
Abstract
Human renal cell carcinoma which is a highly vascular tumor is the leading cause of death from urologic cancers. Angiogenesis has a pivotal role in oncogenesis and in the viability and expansion of renal cell carcinoma. Rap2B, as a small guanosine triphosphate–binding protein of the Ras family, was first discovered in the early 1990s during the screening of a platelet complementary DNA library. Previous studies have shown that Rap2B aberrantly expressed in human carcinogenesis and promoted the development of tumors via multiple signaling pathways. However, the function of Rap2B in tumor angiogenesis that is necessary for tumor growth and metastasis remains unknown. In this study, we examined the role of Rap2B in angiogenesis in renal cell carcinoma by Western blot, quantitative polymerase chain reaction, enzyme-linked immunosorbent assay, human umbilical vascular endothelial cells growth assay, and endothelial cell tube formation assay. We found that Rap2B promoted angiogenesis in vitro and in vivo. Moreover, our data illustrated that phosphoinositide 3-kinase/AKT signaling pathway is involved in Rap2B-mediated upregulation of vascular endothelial growth factor and renal cell carcinoma angiogenesis. Taken together, these results revealed that Rap2B promotes renal cell carcinoma angiogenesis via phosphoinositide 3-kinase/AKT/vascular endothelial growth factor signaling pathway, which suggests that Rap2B is a novel therapeutic target for renal cell carcinoma anti-angiogenesis therapy.
Keywords
Introduction
Human renal cell carcinoma (RCC) is probably the most common cause of cancer deaths in humans affected by urologic tumors, 1 and the incidence of RCC is increasing by a rate of approximately 2.5% each year. 2 RCC is characterized by poor prognosis and the survival rate is only 25% after diagnosis of the metastatic disease. 3 RCC originates from the proximal tubule cells of nephrons and is a highly vascular tumor. 4 Therefore, metastasis is the primary mediator of mortality for this disease, which remains a challenging clinical problem. 5 Tumor cells’ ability to produce new blood supply has great effects on its growth and metastasis. Angiogenesis is a fundamental early step for human cancer progression and development. 6 Vascular endothelial growth factor (VEGF) is a pro-angiogenesis factor and has been shown that its expression was mediated via phosphoinositide 3-kinase (PI3K)/AKT signaling pathway. 7 Ping et al. 8 showed that PI3K/AKT signaling pathway could induce the expression of VEGF in glioblastoma. Newly developed targeting angiogenesis therapies are the standard of care in metastatic RCC, 9 but the acquisition of resistance often occurs during the treatment. Herein, any investigation of the RCC therapy with the molecular mechanisms underlying angiogenesis and progression is required so as to develop more effective and specific therapies for patients with advanced RCC.
Rap proteins or Ras-related guanosine triphosphate (GTP)-binding proteins are a subset of Ras superfamily of small GTP-binding proteins and consist of five members, Rap1A, Rap1B, Rap2A, Rap2B, and Rap2C. 10 Previous studies have suggested that the members of Ras family are involved in regulation of a great diversity of biological functions, including apoptosis, signal transduction, proliferation, as well as migration in human cells.11,12 A recent study showed that Rap1 promotes VEGF receptor 2 (VEGFR2) activation and angiogenesis through involving integrin αvβ3. 13 More recently, a large number of articles illuminate the Rap2 biological effects. Although the effector region varies from that of Rap1 proteins by only one residue, 14 there is no literature about the unique function of Rap2 in tumor angiogenesis so far.
Rap2B was first discovered in the early 1990s during the screening of a platelet complementary DNA (cDNA) library.15,16 Its identity is approximate 50% with classical Ras proteins. There are three well-defined regions in Rap2B being responsible for subcellular localization: 17 an effector domain (amino acids 32–40), a nucleotide binding domain, and C terminal tetra peptide motif Cys-Val-Ile-Leu (CVIL). 18 Previous studies have shown the evidences that Rap2B presents a high level of expression in human carcinogenesis.19,20 Our recent studies have shown that Rap2B could regulate the cytoskeleton in a p53-dependent fashion. 21 Besides, our research shows that aberrant expression of Rap2B contributed to proliferation and accelerated migration as well as invasion of human breast cancer via mediating calcium-related extracellular signal–regulated protein kinases 1 and 2 (ERK1/2) signaling pathway. 22 More recently, we demonstrated that prostate cancer (PCa) growth and metastasis are positively regulated by Rap2B, as well as the expression of phosphorylated focal adhesion kinase (FAK). 23 Xie et al. 24 also demonstrated that miR-342-3p inhibited proliferation and invasion of non–small cell lung cancer through suppressing Rap2B. In addition, Rap2B is also a novel p53 target and participates in p53-mediated pro-survival function under DNA damage condition. 25
In this study, we try to investigate the role of Rap2B in RCC angiogenesis. Our researchers illuminated that Rap2B can promote RCC growth in vivo. Furthermore, we found that Rap2B stimulated endothelial cells proliferation and angiogenesis. Moreover, we first indicated that PI3K/AKT signaling pathway is involved in Rap2B-mediated upregulation of VEGF and RCC angiogenesis in vitro as well as in vivo, respectively. Our data may provide a potential therapeutic target for this aggressive human RCC.
Materials and methods
Cell culture and stable cell line generation
Human RCC lines (786-O, ACHN) were purchased from the Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences (Shanghai, China). Human umbilical vascular endothelial cells (HUVECs) were obtained from KeyGen Biotech (Nanjing, China). 786-O and HUVECs cells were grown in RPMI-1640 medium, ACHN cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) (Invitrogen, Shanghai, China). All these media were supplemented with 10% fetal calf serum (Invitrogen, Shanghai, China), and all these cells were incubated in a 37°C humidified incubator with 5% CO2. The control 786-O cell lines (Ctrl-786-O), Rap2B-overexpressed 786-O cell lines (Rap2BOE-786-O), and Rap2B-knockdown 786-O cell lines (Rap2BKD-786-O) were constructed by infecting with lentivirus packing control vector, Rap2B expression vector (enhanced green fluorescent protein (EGFP) is not fused with Rap2B in this vector), and Rap2B short hairpin RNA (shRNA) expression vector, respectively (GenePharma, Shanghai, China), for 48 h and then selected with puromycin (Santa Cruz Biotechnology, Shanghai, China) for 3 weeks.
Preparation of conditioned medium
In the series of experiments, stable cell lines Rap2BOE were pre-treated with either control diluent or pharmacological inhibitors, including AKT inhibitor MK2206 (Selleck Cat. No. S1078) and PI3K inhibitor LY294002 (Selleck Cat. No. S1105) for 24 h, respectively. Then, the cells were washed and replaced with serum-free medium. Conditioned medium (CM) was then collected 24 h after the change of medium and stored at −80°C until use.
Western blot analysis
Western blot analysis was carried out as described previously. 22 All protein samples were separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and electro-transferred onto nitrocellulose (NC) filter membrane. The membrane was blocked for 2 h in Blotto (5% non-fat milk and phosphate-buffered saline (PBS)) and then incubated overnight at 4°C with the following primary antibodies: anti-Rap2B (AB1369, Abcam, Shanghai, China), anti-PI3K(#4249, Cell Signaling Technology, Shanghai, China), anti-AKT (#9272, Cell Signaling Technology, Shanghai, China), anti-p-AKT (#4060, Cell Signaling Technology, Shanghai, China), and anti-β-actin (Zhongshan Biotech, Beijing, China). After washing, the membranes were incubated with secondary antibodies conjugated with IRDye 680 or IRDye 800 (Rockland Immunochemicals Inc., Limerick, PA) at 37°C for 2 h. The corresponding bands were imaged with Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE).
Enzyme-linked immunosorbent assay for VEGF
786-O and ACHN cells were plated on six-well tissue culture plates at a density of 1 × 106 cells/well. Then, the supernatants were collected after 24 h. VEGF concentration was determined using Quantikine ELISA Kits according to the manufacturer’s instructions (Westang Biotech, Shanghai, China).
HUVECs growth assay
The HUVECs growth was assayed using Cell Counting Kit-8 (CCK-8) purchased from Beyotime Institute of Biotechnology (Nanjing, China). 26 In brief, 6 × 103 HUVECs were suspended in 100 µL CM obtained from 786-O and ACHN cells in a 96-well culture plate, respectively, and incubated at 37°C in a humidified atmosphere containing 5% CO2 for 24 h. After which, cell proliferation was detected according to the manufacturer’s instructions.
Endothelial cell tube formation assay
786-O and ACHN cells (1 × 106) were cultured on six-well plates with fresh complete medium. After 24 h, the medium was collected and centrifuged to remove any cell debris before its use as a CM. The 48-well plates was coated with matrigel and kept at 37°C for 30 min. Then, 4 × 104 HUVECs were suspended in 100 µL CM, planted onto the pre-coated 48-well plates and incubated at 37°C. After 10 h of incubation, the number of capillary-like tubes from four randomly chosen fields was counted.
Animals and angiogenesis assay in vivo
Five-week-old male BALB/c nude mice were purchased from the Shanghai Laboratory Animal Center (Shanghai, China) for studies approved by the Animal Care Committee of Xuzhou Medical University. For RCC proliferation assays in vivo, the BALB/c nude mice were randomly divided into three groups consisting of eight mice each. Ctrl-ACHN, Rap2BOE-ACHN, Rap2BKD-ACHN (4 × 106) cells were suspended in 200 µL medium containing 100 µL PBS and 100 µL matrigel and injected into flank sites of mice, respectively. Tumor growth was monitored. 48 days later, three groups of mice were sacrificed and their subcutaneous tumors were dissected. For angiogenesis assays in vivo, the BALB/c nude mice were randomly divided into five groups consisting of four mice each. Stable cell lines Rap2BOE-ACHN, Rap2BKD-ACHN, and Ctrl-ACHN (4 × 106) cells were suspended in 200 µL medium containing 100 µL PBS and 100 µL matrigel, and injected into flanks of three groups, respectively. In addition, other two groups were injected with Rap2BOE-ACHN (2 × 106) cells, and those mice were treated with either MK2206 by oral gavaging (120 mg/kg on day 1, day 3, and day 5) or LY294002 intraperitoneally (i.p., 50 mg/kg on day 2 and day 5). After seven days, the five groups of mice were sacrificed and their tumor were removed and fixed in 10% buffered formalin for further angiogenesis analysis by CD31 immunohistochemistry. The histopathological analysis of each set of tumor angiogenesis was visually inspected by means of a stereoscopic dissecting microscope.
CD31 immunohistochemistry
Based on the streptavidin–peroxidase (SP) method, a standard Sp Kit (Zhongshan Biotech) was utilized. Tissue microarray (TMA) slides were dewaxed at 65°C for 4 h followed by two 30-min washes with xylene and then rehydrated with graded ethanol and distilled water. Antigen retrieval was carried out by heating the sections in 10 mM citrate buffer (pH 6.0) at 95°C for 30 min, and endogenous peroxidases were interdicted by 3% hydrogen peroxide for 30 min. After 30-min blocking with 5% normal goat serum, the sections were incubated overnight at 4°C with polyclonal rabbit anti-CD31 antibody (#28364, Abcam). Then, the slides were incubated with a biotinylated secondary antibody (Zhongshan Biotech) for 30 min at room temperature, followed by avidin–peroxidase reagent for an additional 30 min and 3,3′-diaminobenzidine (DAB, Zhongshan Biotech) substrate for 5 s. The sections were sealed with coverslips after hematoxylin counterstain and dehydration.
Real-time polymerase chain reaction
A two-step real-time reverse transcription polymerase chain reaction (RT-PCR) procedure was used. All the reagents were purchased from TaKaRa (Shanghai, China), and all methods were followed according to the manufacturer’s instructions. Sequences for all target gene primers (glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as internal control) were purchased commercially (Applied Biosystems, Foster City, CA). The total RNA was isolated from the cells using RNAiso Plus (cat# 9108; TaKaRa). For micro RNA (miRNA) assay, cDNA was synthesized from total RNA by PrimeScript™ RT Reagent Kit (cat# RR037A; TaKaRa). SYBR Green reagents (cat# RR420A; TaKaRa) were used for the 40-cycle real-time PCR, and the reactions were performed using an Applied Biosystems 7500 Real-Time PCR machine (Applied Biosystems). For quantification, GAPDH was used to normalize the levels of messenger RNA (mRNA) using the relative quantification method of analysis. The mRNA levels of target genes from virus-treated cells were expressed as the percentage of expression relative to that of the control cells.
Statistical analysis
Quantitative data are expressed as mean ± standard deviation (SD). Statistical analysis in treatment groups was evaluated by Student’s t tests. All experiments were carried out at least three times unless otherwise indicated. *p < 0.05, **p < 0.01, and ***p < 0.001 were considered as statistically significant difference.
Results
Rap2B promotes tumor growth in vivo
To investigate the diverse roles of Rap2B in RCC, Rap2BOE-786-O, Rap2BKD-786-O, and Ctrl-786-O cells were established. Likewise, the three stable cell lines of ACHN were also constructed. After 3-week selection followed by lentivirus infection, the Rap2B protein levels in these cells were determined by Western blot (Figure 1(a)). To further address the role of Rap2B in RCC growth, Rap2BOE-ACHN, Rap2BKD-ACHN, and Ctrl-ACHN cells were injected through subcutis into nude mice and then tumor growth was monitored. After 48 days, three groups of mice were sacrificed and their subcutaneous tumors were dissected (Figure 1(b)). The result was consistent with our previous study indicating that Rap2B could promote PCa growth. 23 The weight of Rap2B-overexpressed tumors was increased; however, Rap2B-deleted tumors were much smaller, compared with controls (Figure 1(c)). As shown in Figure 1(d), tumors in Rap2BOE group grew at a quicker rate and had larger sizes compared with those in controls, while tumors in mice implanted with Rap2BKD-ACHN cells had sustained a significant growth arrest. The 786-O cells were also injected into nude mice, but there is no tumor formation, which is consistent with previous studies. 27

Rap2B promotes RCC growth in vivo. (a) Western blot of Rap2B expression level in Rap2BOE-786-O/ACHN, Rap2BKD-786-O/ACHN, and Ctrl-786-O/ACHN stable cell lines. (b) Nude mice were subcutaneously implanted with 4 × 106 Rap2BOE, Rap2BKD, and Ctrl-ACHN cells lines. After 48 days, the three groups of mice were sacrificed and their subcutaneous tumors were dissected. (c) The tumor diameters were measured, and the tumor volumes were calculated. (d) The xenograft tumors were collected and weighed. Data are presented as mean ± SD (n = 8).
Rap2B upregulates PI3K mRNA levels
Since cumulative evidence suggests that angiogenesis, a hallmark of cancer development, plays a necessary role in tumor growth, we were interested in addressing whether Rap2B plays an important role in angiogenesis in RCC. PI3K has shown potential capacity for mediating tumor angiogenesis. Thus, quantitative PCR (qPCR) assay was utilized to determine Rap2B and PI3K mRNA level in RCC. As shown in Figure 2(a)–(d), both Rap2BOE-ACHN and Rap2BOE-786-O have higher levels of Rap2B and PI3K mRNA, compared with those in control-infected stable cells. Nevertheless, silencing of Rap2B lead to a decrease in levels of PI3K mRNA, compared with those in control-infected stable cells. Previous study has implicated that PI3K might be involved in gene expression regulation through its downstream transcriptional factors 28 and AKT is the most important downstream effector of PI3K. 29 Herein, we next use PI3K inhibitor LY294002 or AKT inhibitor MK2206 to restrain the function of PI3K and examined the expression of Rap2B in Rap2BOE-ACHN and Rap2BOE-786-O cells. Unexpectedly, the results in Figure 2(a) and (b) have shown that the levels of Rap2B mRNA was not affected in the presence of LY294002 or MK2206, while the levels PI3K mRNA were decreased by LY294002 in Rap2BOE-ACHN and Rap2BOE-786-O cells, indicating that PI3K signaling pathway is critical for its own expression but not for Rap2B regulation. However, Rap2B could stimulate the expression of PI3K mRNA.

Rap2B can upregulate PI3K mRNA level. (a and b) Rap2BOE-ACHN/786-O cell lines were incubated with LY294002 (2 µM) or MK2206 (50 µM) for 24 h or Ctrl-ACHN/786-O, Rap2BKD-ACHN/786-O, and Rap2BOE-ACHN/786-O cell lines were cultured for 24 h. Rap2B mRNA expression was rated by qPCR (n = 3). (c and d) Rap2BOE-ACHN/786-O cell lines were incubated with LY294002 (2 µM) for 24 h or Ctrl-ACHN/786-O, Rap2BKD-ACHN/786-O, and Rap2BOE-ACHN/786-O cell lines were cultured for 24 h. PI3K mRNA expression was rated by qPCR (n = 3).
Rap2B promotes PI3K and AKT phosphorylation
Previous studies have implicated that PI3k/AKT signaling pathway is one of the most important pathways contributed to tumor angiogenesis.30,31 Thus, we carried out Western blot to determine whether Rap2B affects the phosphorylated forms of AKT in those stable cell lines Ctrl, Rap2BKD, and Rap2BOE in RCC. In addition, Rap2B upregulated the levels of PI3K mRNA as shown above, which was also confirmed by Western blot. In Figure 3(a) and (b), results revealed that the expression of PI3K and p-AKT was dramatically increased after forced-expression of Rap2B in ACHN and 786-O cells. As expected, knocking down of Rap2B decreased the levels of PI3K and p-AKT in both the RCC cell lines, whereas the total protein levels of AKT were not affected. Next, to further confirm the effects of PI3k/AKT pathway on Rap2B-mediated signaling, we treated these Rap2B-overexpressed cells with LY294002 or MK2206 for 24 h. The upregulation of p-AKT induced by Rap2B overexpression was attenuated by MK2206 in both the cell lines. However, Rap2B, PI3K, and total AKT were not affected after treatment with MK2206. Meanwhile, forced-expression of Rap2B-mediated upregulation of PI3K and p-AKT was decreased after administration of LY294002 in both the cell lines (Figure 3(a) and (b)). Above all, these results indicated that Rap2B was involved in the activation of PI3K/AKT signaling pathway.
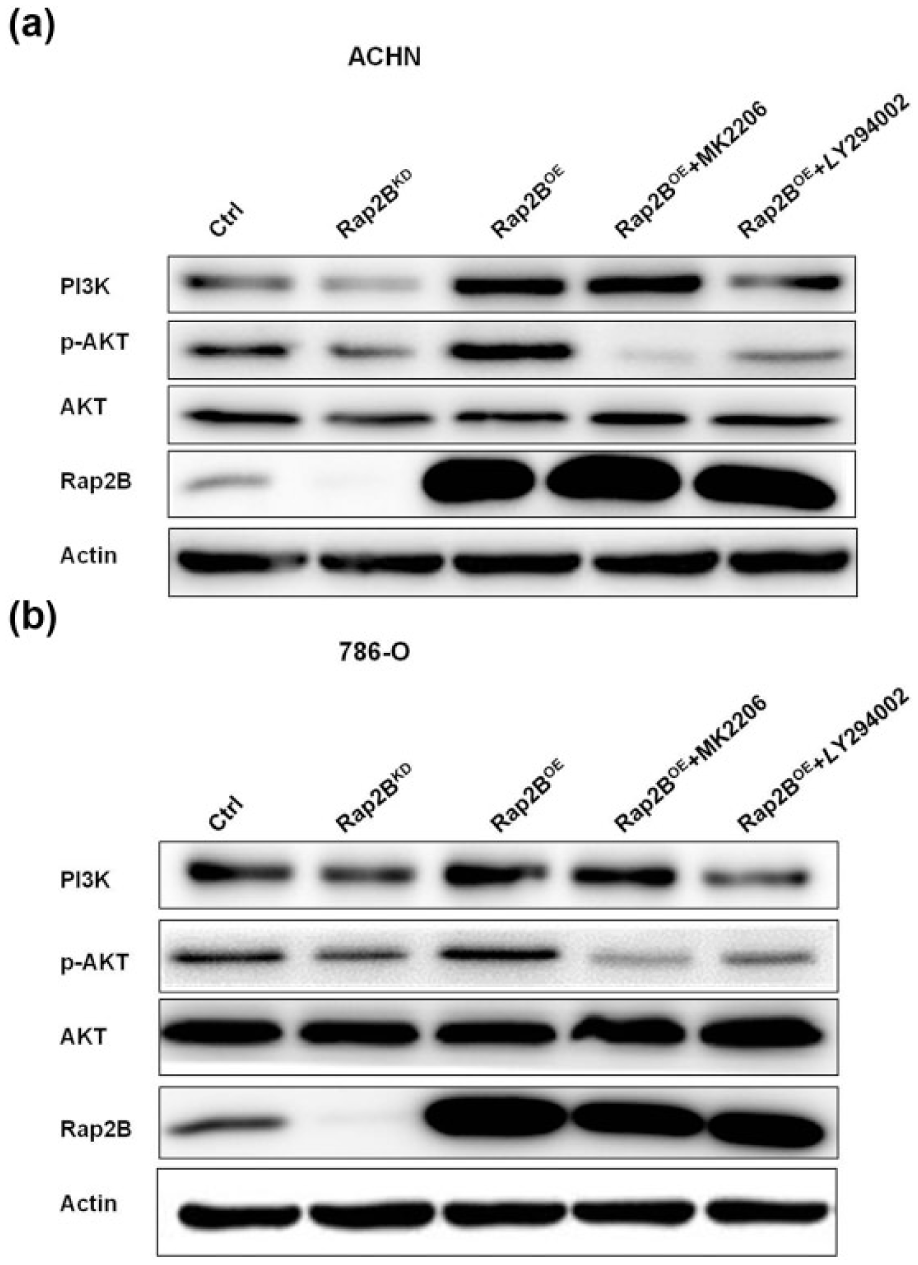
Rap2B can upregulate PI3K and AKT phosphorylation. (a and b) Stable Rap2BOE-ACHN/786-O cell lines were incubated in the AKT inhibitor MK2206 (50 µM) and PI3K inhibitor LY294002 (2 µM) for 24 h. Western blot analysis of the protein levels of Rap2B, p-AKT, and PI3K in Ctrl-ACHN/786-O, Rap2BKD-ACHN/786-O, Rap2BOE-ACHN/786-O, Rap2BOE+MK2206-ACHN/786-O, and Rap2BOE+LY294002-ACHN/786-O cell lines.
Rap2B induced upregulation of VEGF mRNA and VEGF secretion through PI3K/AKT pathway
Studies have shown that activated PI3K/AKT could upregulate the expression of VEGF, which is a 45 kDa glycoprotein secreted by various cells, including endothelial cells, podocytes, macrophages, fibroblasts, and malignancies. 32 VEGF plays an indispensable role in tumor angiogenesis. Therefore, we investigated whether Rap2B affected the expression of VEGF mRNA level by qPCR assay (Figure 4(a) and (b)). As expected, the mRNA levels of VEGF were significantly increased after Rap2B overexpression and also modestly decreased when Rap2B was downregulated in these cells. In addition, we also determined the secreted levels of VEGF in Rap2BOE, Rap2BKD, Ctrl, Rap2BOE+MK2206, and Rap2BOE+LY294002-ACHN/786-O CM by enzyme-linked immunosorbent assay (ELISA). The upregulated levels of secreted VEGF were suppressed by MK2206 and LY294002 in both the cell lines, indicating that Rap2B can upregulate VEGF mRNA level through PI3K/AKT pathway.

Rap2B can upregulate VEGF mRNA level and increase VEGF secretion through PI3K/AKT pathway. (a and b) Rap2BOE-ACHN/786-O cell lines were incubated with LY294002 (2 µM)/MK2206 (50 µM) for 24 h or Ctrl-ACHN/786-O, Rap2BKD-ACHN/786-O, and Rap2BOE-ACHN/786-O cell lines were cultured for 24 h. VEGF mRNA expression rated by qPCR (n = 3). (c and d) Stable Rap2BOE-ACHN/786-O cell lines were incubated in the AKT inhibitor MK2206 (50 µM) or PI3K inhibitor LY294002 (2 µM) for 24 h. The conditioned mediums of Ctrl-ACHN/786-O, Rap2BKD-ACHN/786-O, Rap2BOE-ACHN/786-O, Rap2BOE+MK2206-ACHN/786-O, and Rap2BOE+LY294002-ACHN/786-O cell lines were collected and the secreted level of VEGF was determined by ELISA. VEGF secretion level was positively regulated by Rap2B through PI3K/AKT in both ACHN and 786-O cells. Data are presented as mean ± SD (n = 5).
Rap2B promotes HUVECs proliferation and angiogenesis through PI3K/AKT pathway in vitro
VEGF could accelerate vascular endothelial cell proliferation and the angiogenesis ability.33,34 Therefore, we sought to investigate whether Rap2B had an influence on endothelial cell proliferation and angiogenesis, which is crucial for new vessel development. HUVECs growth assay was performed utilizing the collected CM containing secreted cytokines from Rap2B overexpression, knockdown, Ctrl, or Rap2B overexpression group treated with PI3K or AKT inhibitors in RCC cells. As we expected, all the data showed that Rap2BOE-ACHN/786-O CM significantly augmented the ability of HUVECs to grow, while HUVECs suspended in Rap2BKD-ACHN/786-O CM had a significant growth arrest compared with that in control-infected CM. Besides, Rap2BOE+MK2206-ACHN/786-O CM and Rap2BOE+LY294002-ACHN/786-O CM have no effect of HUVECs proliferation compared with Rap2BOE-ACHN/786-O CM (Figure 5(a) and (b)). To further confirm the function of Rap2B in regulating angiogenesis in RCC, tube formation assays were performed. The average number of complete tubular structures formed by HUVECs suspended in Rap2B-overexpressed CM was increased compared with that of HUVECs suspended in control CM (Figure 5(d) and (f)). However, the number was significantly decreased in HUVECs suspended in Rap2B-knockdown CM (Figure 5(d) and (f)). Moreover, we found that both MK2206 and LY294002 could inhibit the tube formation ability mediated by forced-expression of Rap2B (Figure 5(d) and (f)).

Rap2B promotes HUVECs proliferation and angiogenesis through PI3K/AKT pathway in vitro. (a and b) HUVEC growth assay was performed to detect the HUVECs proliferation. Rap2B overexpression significantly promoted proliferation of HUVECs. Nevertheless, Rap2B knockdown significantly inhibited proliferation of HUVECs. MK2206 and LY294002 could inhibit the HUVECs proliferation ability accelerated by Rap2B overexpression (n = 3). (c–f) Representative pictures were taken in situ for tube formation in the CM of 786-O and ACHN cells. The degree of tube formation was assessed as the number of tubes. The result was in accordance with HUVEC growth assays (n = 3). Data are shown as mean ± SD.
Rap2B promotes angiogenesis through PI3K/AKT pathway in vivo
Next, we investigated whether and how Rap2B could promote tumor angiogenesis in vivo. To generate experimental metastasis, the BALB/c nude mice were randomly divided into five groups consisting of four mice each. Stable cell lines Rap2BOE-ACHN, Rap2BKD-ACHN, and Ctrl-ACHN were mixed with the same volume of matrigel and injected into flanks of mice in three groups. In addition, other two groups were injected with Rap2BOE-ACHN (2 × 106) cells, and those mice were treated with either MK2206 by oral gavaging (120 mg/kg on day 1, day 3, and day 5) or LY294002 i.p. (50 mg/kg on day 2 and day 5). After 7 days, the five groups of mice were sacrificed and the tumors were resected (Figure 6(a)). We then conducted immunohistochemical analysis of CD31, a marker of endothelial cells, on tumors to investigate the underlying mechanisms of Rap2B on angiogenesis. As shown in Figure 6(b), the relative expression of CD31-positive cells in Rap2BOE-ACHN group was much higher compared with control-infected ACHN cells, while the expression of CD31-positive cells in Rap2BKD-ACHN group was lower compared to control-infected ACHN cells. Besides, inhibition of PI3K/AKT pathway by either MK2206 or LY294002 decreased the levels of CD31-positive blood vessels expressed in Rap2BOE-ACHN group (Figure 6(b)). Consistent with previous study, 786-O cells cannot form tumors in vivo. These results are in an agreement with the effects of Rap2B in vitro and further validated that Rap2B functions as a tumor angiogenesis promoter in RCC, possibly attributable to PI3K/AKT signaling pathway.

Rap2B can promote angiogenesis through PI3K/AKT pathway in vivo. (a) Representative images of 10% buffered formalin-fixed tumors resected after 7 days of intravenous implantation with 4 × 106 different treatments of ACHN cells. In Rap2BOE+MK2206 group, MK2206 was administered by oral gavaging (120 mg/kg on day 1, day 3, and day 5). In Rap2BOE+LY294002 group, LY294002 was administered intraperitoneally (i.p., 50 mg/kg on day 2 and day 5). (b) Immunostaining of CD31 in tumors of the five groups.
Discussion
Rap2B gene is located at 3q25.2 of human chromosome and was first discovered in the early 1990s during the screening of a platelet cDNA library.35,36 Rap2B is a subset of Ras superfamily of small GTP-binding proteins, and it is expressed in high level in human carcinogenesis.19,20 Recently, many groups pay attention to the role that Rap2B regulates tumor migration and invasion abilities. A large number of evidences indicated that mutations or overexpression of Ras gene was observed and associated with poor prognostic survival in various human tumors. 37 In our previous study, we found that overexpression of Rap2B could accelerate proliferation, migration, and invasion of breast cancer as well as PCa via upregulating calcium-related ERK1/2 and FAK-dependent pathway, respectively. 22 Moreover, Rap2B as a novel p53 target participates in Ca2+-dependent FAK–ERK1/2 signaling, Wnt signaling, c-Jun N-terminal kinase (JNK), and nuclear factor (NF)-κB signaling pathway, regulating cell growth, differentiation, bacterial autophagy, synaptic transmission, and neuronal morphology. 14 But to our knowledge, whether Rap2B has effects upon tumor angiogenesis has not been reported, and the mechanism has not been illuminated.
In this study, we first carried out a series of experiments in vivo, the result (Figure 1) of which was in line with our previous in vitro study, 19 showing that Rap2B could accelerate RCC growth. Subsequently, we investigated whether tumor angiogenesis could be affected by Rap2B. To our knowledge, serine/threonine kinase AKT is the most important downstream effector of PI3K. 29 PI3K/AKT is one of the important signaling pathways, which is necessary for new blood vessel formation. 38 In addition, PI3K has been implicated that it plays an important role in regulating the expression of pro-angiogenesis factor VEGF. 39 Therefore, we performed Western blot and ELISA assay to further investigate the function of Rap2B on the expression of PI3K/AKT pathway and the levels of secreted VEGF. The results (Figures 3 and 4) showed that the expression of PI3K, phosphorylated forms of AKT, as well as the levels of secreted VEGF were upregulated by Rap2B. On the contrary, either LY294002 or MK2206 could block the levels of phosphorylated AKT and secreted VEGF induced by Rap2B, suggesting that PI3K/AKT signaling pathway could be involved in Rap2B-mediated VEGF upregulation. Interestingly, LY294002 blocked Rap2B-mediated expression of PI3K mRNA but not Rap2B mRNA, indicating that Rap2B indirectly regulated PI3K expression via other signaling pathway, and PI3K/AKT signaling pathway was not required for Rap2B expression (Figure 2).
In the previous studies, the mechanism of a new vasculature formation has been fully recognized.40,41 Angiogenesis induces endothelial cell proliferation and migration, which is a multi-step process of vessel formation. 13 In this study, we provided a novel function of Rap2B in regulating angiogenesis in RCC cells. From the data of HUVEC growth assay and HUVEC tube formation assay, we found that Rap2B can promote HUVECs proliferation and angiogenesis through PI3K/AKT pathway, which is in accordance with the results mentioned above (Figure 5). Subsequently, to further investigate the effects of Rap2B on tumor angiogenesis, we urged to carry out a series of experiments in vivo. As we expected, the result that Rap2B can promote angiogenesis through PI3K/AKT pathway in vivo is in accordance with the results in vitro.
In summary, our data first provided that Rap2B not only can promote tumor growth in vivo but also can promote angiogenesis in vivo and in vitro. Furthermore, we revealed the mechanisms by which Rap2B activated PI3K/AKT pathway, which increased VEGF mRNA level in cell nucleus and finally promoted the VEGF secretion level. This finding probably accounts for the important effect of Rap2B on RCC angiogenesis (Figure 7). Besides, we speculate that there are other potential pathways to support Rap2B-mediated PI3K mRNA level. Based on these results and combined with the fact that angiogenesis is the early step of tumor metastasis which is closely linked with RCC patient death, we can conclude that loss of Rap2B may be a new strategy for RCC anti-angiogenesis therapy. We hope these findings might have shed light on future directions to help block the progression of RCC.

Mechanism model of Rap2B in promoting RCC angiogenesis. Rap2B can activate PI3K/AKT pathway, which increases VEGF mRNA level and the VEGF secretion level and finally promotes RCC angiogenesis.
Footnotes
Acknowledgements
J.D., K.G., and D.Q. contributed equally to this work.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China (No. 81201637) and Natural Science Foundation for colleges and universities in Jiangsu Province (No. 14KJB320023).