
Abstract
To investigate the effects of soluble fms-like tyrosine kinase-1 on the vascular mimicry formation, proliferation, migration, and invasion of colorectal cancer SW480 cells. The recombinant plasmid pBLAST49-sFLT-1 or pBLAST49 control plasmid was transfected into SW480 cells to obtain hsFLT-1-SW480 or Ctrl-SW480 cells. The three-dimensional model culture, sulforhodamine B assay, scratch assay, and Transwell assay were performed to detect the vascular mimicry formation, proliferation, migration, and invasion of colorectal cancer SW480 cells, respectively. Western blotting was used to detect the expression of vascular endothelial–cadherin protein. Compared with Ctrl-SW480 cells, vascular mimicry formation ((0.85 ± 0.04) vs (7.40 ± 0.69), p < 0.05) and vascular endothelial–cadherin expression ((1.25 ± 0.08) vs (1.89 ± 0.03), p < 0.05) were significantly decreased, and the growth rate was also significantly decreased in hsFLT-1-SW480 cells ((32.54 ± 5.12) vs (88.13 ± 11.52), p < 0.05). Moreover, the migration ((0.46 ± 0.08) vs (0.94 ± 0.03), p < 0.05) and invasion capacity ((59.14 ± 3.64) vs (134.85 ± 10.16), p < 0.05) of SW480 cells were significantly inhibited upon soluble fms-like tyrosine kinase-1 transfection. soluble fms-like tyrosine kinase-1 inhibits cell proliferation, migration, and invasion of colorectal cancer SW480 cells through suppression of vascular mimicry formation, which provides a good basis for the development of new drugs for the treatment of colorectal cancer by targeting both angiogenesis and vascular mimicry formation.
Introduction
Tumor angiogenesis plays an important role in tumor growth, proliferation, migration, and invasion; therefore, anti-angiogenic therapy has become a major strategy for cancer treatment. So far, despite the rapid development of anti-angiogenic drugs, the clinical efficacy remains unsatisfied. In tumor tissues, tumor blood vessels mainly exist in two forms: one is the vascular structure surrounded by vascular endothelial cells and the other is the lumen structure surrounded by the tumor cells, also known as vascular mimicry (VM). 1 It has been reported that the anti-angiogenic drug bevacizumab (BV) has no inhibitory effect on the VM, leading to tumor metastasis promotion. Therefore, development of drugs targeted for blood vessels and/or VM is imminent. 2 Soluble fms-like tyrosine kinase-1 (sFLT-1) is a transmembrane glycoprotein with tyrosine kinase activity and is a potent inhibitor of vascular endothelial growth factor (VEGF). sFLT-1, which is mainly produced by trophoblast cells and vascular endothelial cells, is supposed to be a major angiogenesis inhibitor with strong anti-angiogenic effect.3,4 In this study, we applied the colon cancer SW480 cell line which has high VM-forming ability to investigate the effect of sFLT-1 on VM formation.
Materials and methods
Cells and reagents
Human colon cancer cell line SW480 was purchased from Cell Bank of the Chinese Academy of Science, Shanghai, China. SW480 and human umbilical vein endothelial cells (HUVEC) were cultured in RPMI 1640 medium containing penicillin, streptomycin, and 10% fetal bovine serum (FBS) at 37°C in an incubator with 5% CO2. RPMI 1640 medium, FBS, and trypsin were purchased from Gibco (Carlsbad, CA, USA). Reverse transcription kit was purchased from TaKaRa (Dalian, China). Polymerase chain reaction (PCR) kit was purchased from Invitrogen (Carlsbad, CA, USA). Antibodies against FLT-1, VE-cadherin, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were from Abcam (Cambridge, MA, USA), and pBLAST49-hsFLT-1 recombinant plasmid and pBLAST49 were purchased from Invitrogen (Carlsbad, CA, USA). Matrigel gum was purchased from BD Biosciences (San Diego, CA, USA).
Transient transfection
SW480 cells were seeded in six-well plates to a confluence of 80%–90% with antibiotic-free medium 24 h prior to transfection. A volume of 4 µg pBLAST49-hsFLT-1 or pBLAST49 control plasmid was diluted in 250 µL of serum-free medium and 5 µL of liposomes was diluted in 250 µL serum-free medium. After 5 min incubation at room temperature, the diluted plasmid was combined with the diluted liposome and mixed gently. After incubation at room temperature for 20 min, the final mixture was added onto the cells and incubated at 37°C in an incubator with 5% CO2. Culture medium was changed to be completed medium 6 h after incubation. The transfected cells were named as hsFLT-1-SW480 cells and Ctrl-SW480 cells, respectively, for the subsequent experiments.
Reverse transcription polymerase chain reaction to detect the expression of sFLT-1 messenger RNA
Total RNA was extracted from SW480 cells and then reverse-transcribed to complementary DNA (cDNA) using the reverse transcription kit according to manufacturer’s instructions (TaKaRa). The expression of FLT-1 was determined using reagent from Invitrogen. GAPDH was used as an internal control. The reaction system was as follows: 95°C for 5 min followed by 36 cycles of 95°C for 30 s, 60°C for 60 s, and 72°C for 60 s, with an extension at 72°C for 10 min. The primers were as follows: FLT-1 (forward: 5′-ATTTCTGTGAAGGGCGAGTT-3′, reverse: 5′-GCCGAGGTGAAGAGGTTT-3′) and GAPDH (forward: 5′-TGATGACATCAAGAAGGTGG-3′, reverse: 5′-TTTCTTACTCCTTGGAGGCC-3′). All primers were synthesized by Invitrogen Corporation. The photographs of gels were analyzed by Quantity One image analysis software. Each experiment was repeated three times.
Three-dimensional cell culture to detect the effect of sFLT-1 on VM-forming ability of SW480 cells
The experimental groups were divided into HUVEC cell group (positive control group), untransfected group, Ctrl-SW480 group, hsFLT-1-SW480 group, and FLT-1 neutralizing antibody treatment group (FLT-1Ab-SW480); 24-well plates were coated with 100 µL Matrigel per well. The HUVEC cells, SW480 cells, Ctrl-SW480 cells, hsFLT-1-SW480 cells, and FLT-1Ab-SW480 cells were trypsinized, re-suspended, and adjusted into a density of 1.5 × 105 mL−1. Then, 1 mL cell suspension was added into 24-well plates with good Matrigel plastic bag (for FLT-1Ab-SW480 group, 1 ng/mL FLT-1 neutralizing antibody was added) and incubated at 37°C in an incubator with 5% CO2 for 48 or 72 h. Five horizons were randomly selected under a powerful microscope, and the number of luminal cells formed was counted. Each experiment was repeated three times.
Western blotting to detect the expression of sFLT-1 and VE-cadherin proteins
Total protein was extracted from cells and the concentration was measured using the bicinchoninic acid assay (BCA) protein assay. Proteins were separated by 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel and transferred to polyvinylidene difluoride (PVDF) membranes. Membranes were incubated with mouse anti-human FLT-1 antibody (1: 2000 dilution), VE-cadherin antibody (1: 2000 dilution), or GAPDH antibody (1: 5000 dilution) at 4°C overnight followed by incubation with horseradish peroxidase (HRP)-labeled goat anti-mouse secondary antibody (1: 5000 dilution) at room temperature for 1 h. Protein bands were visualized using ECL kit and image gradation was quantified using the Quantity One analysis software. Each experiment was repeated three times.
SRB assay to detect the effect of sFLT-1 on the proliferation of SW480 cells
A total of 5 × 103 cells were seeded in 96-well plates. After cultured for 48 h, cells were fixed with 50 µL TCA 50 Yu per well at 4°C for 1 h, and then, cells were stained with 100 µL SRB per well at room temperature for 30 min. After washing with 1% acetic acid, 150 µL Tris solution per well was added onto cells. After shaking for 5 min, the enzyme-linked immunosorbent assay (ELISA) optical density (D) was measured at a wavelength of 515 nm. Each experiment was repeated three times. The cell proliferation rate is calculated as follows: the cell proliferation rate (%) = (D value of untreated cells − cells treated with D value)/D value of the cells were treated group × 100%.
Scratch assay to detect the effect of sFLT-1 on SW480 cell migration
A total of 8 × 106 cells were seeded in six-well plates. After the cell confluence reaches 80%, cell layers were carefully scratched with 200 µL sterile pipette tips and washed three times with phosphate-buffered saline (PBS). Cells were photographed at the indicated time points (0, 12, 24, 48, and 72 h). Image analysis was processed using ImageJ software. Each experiment was repeated three times.
Transwell invasion assay to detect the effect of sFLT-1 on SW480 cell invasion
Transwell invasion assay was performed using Transwell chambers with filter inserts of Matrigel basement membrane. Cells were trypsinized and adjusted to be a density of 50 × 105 mL−1. A volume of 200 µL of cell suspension was placed in the upper chamber, whereas 500 µL conditioned medium was placed in the lower chamber. After cultured for 48 h, cells that migrated to the bottom of the membrane were stained with crystal violet. Five randomly selected horizons were counted under the microscope. Cell invasiveness was represented as cell number per field. Each experiment was repeated three times.
Statistical analysis
Statistical analysis was performed using SPSS 16.0 software package (SPSS Company, Chicago, IL, USA). The results are expressed as the mean ± standard deviation (SD) of at least three independent experiments. Multiple comparisons were performed using one-way analysis of variance (ANOVA). A probability level of 5% was considered significant (p < 0.05) unless otherwise stated.
Results
sFLT-1 transfection enhanced the expression of sFLT-1 mRNA in SW480 cells
Primers among sFLT-1 extracellular domain were used to detect the expression of sFLT-1 mRNA by reverse transcription polymerase chain reaction (RT-PCR), which indirectly reflected the expression of sFLT-1 mRNA in hsFLT-1-SW480 cells. The results showed that the extracellular domain of FLT-1 was expressed in HUVEC cells, SW480 cells, Ctrl-SW480 cells, and hsFLT-1-SW480 cells (Figure 1). However, expression of sFLT-1 mRNA was significantly higher in hsFLT-1-SW480 cells than that in Ctri-SW480 cells ((0.36 ± 0.02) vs (0.19 ± 0.03), p < 0.01), whereas there was no statically significant difference between SW480 cells and Ctrl-SW480 cells ((0.19 ± 0.04) vs (0.16 ± 0.02), p > 0.05).

Expression of sFLT-1 mRNA in SW480 cells after hsFLT-1 transfection.
Expression of sFLT-1 protein in SW480 cells after transfection
Western blotting results showed that FLT-1 protein was expressed in HUVEC cells, SW480 cells, and Ctri-SW480 cells (Figure 2). In hsFLT-1-SW480 cells, not only the FLT-1 protein, but also the small-fragment sFLT-1 protein that exhibited a relative molecular mass of 58,000 Da was detected, whereas expression of sFLT-1 protein was not observed in other cell lines. These results suggested that exogenous sFLT-1 protein was successfully expressed in colon cancer SW480 cells.

Expression of sFLT-1 protein in SW480 cells after hsFLT-1 transfection.
sFLT-1 transfection inhibited VM formation
In three-dimensional (3D) culture model, HUVEC cells formed a number of typical VM lumen structures after cultured for 24 h, and similar VM lumen structures were found in SW480 and Ctrl-SW480 cells. However, VM lumen structures in hsFLT-1-SW480 cells or FLT-1 Ab-SW480 cells were significantly decreased compared with those in Ctrl-SW480 cells or SW480 cells (Figure 3). The number of lumen was significantly reduced in hsFLT-1-SW480 cells and FLT-1 Ab-SW480 cells compared with Ctrl-SW480 cells ((0.85 ± 0.04) and (1.26 ± 0.37) vs (7.40 ± 0.69), p < 0.01). However, no significant difference was found in the number of VM lumens between Ctrl-SW480 cells and SW480 cells ((7.40 ± 0.69) vs (7.83 ± 0.92), p > 0.05; Figure 4).

Effect of hsFLT-1 transfection on VM formation in SW480 cells.

Effect of hsFLT-1 transfection on the number of VM lumens in SW480 cells (*p < 0.05 vs Ctrl-SW480 cells).
sFLT-1 transfection inhibited the expression of VE-cadherin protein
Western blotting analysis showed that the protein levels of VE-cadherin in hsFLT-1-SW480 and FLT-1 Ab-SW480 cells were significantly lower than that in Ctrl-SW480 cells ((1.25 ± 0.08) and (1.31 ± 0.01) vs (1.89 ± 0.03), p < 0.05; Figure 5), whereas there was no significant difference in VE-cadherin expression between SW480 cells and Ctrl-SW480 cells. These results suggested that sFLT-1 or FLT-1 neutralizing antibody significantly inhibited the expression of VE-cadherin protein.
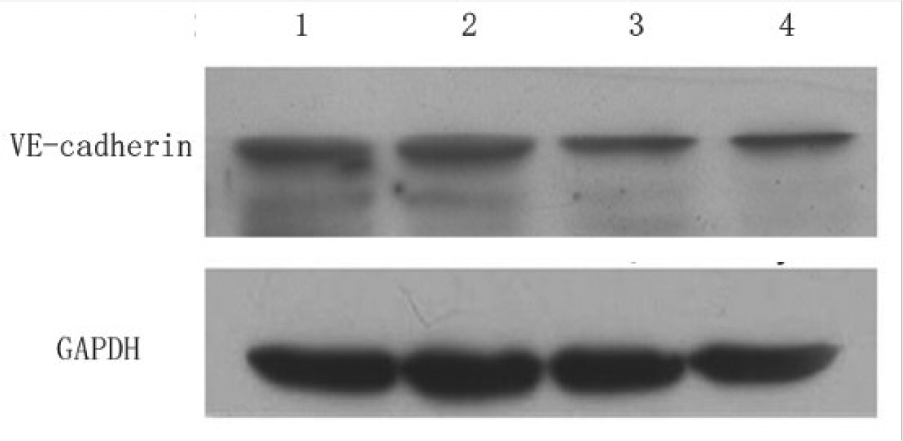
VE-cadherin expression in SW480 cells transfected with hsFLT-1.
sFLT-1 transfection inhibited the proliferation of SW480 cells
SRB assay showed that compared with Ctrl-SW480 cells, the viability of FLT-1 Ab-SW480 cells was significantly decreased ((54.28% ± 9.97%) vs (88.24% ± 21.94%), p < 0.05). Moreover, the growth rate of hsFLT-1-SW480 cells was even lower than that of FLT-1 Ab-SW480 cells ((32.54% ± 5.12%) vs (54.28% ± 9.97%), p < 0.01), while there was no significant difference between Ctrl-SW480 and SW480 cells (p = 0.116). These results suggested that sFLT-1 or FLT-1 neutralizing antibody significantly inhibited the proliferation of human colon cancer SW480 cells.
sFLT-1 transfection inhibited SW480 cell migration
Scratch assay showed that scratches in SW480 and Ctrl-SW480 cells were almost completely healed after 72 h, while scratches in FLT-1 Ab-SW480 cells and hsFLT-1-SW480 cells were not (Figures 6 and 7). Results from ImageJ software analysis showed that compared with Ctrl-SW480 cells, the healing capacity was significantly decreased in FLT-1 Ab-SW480 cells and hsFLT-1-SW480 cells ((0.38 ± 0.07) and (0.46 ± 0.08) vs (0.94 ± 0.03) mm, p < 0.01). But no significant difference was observed between Ctrl-SW480 and SW480 cells (p = 0.520). These results suggested that FLT-1 or FLT-1 neutralizing antibody significantly inhibited the migratory ability of SW480 cells.

Effect of hsFLT-1 transfection on SW480 cell proliferation (*p < 0.05 vs Ctrl-SW480 cells).

Effect of hsFLT-1 transfection on SW480 cell migration (100×).
sFLT-1 transfection inhibited SW480 cell invasion
Transwell invasion assay showed that the invasion ability of FLT-1 Ab-SW480 cells and hsFLT-1-SW480 cells was significantly lower than that of Ctrl-SW480 cells ((51.27 ± 4.08) and (59.14 ± 3.64) vs (134.85 ± 10.16), p < 0.01; Figure 8). However, no significant differences were found between Ctrl-SW480 cells and SW480 cells (p = 0.148), indicating that sFLT-1 or FLT-1 neutralizing antibody significantly inhibited the invasion of SW480 cells.

Effect of hsFLT-1 transfection on SW480 cell invasion (200×).
Discussion
VM refers to the network of blood vessel lumen that was surrounded by aggressive tumor cells. VM was first observed in aggressive human intraocular and metastatic cutaneous melanomas by Maniotis et al. 5 The 13-year extensive and in-depth studies have demonstrated that this VM is different from the classic tumor angiogenesis VM widespread in colorectal cancer, 6 melanoma, 7 liver cancer, 8 osteosarcoma, 9 and other tumor tissues. VM is a new model of microcirculation, which plays an important role in tumor invasion and is a key factor of poor prognosis. VM is an appealing target for novel strategies of cancer treatment. 10 The main anti-angiogenesis drugs discovered in current years not only influence the VM formation but also promote its growth. For example, Soda et al. 11 reported that in a mouse ovarian cancer model, short-term use of BV significantly promotes the formation of VM, and VM cannot be suppressed by cisplatin. Combination of the two drugs promotes VM formation and accelerates tumor metastasis. In vitro and in vivo studies showed that treatment with anti-VEGF cannot suppress VM formation in glioblastoma cells.12,13 van der Schaft et al. 12 have reported that the classical angiogenesis inhibitor endostatin fails to inhibit the formation of the VM. Other reports have also showed that treatment with BV or sunitinib generates hypoxia and increased the population of breast cancer stem cells (CSC), which are important factors prompting VM formation.13,14 Therefore, development of new drugs that inhibit both angiogenesis and VM formation of cancers is of imperative.
Angiogenesis is regulated by a variety of complex signaling pathways. Aside from the Akt/mTOR pathway, AT1R/FLT-1 axis is a newly discovered signaling pathway closely related to blood vessels.13,15 FLT-1 is a tyrosine kinase receptor widely expressed in tumor tissues and peripheral vascular endothelial cells, which plays an important role in tumor angiogenesis. FLT-1 is involved in tumor angiogenesis and development, maturation, plasticity, as well as reconstruction. 16 Low expression of FLT-1 is found in normal vascular invasion in colorectal cancer lack of VM, while significantly higher expression was observed in colorectal cancer and its surrounding angiogenesis. 17 FLT-1 induces tumor blood vessels and promotes blood vessel sprouting extension, promoting the tumor vascular angiogenesis, and blockage of FLT-1 inhibits tumor growth and angiogenesis. 18 Part of blocking VEGF pathway has no biological effect on tumors that exhibit abnormal expression of FLT-1, indicating that FLT-1 is an important modulator of tumor blood vessels. Ang/FLT-1 signaling pathway can be raised by MSK1 survivin and Refl can enhance the resistance of human myeloma cells IM9 to BV. 19 Given the important role of AT1R/FLT-1 in the regulation of tumor occurrence and development, FLT-1 is supposed to be a new target for cancer therapy. 16 It has been showed that the synthesized sFLT-1 competitively binds Ang and inhibits AT1R/FLT-1 signaling pathway, thus effectively inhibiting the growth and angiogenesis in mouse colon cancer cells. 20 In this study, we found that transfection of hsFLT-1 significantly decreased VM formation and VE-cadherin expression in SW480 cells. Similar results were also observed by using a post-FLT-1 neutralizing antibody. hsFLT-1 and FLT-1 neutralizing antibodies significantly inhibited the proliferation, migration, and invasion of SW480 cells. These results suggest that sFLT-1 suppresses SW480 cell proliferation, migration, and invasion via inhibition of VM formation, indicating that sFLT-1 may be used for colon cancer treatment; however, the detailed molecular mechanism still needs further investigations.
Current researches on the mechanism of VM formation 21 suggest that hypoxic conditions, VEGF, matrix metalloproteinase (MMP), microRNA (miRNA), CSC, and epithelial–mesenchymal transition (EMT) are the important mediators regulating VM formation. Among these, Ang/FLT-1 signaling pathway and the MMPs, in particular MMP2 or MMP9, are most closely related. 22 It is possible that FLT-1 inhibits VM formation by inhibiting MMP2 or MMP9 expression, but this hypothesis still needs to be experimentally demonstrated in the follow-up studies.
In summary, sFLT-1 inhibits tumor VM formation and thereby inhibits tumor cell proliferation, migration, and invasion. These results will provide ideas for developing anti-VM and anti-angiogenic double-targeted drugs.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.