 Regulation of these molecules is mediated by Nrf2. More than 100 genes are regulated by Nrf2 that binds to the response elements on DNA, which are known as AREs or electrophile response elements (EpREs), and upregulates cellular stress response genes by its transcriptional activity.18,20 In fact, upregulation of more than 20 of such cytoprotective “phase 2” enzymes are mediated by Nrf2 that can also control drug transport via the activation of the multidrug resistance–associated gene family. 50 Chemopreventive activity of many photochemical or synthetic compounds is mediated by Nrf2-dependent responses, including phase-II detoxifying enzymes, antioxidants, and transporters that protect cells from harmful events, such as carcinogenic insults. Consequently, several studies have used Nrf2-null mice for further investigation of the fundamental role of Nrf2 in cancer protection. For instance, reduced levels of phase-II genes, such as glutathione S-transferase (GST), NAD(P)H:quinone oxidoreductase 1 (NQO1), and glutamate cysteine ligase (GCL), have been detected in Nrf2-null mice.52–56 Interestingly, many studies have shown that chemopreventive compounds stimulate Nrf2 activity, for example, sulforaphane (cruciferous vegetables) is a potent Nrf2 inducers. 57 Many chemopreventive compounds induce Nrf2 activity, including curcumin (a widely used spice), epigallocatechin-3-gallate (EGCG; green tea), resveratrol (grapes), caffeic acid phenethyl ester (conifer trees), wasabi (Japanese horseradish), cafestol and kahweol (coffee), cinnamonyl-based compounds, zerumbone (ginger), garlic organosulfur compounds (garlic), lycopene (tomato), carnosol (rosemany), and avicins (Bentham plant).58–74 In addition to the phytochemicals, deterministic synthetic chemicals, such as oltipraz (a substituted 1,2-dithiole-3-thione), 2-indol-3-yl-methylenequinuclidin-3-ols (an indole analogue), and synthetic triterpenoid 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid (CDDO) and its derivative 1-[2-cyano-3-,12-dioxooleana-1,9(11)-dien-28-oyl] imidazole (CDDO-Im), are other powerful Nrf2 inducers (Figure 4).75–78
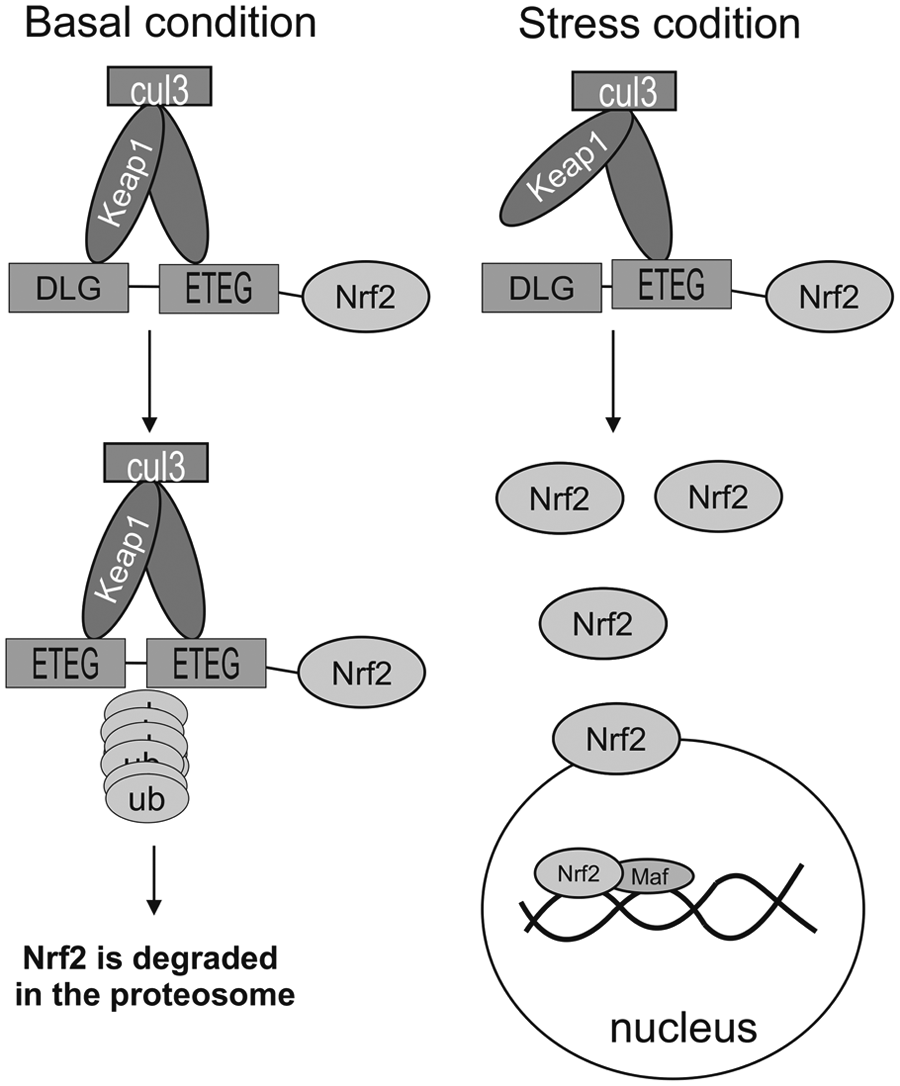
The Nrf2 pathway function in basal conditions and cellular stress conditions.
Oncogenic functions of Nrf2
Nrf2 over-activity was detected in many types of cancer including CRC. Several mechanisms including Nrf2 and Keap1 genes, epigenetic mechanisms, and various proteins that disturb the binding between Keap1 and Nrf2 have been revealed to lead the over-activation of Nrf2. 23 Furthermore, chemo- and radio-resistance provide Nrf2 overactivation-mediated growth advantage in cancer cells that enables these cells to induce proliferation and evade apoptosis. 79 Thus, cancer patients with hyperactive Nrf2 often have a poor prognosis. 80 The efficacy of chemo- and radiotherapy is increased by silencing Nrf2, since the expression of this gene results in radio- and chemoresistance. 80
Nrf2 as a therapeutic target in CRC
The suppressive mechanism of Keap1 in Nrf2 pathway has been previously discussed. 81 However, further studies are required to fully specify the Keap1-Nrf2 pathway in detail, particularly in human.9,82 It has been shown that Nrf2 responds to the oxidative stress via regulating gene transcription. 83 It appears that an internal ribosomal entry site (IRES) within the 5′-untranslated region of the human Nrf2 messenger RNA (mRNA) is involved in redox-sensitive Nrf2-mediated translation. 83 This indicates that Keap1-Nrf2-ARE signaling pathway could be targeted for CRC chemoprevention. Therefore, one of the important mechanisms in cellular protection against oxidative and electrophilic stress is Nrf2-ARE pathway that mediates its effect via the proteins involved in antioxidative stress and detoxification as well as elimination of carcinogens and electrophiles through induction of antioxidative enzymes.5,47,84
Cell lines. It has been shown that Nrf2 plays an important role in HT-29 cell sensitivity to 5-fluorouracil (5-FU). Activation of Nrf2-ARE pathway leads to expressions of antioxidant enzymes in drug-resistant CRC cell line. HT-29 cells were treated with 5-FU, and the nuclear and cytosolic levels of Nrf2 were measured in both control and 5-FU-treated cells in order to find out whether the Nrf2-ARE signaling pathway was activated.85,86 Activation of Nrf2-ARE pathway is dependent on the accumulation and translocation of Nrf2 to the nucleus. The neutral red cytotoxicity test demonstrated that cell survival rate of 5-FU-treated cells, which were transfected with Nrf2 small interfering RNA (siRNA), was lower than that in the cells transfected with non-targeting siRNA. 87 These results suggest that Nrf2 pathway can induce 5-FU resistance in HT-29 which can be broken down by inhibiting the Nrf2 pathway.85,88
Animals. Since Nrf2 is a fundamental transcriptional factor in cytoprotection and oxidative hemostasis, using APC (Min/+) and Nrf2 knockout mice in colorectal carcinogenesis and interposition studies would be extremely informative and beneficial to investigate the potential role of Nrf2 as a chemopreventive target for CRC.89,90 Another ideal model to study oxidative stress–induced carcinogenesis of the colon is azoxymethane (AOM)-dextran sulfate sodium (DSS)-Nrf2 knockout mice. Hence, methods of inhibiting NF-κB can be used to explicitly determine the emerging roles of cross talk between the Nrf2 and NF-κB transcriptional factors which could have therapeutic applications in cancer and inflammatory diseases. While a key player in the inflammatory response, ectopic activation of NF-κB is recurrently considered in many cancers. These findings clearly show the important protective regulatory role of Nrf2 against early-phase inflammation-mediated CRC carcinogenesis. 91 Many studies have shown the higher sensitivity of the Nrf2-null mice to toxic and carcinogenic effect of a variety of xenobiotics, including benzo(a)pyrene, diesel exhaust, cigarette smoke, N-nitrosobutyl(4-hydroxybutyl) amine, pentachlorophenol, and acetaminophen, compared to the wild-type mice. 78 Furthermore, it has been demonstrated that Nrf2 knockout mice are sensitive to severe colitis mediated by inflammatory agents, such as dextran sulfate sodium (DSS), compared to the wild-type mice.78,92–96 The comparison between these two different types of mice has revealed the importance of the Nrf2 pathway in carcinogenesis and chemoprevention. It has been shown that the risk of CRC formation and dysplasia was higher in the Nrf2 knockout mice exposed to carcinogen AOM followed by DSS treatment compared to the wild-type mice with similar treatment. The Nrf2 knockout mice treated with AOM-DSS were significantly more talented to grow colonic tumor compared to the wild-type mice. Moreover, in Nrf2 knockout mice, using nitrotyrosine, cellular damage of inflamed mucous and increased chance of CRC have been reported. Therefore, Nrf2 is necessary for the protective pathways against inflammation-associated CRC. 89 The results of this study have shown that downregulated expression of downstream genes of Nrf2 pathway, including phase-II detoxifying and other antioxidative stress genes, is accompanied by simultaneous upregulated expression of inflammatory markers in Nrf2 knockout mice compared to wild-type mice.
Human. Many epidemiologic and clinical investigations have revealed that there is a correlation between inflammatory bowel disease, ulcerative colitis, and Crohn’s disease and increased risk of CRC.97–100 This increased risk of CRC could be due to the accumulation of ROS and RNS that are produced by inflammatory cells, including white blood cells. Inflammatory cells infiltrate the wounded colonic mucosa and create oxidative stress. It appears that ROS and RNS are the most substantial factors involved in transformation of chronic inflammation in cancer. 101 Exogenous antioxidants and activation of endogenous and phase-II antioxidant potentially inhibit the colorectal disorders. It has been recently emerged that Nrf2 can be applied as a novel target for the prevention of CRC. 7 Nrf2 pathway is the most important signaling pathway in cell protection and survival against xenobiotics and oxidative stress that creates cell protection against the development of some diseases, such as cancer. 102 Nearly 50 various antioxidants and cytoprotective genes are controlled by transcriptional activity of Nrf2, which is a CNC-type basic leucine zipper transcription factor. Keap1 is a repressor factor and negative regulator of cytoplasmic Nrf2 in normal condition. The ubiquitylation and degradation of Nrf2 are mediated by cullin-3 (CUL3) E3 ligase in the proteasome. In response to oxidative stress, Nrf2 dissociates from Keap1 and translocates to the nucleus under physiological conditions. Low levels of nuclear Nrf2 are sufficient for the protection of cellular homeostasis. 32 The heterodimer Nrf2 and small Maf proteins connect to the ARE in the promoter regions of the genes coding various antioxidant proteins of many phase-II detoxifying enzymes and mediate their transcription and consequently control cellular stress. 103
Targeting Nrf2-Keap1 signaling pathway as a therapeutic axis
Targeting Nrf2 pathway would be the rational and potential pathway to prevent or treat CRC. Many of the chemopreventive agents that target Nrf2 can prevent cancer onset and progression.86,104,105 Evidence-based investigations have revealed that chemopreventive compounds suppress inflammation and effectively prevent colitis-associated CRC.11,106–109 Nevertheless, understanding the molecular mechanisms of Nrf2 signaling underlying CRC initiation and progression can lead to design of proper applications for diagnosis and treatment of the disease. For example, chemopreventive agents such as isothiocyanate sulforaphane have potent anticancer activity via increasing the expression of phase-II detoxifying and antioxidant enzymes.110,111 To apply experimental outcomes of targeting Nrf2 for clinical purposes, it is essential to propose appropriate clinical trials as well as pharmacokinetic profile and genetic studies on polymorphisms of key downstream genes. 112 Another strategy would be the inhibition of inflammation by Nrf2-targeted-based therapeutics,113,114 for example, inhibition of Nrf2 in combination with targeting multiple pathways, including the migration inhibitory factor (MIF), cyclooxygenase-2 (Cox-2), Nf-κB, tumor necrosis factor alpha (TNFα), inducible nitric oxide synthase (iNOS), protein kinase B (PKB), and CXC chemokines which are important anti-inflammatory targets, may lead to better efficacy from CRC chemoprevention in the future.5,115–117
Conclusion
Considering the intimate association between ROS/RNS and the progression of cancers, it is believed that chemopreventive compounds targeting Nrf2 not only inhibit the anomalous inflammation and decrease the oxidative stress but they also inhibit the progression of CRC. In this review, we have discussed various underlying mechanisms of Nrf2 regulations to explain dual role of Nrf2 in CRC initiation and progression. Understanding these mechanisms has offered novel approaches to control Nrf2 expression in order to manage cellular redox homeostasis. However, these findings about Nrf2-Keap1 mechanisms of action not only will expand our knowledge about Nrf2-dependent molecular basis of CRC but also consider this axis for therapeutic purposes.
Footnotes
Acknowledgements
The authors also gratefully acknowledge Immunology Research Center (IRC), Tabriz University of Medical Sciences for technical support.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was financially supported by Liver and Gastrointestinal Diseases Research Center, Tabriz University of Medical Sciences.