
Abstract
Alex3, is a newly identified mitochondrial protein, regulates mitochondrial dynamics and is involved in neural development. However, its expression pattern and clinicopathological relevance in human tumors are still unclear. In this study, Immunohistochemistry assay was performed in 109 cases of lung cancer samples and found that Alex 3 expression in lung cancer tissues was significantly lower than adjacent normal lung tissues (28.4% vs 52.6%, p < 0.001). Sequent statistical analysis indicated that negative Alex3 expression was significantly associated with advanced tumor–node–metastasis stages (p = 0.001), positive lymph node metastasis (p = 0.005), and poor prognosis (p = 0.008). After overexpression of Alex3, levels of p-AKT and Slug were downregulated, while level of E-cadherin was upregulated, which results in the inhibition of invasion and migration ability of lung cancer cells. In conclusion, reduction of Alex3 correlates with the development of non–small cell lung cancer and predicts adverse clinical outcome of non–small cell lung cancer patients. The effect of Alex3 on inhibiting invasion and migration may attribute to upregulation of E-cadherin expression through AKT–Slug pathway inactivation.
Introduction
Alex3 (Arm containing protein Lost in Epithelial cancers linked to the X chromosome 3), encoded by ARMCX3 gene, is localized on chromosome X.1,2 It was demonstrated that Alex3 was composed of a transmembrane domain, a nuclear localization signal (NLS), and six arm-like domains.1,2 López-Doménech and colleagues1,2 had confirmed that Alex3 is localized on the mitochondria and controlled mitochondrial dynamics. Previous studies also indicated that Alex3 was highly expressed in central nervous system.1–4 Kurochkin et al. 5 had established that messenger RNA (mRNA) levels of Alex family proteins Alex1 and Alex2 were absent or dramatically decreased in various human malignant tumors and cell lines. Mirra et al. 3 found that Alex3 may be a suppressor of canonical Wnt signaling in chicken spinal cord. However, the expression and localization of Alex3 in cancerous tissues were still unclear, as well as its biological function.
The AKT signal transduction pathway dominates numerous hallmarks of cancer, including metabolism, cell survival, cell cycle progression, regulation of apoptosis, protein synthesis, motility, and genomic instability by phosphorylation of the substrates.6,7 Slug (SNAI2) is a member of the SNAIL family of zinc finger transcriptional factor that binds with E-box of the promoter, thus suppresses the expression of E-cadherin and facilitates tumor invasion and metastasis.8,9
In this study, we explored the protein and mRNA levels and subcellular distribution of Alex3 in both lung cancer tissues and cell lines, as well as their clinicopathological relevances. We also investigated the effects of Alex3 on the invasiveness of non–small cell lung cancer (NSCLC) cell lines followed by Alex3 overexpression or inhibition. We found that Alex3 depressed invasion of NSCLC cells through suppressing the activation of AKT/Slug/E-cadherin signaling pathway.
Materials and methods
Patients and specimens
This study was approved by the local institutional review board of the China Medical University. Tissue samples were obtained from 109 patients (68 males and 41 females) who underwent complete surgical excision at the First Affiliated Hospital of China Medical University between 2010 and 2012 with a diagnosis of lung squamous cell carcinoma or lung adenocarcinoma. No neoadjuvant radiotherapy or chemotherapy was done before surgery, and all received standard chemotherapy after surgery. All the 109 lung cancer cases contained complete follow-up data. The survival of each patient was defined as the time from the day of surgery to the end of follow-up or the day of death due to recurrence or metastasis. Histological diagnosis and grading were evaluated according to the 2015 World Health Organization (WHO) classification of tumors of lung. 10 All 109 specimens were for histological subtype, differentiated, and in tumor stage. Tumor staging was performed according to the seventh edition of the International Union Against Cancer (UICC) Tumor–Node–Metastasis (TNM) Staging System for Lung Cancer. 11 The median age in 109 patients was 60 years (range: 29–79 years). Of the 109 patients, 49 patients were older than 60 years. The samples included 47 squamous cell lung carcinoma and 62 lung adenocarcinoma. A total of 38 tumors were well differentiated, while 71 were classified as moderately or poorly differentiated. Lymph node metastases were present in 48 of the 109 cases. The tumors included 83 stage I and stage II cases and 26 stage III cases.
Immunohistochemistry
Surgically excised tumor specimens were fixed in 10% neutral formalin, embedded in paraffin, and sliced into 4-µm-thick serial sections. Immunostaining was performed by the streptavidin–peroxidase method. The sections were incubated with anti-Alex3 mouse monoclonal antibody (sc-393752; 1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C for overnight, followed by biotinylated goat anti-mouse IgG secondary antibody. After washing, the sections were incubated with horseradish peroxidase–conjugated streptavidin–biotin (Ultrasensitive; MaiXin, Fuzhou, China) and developed using 3,3′-diaminobenzidine tetrahydrochloride (MaiXin). Finally, samples were lightly counterstained with hematoxylin, dehydrated in alcohol, and mounted. Two investigators blinded to the clinical data semi-quantitatively scored the slides by evaluating the staining intensity and percentage of stained cells in representative areas. The staining intensity was scored as 0 (no signal), 1 (weak), 2 (moderate), or 3 (high). The percentage of cells stained was scored as 1 (1%–25%), 2 (26%–50%), 3 (51%–75%), or 4 (76%–100%). A final score of 0–12 was obtained by multiplying the intensity and percentage scores. Tumors were considered to overexpress Alex3 with score ≥4; those with score <4 were considered to be Alex3 negative or with low expression of the protein.
Western blotting
Total protein was extracted using a lysis buffer (Pierce, Rockford, IL, USA) and quantified with the Bradford method. 12 The protease and phosphatase inhibitors were purchased from Cell Signaling Technology (Danvers, MA, USA). A volume of 50 µg of the total protein samples was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). Membranes were incubated overnight at 4°C with the following primary antibodies: Alex3 (1:100; Santa Cruz Biotechnology); glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:5000; Sigma, St. Louis, MO, USA); Snail, Slug, Myc-tag, p-AKT, AKT, p-P38, P38, p-JNK (phosphorylated c-jun n-terminal kinase), JNK, active-β-catenin, and vimentin (1:1000; Cell Signaling Technology); E-cadherin, N-cadherin, and β-catenin (1:1000; BD Transduction Laboratories, Lexington, KY, USA); and ZO-1 and occludin (1:500; Proteintech, Chicago, IL, USA). AKT-specific inhibitor LY294002 was purchased from Cell Signaling Technology. Membranes were washed and subsequently incubated with peroxidase-conjugated anti-mouse or anti-rabbit IgG (Santa Cruz Biotechnology) at 37°C for 2 h. Bound proteins were visualized using electrochemiluminescence (Pierce) and detected with a bio-imaging system (DNR Bio-Imaging Systems, Jerusalem, Israel).
Cell culture
The HBE cell line was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The A549, H460, H1299, and H661 cell lines were obtained from the Shanghai Cell Bank (Shanghai, China). All cells were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 100 IU/mL penicillin (Sigma), and 100 µg/mL streptomycin (Sigma) and passaged every other day using 0.25% trypsin (Invitrogen).
RNA extraction and real-time polymerase chain reaction
Total cellular RNA was extracted from cells using the RNeasy Plus Mini Kit from (QIAGEN, GmbH, Hilden, Germany). Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using SYBR Green PCR Master Mix (Applied Biosystems) in a total volume of 20 µL on a 7900 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA): 50°C for 2 min, 95°C for 10 min, 40 cycles of 95°C for 15 s, 60°C for 60 s. The sequences of the primer pairs are listed in Table 1. β-actin was used as the reference gene. The relative levels of gene expression were represented as ΔCt = Ctgene − Ctreference, and the fold change of gene expression was calculated by the 2−ΔΔCt method. 13 Experiments were repeated in triplicate. Double fluorescein may report gene analysis
Primers for qPCR.
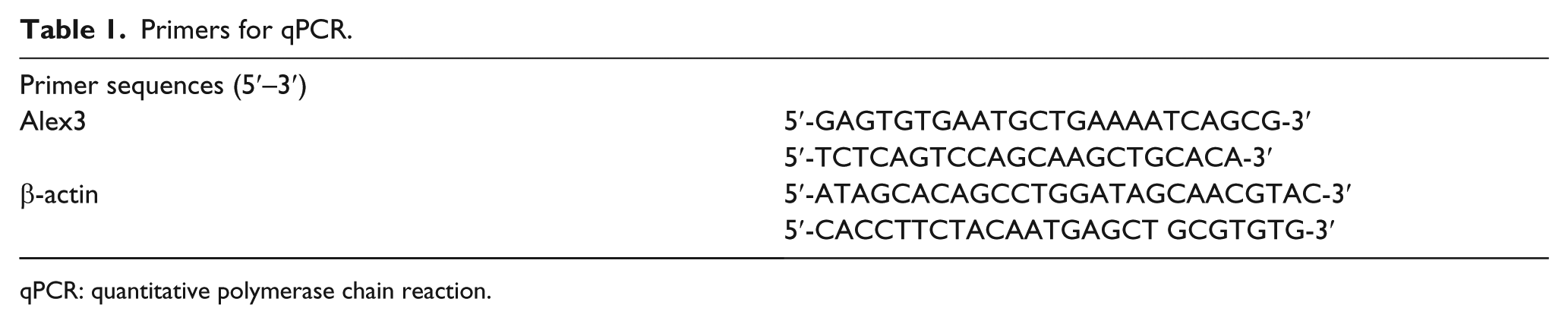
qPCR: quantitative polymerase chain reaction.
Plasmid transfection and small interfering RNA treatment
Plasmids pCMV6-ddk-myc and pCMV6-ddk-myc-Alex3 were purchased from OrigGene (Rockville, MD, USA). Alex3-small interfering RNA (siRNA; sc-91193) and NC-siRNA (sc-37007) were purchased from Santa Cruz Biotechnology. Transfection was carried out using the Lipofectamine 3000 reagent (Invitrogen) according to the manufacturer’s instructions.
Matrigel invasion
Cell invasion assay was performed using a 24-well transwell chamber with 8 µm pores (Costar, Cambridge, MA, USA). The inserts were coated with 20 µL Matrigel (1:3 dilution; BD Biosciences, San Jose, CA, USA). After 48 h of transfection, the cells were trypsinized, and 3 × 105 cells in 100 µL of serum-free medium were transferred to the upper Matrigel chamber for 18 h. Media supplemented with 10% FBS were added to the lower chamber as a chemoattractant. After incubation, the cells that passed through the filter were fixed with 4% paraformaldehyde and stained with hematoxylin. The invasive cells were microscopically counted in 10 randomly selected high-power fields.
Wound-healing assay
In cultures with cell density below 90%, wounds were created in confluent areas using a 200-µL pipette tip after 48 h of transfection. Wound healing within the scrape line was observed at different time points, and representative scrape lines for each cell line were photographed. Duplicate wells were examined for each condition, and each experiment was repeated for three times. The optical wound distances were measured using ImageJ software (National Institute of Health, Bethesda, MD, USA).
Immunofluorescence staining
Cells were fixed with 4% paraformaldehyde, blocked with 1% bovine serum albumin, and incubated overnight with E-cadherin, occludin, and ZO-1 antibodies (1:100; BD Biosciences and Proteintech) at 4°C. Then, the cells were incubated with tetramethylrhodamine isothiocyanate–conjugated secondary antibodies (Cell Signaling Technology) at 37°C for 2 h; cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). Epifluorescence microscopy was performed using an inverted Nikon TE300 microscope (Nikon Co., Ltd, Tokyo, Japan), and confocal microscopy was performed using a Radiance 2000 laser-scanning confocal microscope (Carl Zeiss, Oberkochen, Germany).
Statistical analysis
SPSS version 22.0 for windows (SPSS, Chicago, IL, USA) was used for all analyses. The Pearson’s chi-square test was used to assess possible correlations between Alex3 and clinicopathological factors. Kaplan–Meier survival analyses were carried out in 109 case specimens and compared using the log-rank test. Mann–Whitney U test was used for the image analysis of WB results and the invasive assay results. p < 0.05 was considered to indicate statistically significant differences.
Results
The expression of Alex3 in lung cancer specimens
Initially, we performed immunohistochemical analaysis of 109 cases of lung cancer specimens and 38 cases of non-cancerous tissues. It was revealed that Alex3 was highly expressed in the cytoplasm of normal lung tissues (Figure 1(a)); however, it showed negative or to decrease the cytosolic expression in lung cancer samples (Figure 1(b) and (c)). The positive incidence of Alex3 in lung carcinomas was obviously lower than that in normal lung tissues (31/109, 28.4% vs 20/38, 52.6%, p < 0.001, Figure 1(d)). Statistical analysis results indicated that Alex3 negative expression significantly correlated with advanced TNM stage (p = 0.001) and positive lymph node metastasis (p = 0.005). However, it showed no visible correlation with age, sex, histological type, and differentiation (p > 0.05, Table 2). Kaplan–Meier analysis results showed that the overall survival of patients with positive Alex3 expression (67.222 ± 2.065 months) was significantly longer than those with negative or weak Alex3 expression (53.427 ± 2.930 months, p = 0.008, Figure 1(e)).

The expression of Alex3 in lung cancer specimens. (a) Alex3 revealed strong cytosolic expression in normal bronchial epithelium cells; however, Alex3 showed negative or dim expression in (b) squamous cell carcinoma and (c) adenocarcinoma (magnification: 200×). (d) Alex was highly expressed in non-cancerous tissues than lung carcinoma (magnification: 200×). (e) Kaplan–Meier analysis results indicated that the overall survival of patients with positive Alex3 expression was significantly longer than those without.
Correlation of the expression of Alex3 with clinicopathological features in 109 cases of NSCLC.

Alex3: Arm containing protein Lost in Epithelial cancers linked to the X chromosome 3; NSCLC: non–small cell lung cancer; TNM: tumor–node–metastasis.
The expression of Alex3 in lung cancer cell lines
We next detected Alex3 expression in lung cancer cell lines using western blotting (WB) and quantitative PCR (qPCR). The WB results suggested that Alex3 was weakly expressed in (4/4) lung cancer cells than that in normal bronchial epithelial cells (HBE, Figure 2(a)). qPCR assay also indicated that the mRNA expression level in all the lung cancer tested was lower than that in HBE cells (Figure 2(b)).

The expression of Alex3 in lung cancer cell lines. The (a) protein and (b) mRNA levels of Alex3 was significantly higher in normal bronchial epithelium cell (HBE) than that in lung cancer cells.
Alex 3 decreased lung cancer cell invasion and migration
Then, we overexpressed or depleted Alex3 in H1299 and A549 cells (Figure 3(a) and (b)). Transwell assay results revealed that invasion of H1299 was obviously decreased by transfecting Alex3 plasmids (Figure 3(c)). Correspondingly, the invasiveness of A549 cells was dramatically increased followed by interfering Alex3 (Figure 3(c)). The migration abilities were suppressed or enhanced when we overexpressed or inhibited Alex3 in H1299 or A549 cells (Figure 3(d)).

Overexpression of Alex3 reduced lung cancer cell invasion and migration. (a) Western blot and (b) qPCR analysis of Alex3 protein and mRNA levels after overexpression or silencing of Alex3 in H1299 and A549 cells, respectively. (c) Sequent transwell and (d) wound-healing assay results suggested that overexpression or depletion of Alex3 suppressed or enhanced the invasive and migration abilities of H1299 or A549 cells.
Overexpression of Alex3 downregulated Slug and upregulated E-cadherin
We explored the expression of epithelial–mesenchymal transition (EMT)-related proteins after overexpressing or depleting Alex3 in H1299 or A549 cells. WB results revealed that Slug was downregulated and E-cadherin was upregulated followed by overexpression of Alex3 in H1299 cells (Figure 4(a)). The expression of Slug was upregulated and E-cadherin was downregulated when we depleted Alex3 by siRNA in A549 cells (Figure 4(a)). The other proteins such as Snail, occludin, ZO-1, vimentin, and N-cadherin revealed no obvious alteration (Figure 4(a)). Immunofluorescence (IF) assay results also indicated that E-cadherin expression was increased after the ovexpression of Alex3 in H1299 cells. Accordingly, E-cadherin was decreased followed by interfering of Alex3 by siRNA (Figure 4(b)), whereas the expression and localization of occludin and ZO-1 revealed no remarkable alteration (Figure 4(b)).

Overexpression of Alex3 downregulated Slug and upregulated E-cadherin. (a) When we overexpressed Alex3 in H1299 cells, WB results indicated that the expression of Slug was decreased and E-cadherin was increased. Correspondingly, Slug was upregulated and E-cadherin was downregulated by depleting Alex3 using siRNA. However, the other EMT-related proteins revealed no visible changes. (b) IF results suggested that membranous expression of E-cadherin was increased or decreased followed by upregulating/downregulating Alex3 in H1299 or A549 cells. However, the expression and sublocalization of occludin and ZO-1 revealed no visible changes.
Alex3 suppressed the phosphorylation of AKT
Finally, we screened key signaling involved in regulating Slug and E-cadherin by performing WB. It was shown that the phosphorylation of AKT was decreased or increased after overexpressing or interfering Alex3 in H1299 and A549 cells (Figure 5(a)). However, the other proteins such as p-p38, P38, p-JNK, JNK, active-β-catenin as well as β-catenin displayed no visible changes (Figure 5(a)). To further test whether the upregulation of Slug and downregulation of E-cadherin were caused by suppressing AKT signaling, we added LY294002 (10 µm), a specific inhibitor of AKT signaling, into the medium followed by Alex3 depletion in A549 cells. Sequent WB assay results indicated that the increase of p-AKT and Slug was counteracted, and the expression of E-cadherin was restored (Figure 5(b)).

Overexpression of Alex3 promoted the phosphorylation of AKT. (a) The phosphorylated AKT was decreased or increased followed by overexpressing or interfering Alex3 in H1299 or A549 cells. However, the other key signaling pathway proteins showed no obvious alteration. (b) Upregulation of p-AKT and Slug and downregulation induced by overexpression of Alex3 were reversed by adding AKT-specific inhibitor LY294002.
Discussion
We found that Alex3 was underexpressed in lung cancer specimens compared with corresponding non-cancerous tissues. The negative expression of Alex3 was significantly correlated with advanced TNM stage, positive lymph node metastasis, and poor prognosis. Although there is a NLS sequence localized in the N-terminal region,1,2 we had detected no nuclear expression in lung cancer specimens. Kurochkin et al. 5 had demonstrated that the mRNAs of Alex1 and Alex2 were downregulated in numerous human carcinomas, which was consistent with our findings. Our finding as well as Kurochkin’s studies indicated that Alex family proteins may be reduced in cancerous tissues, which may be tumor suppressor. The reason why Alex family proteins were decreased in cancerous tissues should be further tested in future.
We also suggested that overexpression of Alex3 inhibited lung cancer cell invasion and migration. As we know, EMT was mainly involved in NSCLC invasion and metastasis; therefore, we detected EMT-related proteins after upregulating/downregulating Alex3. Sequent WB assay revealed that Slug was downregulated and E-cadherin was upregulated. Previous studies had demonstrated that Slug may reduce E-cadherin, thus enhanced tumor invasion.8,14,15 We then explored key signaling pathways involved in the downregulation Slug. The results suggested that the phosphorylation of AKT was dramatically interfered after overexpression of Alex3. Adding LY2094002 after inhibiting Alex3 counteracted the increase of p-AKT and Slug and restored the expression of E-cadherin, which indicated that AKT was served as an upstream factor of Slug and E-cadherin. Previous studies had demonstrated that activation of AKT signaling increased Slug expression,16,17 which was consistent with our finding. However, the other key signaling proteins such as active-β-catenin and p-JNK revealed no visible changes. Serrat had demonstrated that activation of non-canonical Wnt pathway promoted the degradation of Alex3, 2 which indicated that Alex3 may serve as downstream factor of non-canonical Wnt signaling. However, Mirra et al. 3 had confirmed that Alex 3 inhibited canonical Wnt signaling in chicken spinal cord, which was diverse from what we had found. It may be caused by species and histological differences. The effect of Alex3 on canonical Wnt signaling should also be further studied.
In conclusion, Alex3 was expressed in the cytoplasm of non-cancerous tissues and reduced in lung cancer specimens. Negative Alex3 expression was significantly correlated with high TNM stage, positive lymph node metastasis, and poor prognosis. Alex3 inhibited Slug and restored E-cadherin expression via inactivating AKT signaling, thus depressed invasion and migration of NSCLC cells.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (No. 81572854 to E.W. and No. 81602012 to X.Z.).