
Abstract
Prostate cancer is the most commonly diagnosed malignancy and is the second leading deadly reason among male cancer. WDFY2, which is found to be a cancer-specific fusion gene with CDKN2D in ovarian cancer, is a new gene with unknown function in carcinogenesis. In this study, we investigated the role of WDFY2 in prostate cancer development. We examined WDFY2 expression in human prostate tissue specimens and prostate cancer cell lines BPH-1, LNCaP, PC3, and DU-145. Overexpression of WDFY2 was performed to evaluate the role of WDFY2 in cell proliferation, migration, and colony formation of prostate cancer cells. We analyzed the clinical impact and prognosis of WDFY2 expression on the progress of prostate cancer through data from online datasets. Our results showed that WDFY2 had lower expression level in prostate tumors than in normal tissues. Overexpression of WDFY2 in prostate cancer cells DU145 and PC-3 led to the suppression of cancer cell migration and colony formation. Furthermore, we found that WDFY2 exerted its role by suppressing the activity of Akt pathway other than the epithelial–mesenchymal transition progression. In conclusion, we have uncovered WDFY2 as a tumor suppressor gene and a new potential biomarker for cancer progression. Our results showed that WDFY2 inhibited cancer cell colony formation and migration via suppressing Akt pathway, making it a potential new therapeutic target in prostate cancer.
Introduction
Prostate cancer (PCa) is the cancer with the highest incidence in countries with the western-type living habits. It also has the second highest mortality among male cancer. 1 For now, we already have good diagnostic indexes on non-metastatic PCa, including serum prostate-specific antigen (PSA), clinical stage, and prostate biopsy Gleason score, which can provide a better clinical outcome. 2 However, metastatic PCa and castration-resistant prostate cancer (CRPC) are still the biggest challenges for clinical issues, which cause poor prognosis condition. So, it is urgently necessary to search for sensitive and specific biomarkers to monitor and diagnose PCa.
WD repeat and FYVE domain-containing protein 2 (WDFY2) has been reported as an adaptor-like protein to regulate protein phosphorylation and endocytosis. 3 In Caenorhabditis elegans, the silence of WDFY2 causes strong defects on coelomocyte endocytosis. 3 In mammalian cells, WDFY2 drives the maturation of endosomes from APPL endosomes to EEA1-positive endosomes. 4 Interestingly, a yeast two-hybrid screen showed that WDFY2 could interact with protein kinase Akt and protein kinase C.5,6 Perhaps WDFY2 could modulate the interaction between APPL and Akt downstream kinases. 4
Studying the molecular biomarkers will help to evaluate the progression of PCa and provide accurate therapy. Our study on the expression and function of WDFY2 in PCa may help to provide a new biomarker and further improve clinical outcome.
Materials and methods
Constructs
pPB-CAG-EBNXN vector was a kind gift from Sanger Institute. pPB-CAG-ires-Pac was generated as previously described.7,8 pPB-CAG-WDFY2-ires-Pac was generated by ligating full-length WDFY2 into the multiple-cloning sites (MCS) of pPB-CAG-ires-Pac.
Cell lines and cell culture
BPH-1, LNCaP, PC-3, and DU145 cells were kept under the condition of RPMI 1640 with 10% fetal bovine serum (FBS). Penicillin 100 U/mL, streptomycin 0.1 mg/mL, and amphotericin B 0.25 mg/mL were chosen to be an antibiotic–antimycotic solution added for all cells, which were put in standard culture conditions (37°C, 5% CO2, and 95% humidity). Neo and WDFY2 stable expressed cell lines were generated from co-transfection of pPB-CAG-WDFY2 and pCMVPBase in DU145 and PC-3 cells. After 2 µg/mL puromycin (AMRESCO) screening for 2 weeks, stable cell lines were selected and identified by western blotting.
Patient information
In this research, 90 patients with PCa were included for one group. Among them, 60 pairs of PCa tissues and their neighboring non-tumorous (NT) tissues were collected between 2011 and 2015. These tissue samples were processed by abruptly freezing in liquid nitrogen at once. The Clinical Research Ethics Committee of Huashan Hospital, Fudan University allowed the research protocols, and the participants agreed to give informed consents. Table 1 demonstrates the clinical information of PCa participants.
Clinical characteristics of 90 PCa patients and WDFY2 expression.
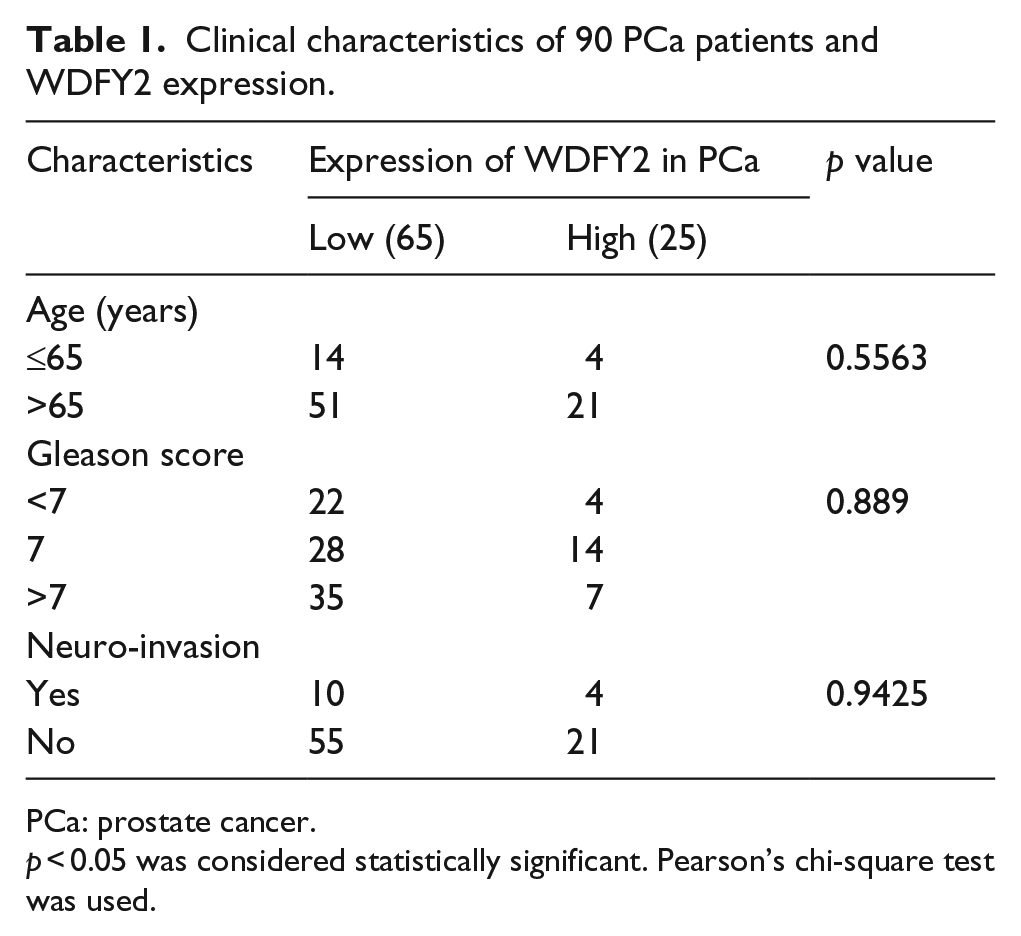
PCa: prostate cancer.
p < 0.05 was considered statistically significant. Pearson’s chi-square test was used.
Antibodies and immunoblotting
Cells were lysed using 1× sodium dodecyl sulfate (SDS) loading buffer (50 mM Tris-HCl pH 6.8, 10% glycerol, 2% SDS, 0.05% bromophenol blue, and 1% 2-mercaptoethanol). Some manufacturers were responsible for the production of antibodies, including anti-WDFY2 antibody (SAB), anti-E-cadherin (610181; BD Transduction Laboratories), anti-phospho-GSK3β, anti-Akt, anti-phospho-Akt (Ser473), anti-STAT3, anti-pSTAT3 (Tyr705), anti-P65, anti-pP65 (Ser536), anti-b-actin (Sigma-Aldrich), and also anti-tubulin (Proteintech). They were purchased from Cell Signaling Technology. For immunoblotting, proteins were isolated by SDS–polyacrylamide gel electrophoresis (PAGE) and removed toward polyvinylidene difluoride membranes (Millipore). Horseradish peroxidase (HRP)-conjugated secondary antibodies (from Jackson Laboratories) and increased chemiluminescence system were applied to signal detection.
Colony formation assay
Neo and WDFY2 stable DU145 and PC-3 cells could be trypsinized, calculated, and planted in the amount of 1000 cells per dish in 12-well culture dish (triplicate) and raised under the condition of 37°C and 5% CO2 for 7–10 days. Fresh growth media was changed for cells with a 3-day cycle. Colonies were fixed with 4% (w/v) paraformaldehyde for half an hour and stained with crystal violet (Sigma-Aldrich) for 20 min. After that, the photographs of the colonies were taken making use of EPSON Imager 600RGB. Colony quantification was performed by the Ipwin32 software.
Cell migration assays
Cells were trypsinized and suspended again under RPMI 1640 medium including 1% FBS at 3 × 105 cells/mL, and 0.2 cells/mL suspension had been spread to 8-µm Transwell inserts (Corning Costar). In the lower chambers, 0.8 mL of RPMI 1640 with 10% FBS was added. After 48-h period, a cotton swab could be used to remove the cells on the top surface of the membranes, and the fixing and crystal violet coloring of membranes could be achieved within 15 min.
Wound-healing assay
Cells spread within 12-well dishes with 1 × 105 cells density per well and cultivated into confluent type. Cell monolayers were removed by a sterile white micropipette, which resulted in a denuded area with a fixed width. Phosphate-buffered saline (PBS) was used to wash cells off cell debris, and then, culture medium was added to the cell culture. During the indicated period after being wounded, wound closure was monitored and photographed.
Real-time reverse transcription polymerase chain reaction
RNA extraction and reverse transcription polymerase chain reaction. TRIzol reagent (Invitrogen) was used to extract total RNAs from cells. Reverse transcription occurred on RNA. The reverse transcription was carried with reverse transcriptase (Fermentas) as per manufacturer’s instructions. Bio-Rad CFX96 system was used to perform quantitative reverse transcription polymerase chain reaction (qRT-PCR), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control to standardize the relative gene expression. Table 2 showed primer sequences for quantitative polymerase chain reaction (qPCR) probes of target genes.
Sequence of primers for qRT-PCR.
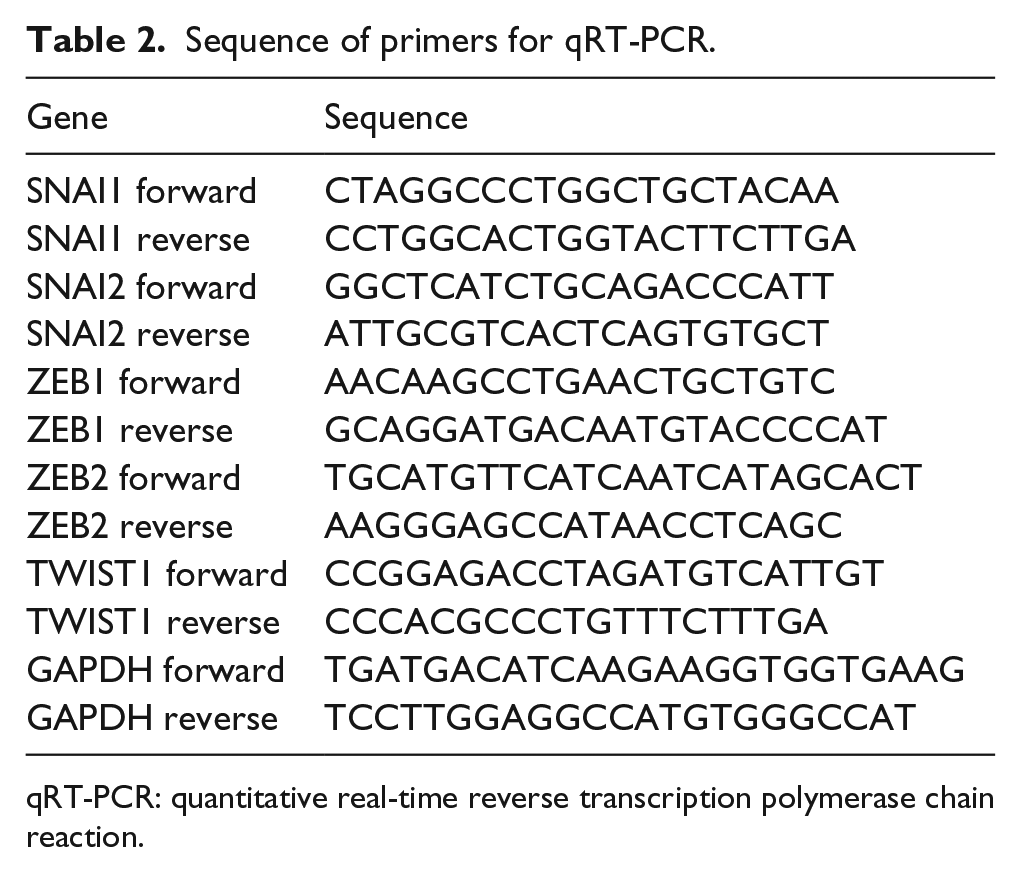
qRT-PCR: quantitative real-time reverse transcription polymerase chain reaction.
Tissue microarrays and immunohistochemistry
As described above, tissue microarrays (TMAs) were constructed, 9 and as described elsewhere, immunohistochemistry (IHC) was performed.10,11 WDFY2 staining could be simultaneously examined by two blinded, self-contained observers (with another pathologist); meanwhile, a consensus score was obtained for each score. A positive reaction for WDFY2 was scored in 4-grade categories based on the intensity of the staining, that is, 0, 1, 2 and 3, and the proportion of WDFY2-positive cells was scored in 4 groups as well: 0 (0%), 1 (1%–33%), 2 (34%–66%), and 3 (67%–100%). Under the circumstances of discrepancies between duplicated scores, the higher score of the two tissues was regarded as the final score. The final staining score was calculated based on the total amount of the intensity and percentage scores. And the pattern of staining could be sorted as follows: 0, negative; 1–2, weak; 3–4, moderate; 5– 6, strong. 12
Publicly available gene expression data sets and clinical data sets
In this article, we used expression data of WDFY2 in different human tissues and cancer cell lines from GTEx portal through The Human Protein Atlas portal (http://www.proteinatlas.org) 13 and the Cancer Cell Line Encyclopedia (CCLE). 14 All data were available directly online. We made use of some separate PCa clinical data sets that were publicly usable. These contained the data from ONCOMINE, 15 data reported in Taylor et al. 16 (GSE21032), a group of similar human prostate tumors with WDFY2 expression data, and a group of similar prostate adenocarcinoma samples at the cBioPortal for Cancer Genomics (The Cancer Genome Atlas (TCGA), Provisional). 17 Standardized gene expression intensity data applied in the research were indicated in Table S1.
Analyzing correlation between the expression of WDFY2 and risk of biochemical recurrence
For analyzing the correlation between the expression of WDFY2 and the risk of biochemical recurrence, we used an array of 484 primary prostate adenocarcinoma samples with WDFY2 expression data from TCGA collection, and the clinical variables were acquired from the cBioPortal for Cancer Genomics. 17 The expression level of WDFY2 was defined as Z-score. As a result, scores more than zero demonstrated higher expression level of WDFY2 in prostate tumor samples than prostate epithelial tissues. Meanwhile, scores less than zero demonstrated lower expression level in prostate tumor samples than prostate epithelial tissue. This study indicated the correlation between the expression level of WDFY2 and the risk of biochemical recurrence, which could reveal the stratification of prostate tumors.
Statistical analysis
The results are represented as the average values ± standard error of mean (SEM). The correlation between WDFY2 staining and the clinicopathological parameters of PCa patients was evaluated by a χ2 test. By the use of Kaplan–Meier method with log-rank test, overall survival rates could be actuarially calculated and evaluated from the day after surgery. Between-group variations were evaluated by the use of the χ2 test, Student’s t-test, the Mann–Whitney U test, and repeated-measures analysis of variance (ANOVA) test. Spearman’s correlation finished the exploration of the relationships. A p value below 0.05 was thought to have statistical significance.
Results
WDFY2 was expressed in human PCa and downregulated in PCa
To investigate the role of WDFY2 in PCa, we first confirmed whether WDFY2 was expressed in prostate. We analyzed the expression level of WDFY2 in different human tissues from the GTEx Portal (www.gtexportal.org). We found that WDFY2 was primarily expressed in skin and testis, and the expression level of WDFY2 in prostate was medium (Figure 1(a)). Meanwhile, we investigated the expression level of WDFY2 in different cancer cell lines through the Broad-Novartis Cancer Cell Line Encyclopedia (CCLE), which included genomic profiles of about 1000 cell lines for diverse tumor types. The results showed a relatively low expression of WDFY2 in PCa cell lines, which suggested that WDFY2 may be downregulated in PCa cells (Figure 1(b)).

Expression of WDFY2 in human prostate cancer. (a) Data from GTEx portal indicated that WDFY2 expressed in different tissues, including prostate (red arrow). (b) Expression of WDFY2 mRNA across various cancer cell lines. The highest expression level can be seen in meningioma cells, and its expression level in prostate cancer cell lines is relatively low (red arrow). The figure was analyzed using the data in Broad Cancer Cell Line Encyclopedia. (c) Gene alteration rate of WDFY2 in prostate cancer patients. (d) Functional plotting of the corresponding mRNA level and the genetic status of WDFY2 indicated that the mRNA level was consistent with gene change. (e) Data from Oncomine datasets showed that the mRNA level of WDFY2 is significantly downregulated in human prostate tumors. (f) WDFY2 mRNA level was downregulated in metastatic prostate cancer. The mRNA level of WDFY2 was exon-normalized as reported in GSE21032. Mann–Whitney U tests were performed to assess statistical significance for the comparisons between groups.
Next, we queried cases with available mutation and copy number variation (CNV) data in the TCGA PCa dataset using cBioPortal for Cancer Genomics. Deletion/mutation of WDFY2 was found in ~17% of cases (Figure 1(c)), among which deletion was the predominant type of alteration for WDFY2. Functional plotting of the corresponding messenger RNA (mRNA) level and the genetic status of WDFY2 revealed that the deletion of WDFY2 was associated with decreased mRNA expression (Figure 1(d)). To further confirm the mRNA expression level in PCa patients, we analyzed the RNA-seq data from Oncomine database, and the results indicated that WDFY2 was significantly downregulated in prostate tumor tissues (Figure 1(e)).
To assess WDFY2 expression level during the progression of PCa, we first analyzed the microarray data from GSE21032 and discovered that the mRNA level of WDFY2 in PCa was much lower than those of corresponding normal tissues (Figure 1(f) and Supplementary Table 1). Next, we tested the protein level of WDFY2 in multiple cell lines. Results revealed that WDFY2 was highly expressed in benign prostate cell line BPH-1 and was expressed relatively lower in weakly metastatic PCa cell line LNCaP. However, in aggressive, androgen-independent human PCa cell lines PC3 and DU145, the protein level of WDFY2 could barely be detected (p < 0.001, Figure 2(a)).

Associations between WDFY2 expression and prostate cancer risk and severity. (a) Western blot of WDFY2 in human prostate cell and prostate cancer cell lines. Tubulin served as the control. (b) Relative quantitation of WDFY2 protein levels in different prostate cancer cell lines. (c) Representative immunohistochemistry (IHC) of WDFY2 on benign prostatic epithelia (benign) and prostate cancer tissues. (d) Quantitation of protein expression scores of WDFY2 in benign prostatic epithelia and prostate cancer tissues. (e) Correlation between WDFY2 mRNA level and Gleason score of prostate tumor patients. mRNA level of WDFY2 was exon-normalized as reported in GSE21032. (f) Correlation between WDFY2 mRNA level and serum PSA level of prostate tumor patients. Mann–Whitney U tests were performed to assess statistical significance for the comparisons between groups in (e) and (f). (g) Kaplan–Meier plots examined the risk of biochemical recurrence in prostate tumor patients with higher or lower expression level of WDFY2 from the TCGA dataset. Note that the patients with tumors expressing higher levels of WDFY2 exhibit the decreased risk of biochemical recurrence. The number of patients was indicated in the x-axis. The p values were calculated by a log-rank test.
To further demonstrate the downregulation of WDFY2’s protein level in PCa, we tested WDFY2 expression by IHC in 90 PCa tissues and matched non-tumor tissues. After identifying PCa by hematoxylin–eosin (H&E) staining, immunoreactivity of WDFY2 protein was discovered in the cytoplasm, and the expression level of WDFY2 in PCa tissues was significantly lower than non-tumor tissues (Figure 2(c)).
Clinical impact of WDFY2 expression on the progression of PCa
To assess whether WDFY2 downregulation was related to the progression and prognosis of PCa, we analyzed the inter-relation between WDFY2 expression and clinicopathological characteristics among 90 participants with PCa. Table 1 shows the details. We also researched the results from two independent clinical data sets. In IHC assays, we found that the expression of WDFY2 was dramatically lower in PCa tissues compared with the benign tissues (Figure 2(c) and (d) and Table 3). Furthermore, prostate tumor samples with either high Gleason score or high PSA level showed the significantly lower level of WDFY2 (Figure 2(e) and (f)). More importantly, we found that patients with a lower WDFY2 level in tumor tissues showed much poorer prognosis (Figure 2(g)). All these results revealed that the low level of WDFY2 positively correlates with advanced stage, increased metastasis, and poor prognosis in PCa patients, which indicated the involvement of WDFY2 in the tumorigenesis and progression of PCa.
The expression of WDFY2 in prostate cancer tissue was significantly lower than that in normal tissue.

p < 0.05 was considered statistically significant. Pearson’s chi-square test was used.
p < 0.0001.
WDFY2 inhibited the colony formation ability and migration of PCa cells
In the previous research, we had observed the downregulation of WDFY2 in PCa samples including primary tumors and metastases, suggesting an unknown function of WDFY2 in the growth inhibition of prostate tumors. To test the biological relevance of WDFY2 expression and prostate tumor growth, we performed WDFY2 overexpression to study the cellular phenotypes. We used androgen-independent PCa cell lines, DU145 and PC-3, to establish the WDFY2 stably expressed cell lines. Western blotting showed overexpression of WDFY2 in stable cell lines (Figure 3(a)).

Overexpression of WDFY2 suppressed the colony formation and migration ability of prostate cancer cells. (a) Western blot showed the establishment of WDFY2 stably expressed cell lines in DU145 and PC-3. (b) Representative results of a migration assay for DU145 and PC-3 cells transfected with control vector or WDFY2 (scale bars = 100 µm). (c) Quantitation of the migration of DU145 and PC-3 cells transfected with control vector and WDFY2. (d) Results of wound-healing assay of the DU145 and PC-3 cells transfected with control vector and WDFY2. Each cell culture monolayers was scratched with a white micropipette tip, and then, the cell images were taken at the indicated time points. (e) Quantitation of migration ratio of DU145 and PC-3 cells transfected with control vector and WDFY2. (f) Results of colony-formation assay in DU145 and PC-3 cells transfected with control vector and WDFY2. (g) Quantitation of clones for DU145 and PC-3 cells transfected with control vector and WDFY2.
Then, we detected the migration ability and colony formation ability of WDFY2 overexpressed cancer cell lines to study its function. We researched the migration ability of cancer cells by Boyden chamber assay and quantified the cells. The results indicated a significant decrease in migration of WDFY2 overexpressed PC-3/DU145 cells compared to control cells in Transwell migration assays (Figure 3(b) and (c)). Meanwhile, to further confirm whether the overexpression of WDFY2 could inhibit the migration of PCa cells, we also performed wound-healing assays on these cell lines. Results showed that wound healing was also inhibited by WDFY2 overexpression (Figure 3(d) and (e)), which indicated that WDFY2 overexpression could inhibit the migration ability of PCa cells. Then, colony formation assay was also performed on these cell lines. After 7-day observation, we found that the clonogenic ability of PCa cells was suppressed in WDFY2 overexpressed cell lines, including clone number and volume (Figure 3(f) and (g)). All the results mentioned above supported that WDFY2 could inhibit the migration and colony formation ability of PCa cells.
We also detected the proliferation of these cell lines. Results indicated that WDFY2 overexpression had no influence on the proliferation ability of PCa cells (data not shown).
WDFY2 selectively affected Akt pathway other than the epithelial–mesenchymal transition
As the results showed above, we have confirmed the function of WDFY2 in prostate progression. As many previous studies indicated, epithelial–mesenchymal transition (EMT) was the key contributor in the metastatic process. EMT played a crucial role on tumor cells acquiring migratory and invasive characteristics. The reduction of E-cadherin is an important hallmark of EMT, as E-cadherin mediates the early contact of epithelial cells. 18 To investigate whether WDFY2 suppresses the migration of PCa cell by induction of EMT, we tested the epithelial marker E-cadherin expression. The results showed that WDFY2 overexpression in DU145 did not induce or suppress the expression of E-cadherin (Figure 4(a)). To further determine whether WDFY2 influenced EMT, we checked whether essential transcription factors in EMT were regulated by WDFY2. The zinc-finger-containing proteins Snail and Slug, zinc finger E-box–binding (ZEB) proteins ZEB1 and ZEB2, and the helix–loop–helix transcription factor TWIST were reported to repress the expression of E-cadherin and led to EMT.18,19 Consistent with the previous results, we found that WDFY2 overexpression in DU145 showed no major difference in the mRNA level of the EMT transcription factors mentioned above (Figure 4(b) and (c)). These results indicated that WDFY2 did not regulate PCa cell migration through EMT.

WDFY2 influenced prostate cancer development via Akt pathway specifically. (a) and (b) Western blotting showed that overexpressing WDFY2 did not affect the expression of E-cadherin and SLUG. (c) Real-time RT-PCR analysis of the mRNA level of Snail, Slug, Twist, ZEB1, and ZEB2 in the DU145 cells transfected with control vector and WDFY2. (d) and (e) WDFY2 overexpression showed specific downregulation of pAkt (Ser473). The values were the mean ± SEM of three independent experiments, and the p value is shown from Student’s t-test analysis.
Many cell signaling pathways were reported to regulate colony formation and migration ability of cancer cells, such as phosphoinositide 3-kinase (PI3K)/Akt pathway, nuclear factor–κB (NF-κB) pathway, and signal transducer and activator of transcription 3 (STAT3) pathway. Previous studies had shown that WDFY2 modulated Akt interactions with its substrates and altered the PI3K/Akt pathway in ovarian carcinomas.20–22 Hence, we tested whether WDFY2 also impaired PI3K/Akt pathway in PCa cell line DU145. By western blotting, we found that WDFY2 overexpression restrained the phosphorylation of Akt at Ser473, while the protein level of total Akt and pGSK-3β remained unchanged. Moreover, the activity of mechanistic target of rapamycin (mTOR) revealed by phosphorylation of S6 showed no significant alteration, and the NF-κB and STAT3 pathways were not influenced either. These results indicated that WDFY2 specifically inhibited Akt pathway in PCa, which could contribute to the suppression of migration in PCa cells (Figure 4(c) and (d)).
Discussion
In this study, we first discovered the downregulation of WDFY2 during the progression of PCa and that PCa patients with lower WDFY2 level showed poorer prognosis. We further uncovered the molecular mechanism that WDFY2 could inhibit the colony formation and migration ability of PCa cells via suppressing Akt pathway specifically. These results suggest WDFY2 as a tumor suppressor and a potential new biomarker of PCa.
Kannan et al. reported that in many tumor samples especially high-grade ovarian carcinoma, the gene fusion of CDKN2D-WDFY2 led to the loss of full-length WDFY2 and the expression of shortened isoform of WDFY2. This short isoform could impair Akt interacting with its substrates. 20 Our results also showed that in PCa cells, WDFY2 repressed Akt pathway specifically, which may suggest a common function of WDFY2 related to Akt pathway in tumors. Further study can be done to test this hypothesis in multiple cancer cells.
Till today, there was no curative treatment for metastatic PCa. This research revealed that overexpression of WDFY2 in PCa cells inhibited their migration and colony formation ability dramatically. It provides us a new thought to prevent the metastasis of PCa through drug activation of WDFY2.
Several oncogenic signaling pathways, for example, transforming growth factor (TGF)-β, Notch, NF-κB, STAT3, and PI3K/Akt, have been confirmed to modulate cancer cell migration. 15 Among them, active Akt promotes cell migration through inducing EMT 23 or other mechanisms in various cancer cells. Previous studies have shown that endosomal protein WDFY2 could regulate AKT2 and that WDFY2 may play a role in the spatial organization of signaling cascades in Akt signaling pathway. 5 To dig out the mechanisms and precise features of WDFY2 for cancer progression and metastasis, we detected the key pathways above. Results showed us that the expression of AKT2 did not change after WDFY2 overexpression (data not shown). However, Akt activity was decreased in WDFY2 overexpressed DU145 cells, while other signaling pathways were not influenced. Thus, we revealed the function and initial mechanisms of WDFY2 in PCa, which would offer many in-depth ideas for the mechanisms of WDFY2 in oncogenesis and cancer progression.
PCa remains the leading cause of death in the world. At present, there have not been reliable markers of PCa for early detection, diagnostic confirmation, and good disease prognosis. 21 Hence, it is important to identify PCa biomarkers for accurate diagnosis and stratification of patients. This study found that WDFY2 was downregulated in the progression of PCa, coupled with bad prognosis, and that WDFY2 inhibited the invasion and metastasis of cancer cells by suppressing the activity of Akt pathway. All these results indicate that WDFY2 may serve as a tumor suppressor gene. Hence, WDFY2 is expected to become a good diagnostic and therapeutic target to control PCa metastasis.
Footnotes
Acknowledgements
The authors thank Professor Yujiang Geno Shi for the help in providing general support.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Key Project of the National Natural Science Foundation of China (grant no. 81130047), National Key Basic Research Program, Scientific Research Project of Huashan Hospital (grant no. 2013QD12), Youth Elites Scientific Funding of Shanghai Municipal Health Bureau (grant no. 20144Y0111), General Program Scientific Funding of Shanghai Municipal Health Bureau (grant no. 201440557).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.