
Abstract
MTA3 overexpression has been implicated in carcinogenesis. The aim of the present study was to explore the clinical significance and biological roles of MTA3 in human colorectal cancer and colorectal cancer cells. A total of 80 cases of colorectal cancer tissues were examined by immunohistochemistry for MTA3 protein expression. We analyzed the relationship between MTA3 and clinical factors and the results showed that MTA3 was overexpressed in 51.25% (41/80) cancer cases. There was significant associations between MTA3 overexpression and advanced TNM stage (p = 0.0086) and Ki67 index (p = 0.001). We overexpressed MTA3 in LoVo cells and depleted its expression in HCT15 cells. The results showed that MTA3 promoted cancer cell proliferation, invasion, migration, and cell cycle progression, and inhibited 5-fluorouracil-induced apoptosis in LoVo cell line. MTA3 depletion in HCT15 cell line showed the opposite effects. In addition, we found that MTA3 positively regulated cell cycle proteins including cyclin D1 and cyclin E. It also upregulated Bcl2 and downregulated Bax expression. Furthermore, we found that MTA3 could activate Wnt signaling pathway by upregulating Wnt target proteins. Our results demonstrated that MTA3 overexpression contributes to colorectal cancer carcinogenesis, progression, and chemoresistance. MTA3 could serve as a potential therapeutic target in colorectal cancer.
Keywords
Introduction
Colorectal cancer (CRC) is a leading cause of cancer-related deaths in many countries.1–3 Despite development in surgery and adjuvant therapies, including chemotherapy and radiotherapy, the prognosis remains poor in CRC patients, especially in those with advanced-stage disease. The limitation of chemo- and radio-therapies suggests a need for new treatment, such as targeted therapy, which may greatly increase the efficiency of cytotoxic drugs. Therefore, it is of paramount importance to identify gene targets that could be used as cancer biomarkers in predicting CRC aggressiveness.
MTA family includes MTA1, MTA2, and MTA3, all of which serve as the subunit of Mi-2/NuRD. Recent studies reported MTA3 as a potential oncogene in mammals. MTA3 was also reported to participate in B-lymphocyte development. 4 In plasmacytoma cell lines, overexpression of BCL6 and MTA3 causes downregulation of plasma cell differentiation genes.5,6 MTA3 upregulation could prevent epithelial–mesenchymal transition (EMT) by directly inhibiting Snail and upregulating E-cadherin. 7 It was found that MTA3 facilitated G2/M progression in proliferating mouse granulosa cells. 8 Furthermore, MTA3 was reported as an independent unfavorable prognostic parameter in uterine non-endometrioid carcinoma. 9 MTA3 also serves as an oncogene and prognostic factor in human lung cancers. 10
Despite growing evidences showing MTA3 as an important cancer-related protein, its involvement in CRC has not been investigated. Herein, we explored clinical significance of MTA3 in CRC. In addition, we investigated the role of MTA3 protein in CRC progression.
Materials and methods
Patients and specimens
The study protocol was approved by the institutional review board of China Medical University. A total of 80 cases of cancer tissues and 15 cases of normal control paraffin-embedded tissues were retrieved from the Pathology Archive in the First Affiliated Hospital of China Medical University from 2008 to 2013. The histological diagnosis and grade of differentiation were evaluated by two independent pathologists according to the World Health Organization (WHO) guidelines of classification.
Immunohistochemical staining
Tumor specimens were fixed with neutral formalin, embedded in paraffin, and 4 µm thick sections were prepared. The sections were deparaffinized in xylene, rehydrated in graded alcohol series, and boiled in 0.01 M citrate buffer for 2 min. A volume of 0.3% hydrogen peroxide was added and incubated for 10 min followed by incubating the sections with non-immune goat serum. The primary antibodies were used as follows: MTA3 diluted at 1:800 (Proteintech, USA) and Ki67 (ready-to-use antibody; Fuzhou Maixin Biotech, Fuzhou, China). Immunostaining was performed using the EliVision plus kit (Fuzhou Maixin Biotech). DAB-Plus kit (Fuzhou Maixin Biotech) was used for 3,3′-diaminobenzidine (DAB) staining. Counterstaining with hematoxylin was performed and the sections were dehydrated in ethanol before mounting.
Immunohistochemical assessment
Two independent, blinded investigators examined all tumor slides randomly. Five views were examined per slide, and 100 cells were observed at a magnification of 400×. MTA3 expression levels in colorectal tissue samples were divided into MTA3 high expression and MTA3 low/negative expression. Absence of reactivity was graded as negative. For nuclear distribution, samples with nuclear staining in less than 10% of cells were graded as MTA3 low expression and samples with nuclear staining in more than 10% cells were graded as MTA3 high expression. According to the classification criteria in a previous studie, 11 the Ki67 score was defined as the percentage of tumor cells with nuclear staining. Ki67 was regarded as high expression at score > 41%.
Cell culture and transfection
LoVo and HCT15 cell lines were obtained from American Type Culture Collection (ATCC, USA). The cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) and McCoy’s 5A (Invitrogen, USA) medium containing 10% fetal calf serum (Invitrogen). Cells were cultured on plastic dishes and were passed every 2 days with 0.25% trypsin.
pcDNA3.1-MTA3 and pcDNA3.1 plasmids were used for transfection. For MTA3 plasmid transfection, Attractene (Qiagen, Hilden, Germany) reagent was used according to the manufacturer’s instructions.
For RNA interference (RNAi) transfection, DharmaFECT1 (GE Healthcare, USA) was used. The transfection efficiency was assessed 48 h later by western blotting. ON-TARGETplus MTA3 siRNA pool was purchased from Dharmacon (GE Healthcare). ON-TARGETplus Non-Targeting siRNAs were used as a negative control.
Western blotting
Total proteins from cells were extracted in radioimmunoprecipitation assay (RIPA) lysis buffer and quantified using the Bradford method. Protein samples were transferred to polyvinylidene fluoride (PVDF) membranes (Merck Millipore, USA) and incubated overnight at 4°C with primary antibody against MTA3 (1:1000; Proteintech), cyclin D1, cyclin E, Bcl2, Bax, MMP7, c-myc (1:1000; Cell Signaling Technology, USA), β-catenin (1:1500, BD Biosciences, USA), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:2000; Santa Cruz Biotechnology, USA). After incubation with peroxidase-coupled secondary antibody (1:2000, Santa Cruz Biotechnology) at 37°C for 2 h, target proteins on PVDF membranes were visualized using Pierce ECL kit (Thermo Fisher Scientific, USA) and obtained using DNR Imaging System (DNR Bio-Imaging System, Neve Yamin, Israel).
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (PCR) was performed using SYBR Green master mix kit from Applied Biosystems (USA). PCR was performed using 7900 Real-Time PCR System (Applied Biosystems). β-actin was used as the endogenous control. The relative levels of gene expression were represented as ΔCt = Ct gene − Ct reference, and the fold change of gene expression was calculated by the 2−ΔΔCt method. The primer sequences are as follows: MTA3 forward: 5′-CCCACCCAGTCAGAAGAAGA-3′, MTA3 reverse: 5′-TTGGACTCCCAGTGTTTCG-3′, β-actin forward: 5′-ATAGCACAGCCTGGATAGCAACGTAC-3′, β-actin reverse: 5′-CACCTTCTACAATGAGCTGCGTGTG-3′.
Colony formation and MTT (3-(4,5-dimethyl-2-thiazoyl)-2,5-diphenyl-2H-tetrazolium bromide) assays
For colony formation assay, cells were transfected for 48 h and plated onto three 6-cm cell culture dishes (1000 cells). Cells were incubated for 2 weeks in medium. Plates were washed with phosphate-buffered saline (PBS) and stained with Giemsa. The number of colonies with more than 50 cells was counted manually using a microscope.
For the MTT (3-(4,5-dimethyl-2-thiazoyl)-2,5-diphenyl-2H-tetrazolium bromide) assay, 24 h after transfection, cells were plated on 96-well plates at a concentration of approximately 2000 cells per well and cultured for 5 days. For quantitation of cell proliferation rate, 20 µL of 5 mg/mL MTT solution was added to each well and incubated for 4 h at 37°C. The medium was removed from each well and 150 µL of dimethyl sulfoxide (DMSO) was added to the well. The plate was measured at 490 nm.
Flow cytometry for cell cycle and apoptosis analysis
Cells were seeded onto 6-cm tissue culture dishes. After 12 h, cells were transfected with the indicated amounts of small interfering RNA (siRNA)/plasmid. At the indicated time points, cells were harvested, fixed in 1% paraformaldehyde, washed with PBS, and stained with 5 mg/mL propidium iodide in PBS supplemented with RNase A (Roche, USA) for 30 min at room temperature. Data were collected using BD Systems (Becton Dickinson, USA).
Cell apoptosis rate was examined using Annexin V/PI kit from BD Biosciences. After transfection, the cells were harvested and washed with cold PBS buffer, followed by resuspension in 250 µL binding buffer. After that, the cells were stained with Annexin V/FITC and propidium iodide (PI) for 30 min in dark. Apoptosis rates were analyzed by FACSCalibur flow cytometer (Becton Dickinson).
TOP-Flash reporter assay
TOP-Flash reporter plasmid and Renilla luciferase reporter plasmid were transfected into cells using Attractene (Qiagen). After 48 h, dual luciferase reporter kit (Promega, USA) was used to detect TOP-Flash luciferase activity.
Statistical analysis
SPSS software was used for all statistical analyses. A chi-square test was used to examine possible correlations between MTA3 expression and clinicopathologic factors. p value < 0.05 was regarded as statistical significance.
Results
Expression pattern and clinical significance of MTA3 protein in CRC
We analyzed MTA3 protein expression in 80 CRC specimens and 15 normal colon specimens by immunohistochemistry. In CRC tissues, MTA3 protein was mainly located in the nucleus (Figure 1(b)–(d)). We divided MTA3 protein levels into negative/low expression and high expression. We observed high MTA3 protein expression in 51.25% (41/ 80) specimens. In normal colon tissues, MTA3 was negative in most cells (Figure 1(a)). We also found weak MTA3 expression in the basal crypt zones, in which were located progenitor cells.

Expression of MTA3 protein in colon cancer tissues. (a) Negative MTA3 staining in normal colon mucosa. (b) Negative CARMA3 staining in case of adenocarcinoma. (c) Strong nuclear MTA3 expression in case of mucinous adenocarcinoma. (d) Strong nuclear MTA3 staining in case of signet ring cell carcinoma (magnification: 400×).
Correlation of MTA3 protein with clinicopathological factors was analyzed. As shown in Table 1, of the 80 tumor samples, 29.4%, 44.8%, 54.5%, and 91.6% of the stage I, II, III, and IV tumors showed MTA3 positivity. There was a close correlation between high MTA3 protein and TNM stage (p = 0.0086). MTA3 expression rate was found to be higher in tumors with positive lymph node metastasis (57.6%) compared with those without (44 %). However, the difference was not significant (p = 0.4238). No statistical significance was found between MTA3 levels and age, gender, and differentiation. We also analyzed the relationship between MTA3 positivity and Ki67 proliferating index. Cases with high MTA3 protein expression also had high Ki67 status (p = 0.001; Figure 2).
Distribution of MTA3 status in colorectal cancer according to clinicopathological characteristics.


Correlation of MTA3 with Ki67 proliferation index. Correlation of MTA3 expression with high Ki67 proliferation index. Cases that have high levels of MTA3 expression showed high proliferation indexes, indicated by strong nuclear Ki67 label. (magnification: 400×).
MTA3 regulates proliferation, invasion, migration, and apoptosis of CRC cells
Protein expression of MTA3 was analyzed in a panel of CRC cell lines and normal cell line HCOEPIC. As shown in Figure 3(a), expression of MTA3 protein was higher in CRC cell lines than normal cell line. We found high MTA3 endogenous expression in HCT15 cell line and low MTA3 endogenous expression in LoVo cell line (Figure 3(a)). To explore the biological role of MTA3 in CRC cells, we used MTA3-specific siRNA to knockdown MTA3 in HCT15 cells and transfected MTA3 plasmid to upregulate its expression in LoVo cells. The transfection efficiency was shown in Figure 3(b).

Expression of MTA3 in colorectal cancer cell lines. (a) Western blot showed that MTA3 expression was higher in colorectal cancer cell lines than normal cell line HCOEPIC. (b) Western blot and real-time PCR analyses showed that siRNA treatment markedly decreases MTA3 levels in HCT15 cells and MTA3 transfection significantly increased its expression in LoVo cells.
Colony formation and MTT assay demonstrated that transfection of MTA3 plasmid promoted proliferation rate and colony number, while MTA3 depletion significantly reduced proliferation rate and colony formation ability (Figure 4(a) and (b)).
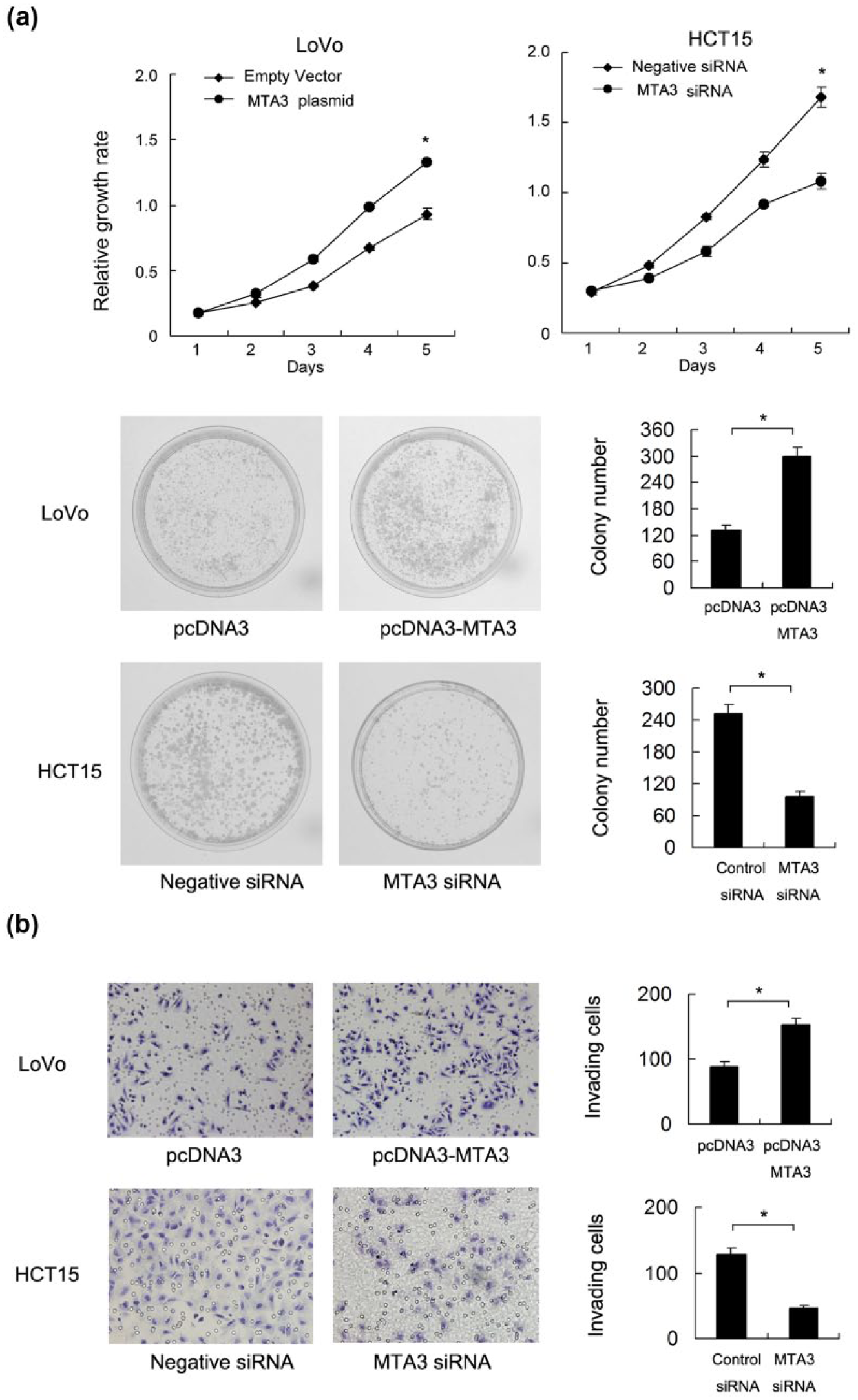
MTA3 promotes CRC cell proliferation and invasion. (a) MTT assay showed that MTA3 knockdown inhibited proliferation in HCT15 cell lines (Day 5: p < 0.05). MTA3 overexpression promoted proliferation in LoVo cell line (Day 5: p < 0.05). (b) Colony formation assay showed a downregulation in colony formation number in the HCT15 with siRNA treatment (p < 0.05). MTA3 transfection upregulated colony formation number in LoVo cells (p < 0.05). (c) Matrigel invasion assay showed that MTA3 depletion downregulated invading cell number, while MTA3 overexpression upregulated invading cell number.
Matrigel invasion assay and wound healing assay (migration assay) showed that MTA3 plasmid promoted invading (Figure 4(c)) and migrating ability (Figure 5), while the depletion of MTA3 led to a significant reduction of the invading and migrating ability.

MTA3 promotes CRC cell migration. Wound healing assay demonstrated that MTA3 overexpression accelerated wound healing speed and migration ability. MTA3 depletion showed the opposite effect.
Flow cytometry (PI staining) was used to detect cell cycle change after introduction of MTA3 siRNA and plasmid. As shown in Figure 6, MTA3 transfection increased S phase percentage and decreased G1 phase percentage in LoVo cells. MTA3 siRNA decreased S phase percentage and increased G1 phase percentage in HCT15 cells.

MTA3 facilitates cell cycle progression. MTA3 knockdown increased G1 phase cells and decreased S phase percentage in HCT15 cells. MTA3 overexpression increased S phase LoVo cells and decreased G1 phase cells.
Annexin V kit was used to detect 5-fluorouracil (5-FU)-induced apoptosis in HCT15 and LoVo cells treated with MTA3 siRNA/plasmid. A significant increase in early and late apoptosis was found in HCT15 cells with MTA3 depletion compared with negative control, indicating that MTA3 depletion increases apoptosis rate of the CRC cells during 5-FU treatment (10 nmol/L, 18 h). In LoVo cells transfected with MTA3 plasmid, the percentage of total apoptosis was decreased (Figure 7(b)). In addition, the cell viability of HCT15 cells treated with siRNA was lower than control after 48 h of 5-FU treatment. MTA3 plasmid showed the opposite effect in LoVo cells (Figure 7(a)).Together, these results demonstrated that MTA3 modulated proliferation, invasion, migration, and apoptosis in CRC cells.

MTA3 reduces 5-FU-induced apoptosis. (a) MTT assay showed that MTA3 overexpression increased LoVo cell viability after 48 h of 5-FU treatment. (b) Annexin V/PI analysis showed that MTA3 overexpression downregulated apoptosis rate. MTA3 depletion exhibited the opposite effects.
MTA3 regulates cell cycle and apoptosis-related proteins in CRC cells
To understand the mechanism of MTA3-induced cell growth and invasion, we analyzed several cell cycle–related factors. We observed that after MTA3 plasmid transfection, the levels of cyclin D1, cyclin E, and Bcl2 were significantly upregulated while p21 and Bax expression were downregulated. MTA3 siRNA depletion in HCT15 cells showed the opposite effects on these proteins (Figure 8).

MTA3 regulates cell cycle and apoptosis-related proteins. MTA3 plasmid transfection upregulated the levels of cyclin D1, cyclin E, and Bcl2, and downregulated p21 and Bax expression. MTA3 siRNA depletion in HCT15 cells showed the opposite effects on these proteins.
MTA3 regulates Wnt signaling pathway
We further explored the potential mechanism of MTA3-induced cell proliferation, invasion, and chemoresistance. Western blot showed that MTA3 depletion downregulated while MTA3 overexpression upregulated protein expression of c-myc and MMP7 (Figure 9(a)). TOP-Flash contains TCF/LEF binding sites that indicate the level of Wnt activation induced by binding of nuclear β-catenin to TCF/LEF transcription factor. Using TOP-Flash luciferase reporter assay, we demonstrated that MTA3 depletion downregulated TOP-Flash activity, while MTA3 overexpression upregulated luciferase activity, suggesting MTA3 positively regulates malignant Wnt signaling pathway (Figure 9(b)). Western blot analysis also showed that nuclear β-catenin was increased after MTA3 transfection and decreased after its depletion (Figure 9(a)).
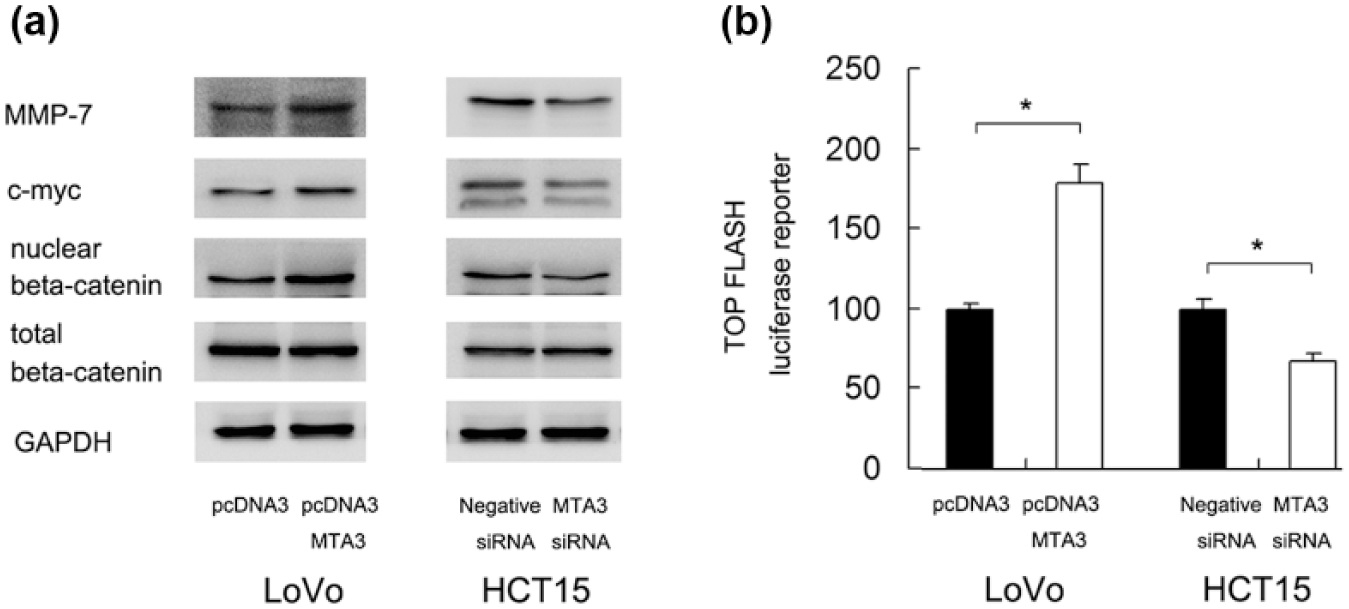
MTA3 regulates Wnt signaling pathway. (a) Western blot showed that MTA3 depletion downregulated while MTA3 overexpression upregulated protein expression of c-myc, MMP7, and nuclear β-catenin. (b). TOP-Flash luciferase reporter assay demonstrated that MTA3 depletion downregulated TOP-Flash activity, while MTA3 overexpression upregulated luciferase activity.
Discussion
MTA3 upregulation has been reported in human choriocarcinoma and non-endometrioid adenocarcinoma.7,12 MTA3 protein overexpression in non–small cell lung cancer was associated with poor patient prognosis. 10 It was reported that MTA3 is an independent prognostic parameter in uterine non-endometrioid carcinomas. 12 In addition, MTA3 expression was an independent prognostic factor in patients with gastroesophageal junction adenocarcinoma. 13 To date, its clinical significance and biological characteristics in colorectal carcinoma have not been well investigated. In this study, we demonstrated MTA3 protein expression was higher in CRC tissues than that in normal colon tissues. Significant correlation was found between high MTA3 expression and advanced TNM stage and Ki67 proliferation index, indicating its association with malignant progression of CRC.
To investigate biological roles of MTA3, we treated HCT15 cells with siRNA and transfected MTA3 plasmid into LoVo cells. MTA3 knockdown induced an obvious decrease in the growth rate and colony number in HCT15 cells, while its overexpression in LoVo cells caused the opposite effects, which was in accordance with previous studies. 10 To investigate the potential mechanism of MTA3-induced cell proliferation, we checked the change of cell cycle progression. The results showed that MTA3 depletion inhibited G1/S progression in HCT15 cells, while MTA3 overexpression facilitated G1/S cell cycle transition. In addition, western blot showed that MTA3 upregulated cyclin D1, cyclin E, and downregulated p27. Cyclin D1 and cyclin E interact with cyclin-dependent kinase (CDK) to form complexes and promote cell cycle progression.14,15 p27 serves as a cell cycle inhibitor and its level negatively correlates with cell growth rate. p27 expression is downregulated in colorectal carcinoma and predicts patient prognosis.16,17 These results were in accord with our immunohistochemical data showing positive correlation between MTA3 and Ki67 proliferation index, which demonstrated that MTA3 facilitated colon cancer proliferation by regulating cell cycle proteins.
In addition to the role of MTA3 on cell proliferation, we found that MTA3 positively regulated cell invasion and migration. The role of MTA3 on promoting cancer cell invasion has been reported in non–small cell lung cancer. 10 The role of MTA3 on migration has not been reported before. Further study showed that MTA3 enhanced 5-FU resistance and inhibited apoptosis of CRC cells. By checking apoptosis related proteins, we found that MTA3 overexpression upregulated Bcl2 and downregulated Bax expression. MTA3 depletion in HCT15 cells showed the opposite results. Both Bcl2 and Bax are involved in the apoptosis regulation. Bcl2 is an anti-apoptosis protein and Bax is a pro-apoptosis protein, both of them play important roles in various cancers. Upregulation of Bcl2 and downregulation of Bax could cause resistance to chemotherapy in CRC.18–20 Thus, the role of MTA3 on 5-FU resistance may be due to its regulation on Bcl2/Bax ratio.
Overactivation of Wnt/β-catenin pathway is one of the early events during colorectal carcinogenesis. 21 Wnt signaling pathway regulates multiple biological processes, including cell proliferation, invasion, and migration, mainly through modulation of target proteins such as c-myc, MMP7, and cyclin D1. 22 It has been reported that activation of canonical Wnt signaling induced chemoresistance of CRC.23,24 Many CRC patients demonstrate dysregulated Wnt signaling. Inhibition of Wnt cascade provides an alternative strategy for targeting chemoresistant cancers. Herein, we demonstrated that MTA3 positively regulated Wnt target proteins, MMP7 and c-myc, in CRC cells. MTA3 also induced nuclear localization of β-catenin and upregulation of TOP-Flash luciferase activity, indicating its positive role on Wnt signaling. Taken together, MTA3 regulates malignant behavior of CRC possibly through its positive role on Wnt signaling pathway.
In conclusion, this study demonstrated that MTA3 was overexpressed in CRC and correlated with advanced TNM stage and Ki67 proliferation index. MTA3 promoted CRC growth by regulating cell cycle progression and related proteins, including cyclin D1, cyclin E, and p27. MTA3 also facilitated invasion, migration, and chemoresistance with the upregulation of Bcl2/Bax ratio. In addition, MTA3 activated Wnt signaling. Our results indicated MTA3 as an important oncoprotein and potential therapeutic target in CRC.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was financially supported by grants from Key Technologies Research and Development Program of Liaoning Science and Technology Department (No. 2013020198-202) and Program of Shenyang Science and Technology Department (No. 131138).