
Abstract
The tumor microenvironment can act so as to stimulate or reject tumor cells. Among the determining factors are cytokines produced, for example, by infiltrating immune cells, tumor cells, and fibroblasts. External radiotherapy has been shown to be able to activate an immune response against tumor cells with cytokine signaling as an important part of the activation. The aim of this study was to evaluate the cytokines present in the tumor microenvironment and whether the cytokine profile changed during tumor regression induced by radioimmunotherapy with the beta emitter 177Lu. Immunocompetent rats with colon carcinoma tumors were injected with 400 MBq/kg 177Lu-mAb, and the tumors were excised after 1, 2, 3, 4, 6, or 8 days post injection (4 rats/day on days 1–6 and 8 rats on day 8). Tumors from 10 untreated rats were used as control tissue. The tumors were divided into half: one half was prepared for cytokine analysis with a cytokine array kit and the other half was used for histological analysis. A total of 18 of the 29 cytokines evaluated were detected in this tumor model, and the majority of these act in a pro-inflammatory manner or stimulate the infiltration of immune cells. The differences between treated tumors and control tumors were small, thus the cytokine profile in the untreated tumors did not transfer to an anti-inflammatory profile during tumor regression induced by radioimmunotherapy with 177Lu. Histological evaluation demonstrated a heterogeneous pattern of ongoing cell death and the formation of granulation tissue.
Keywords
Introduction
Tumors are complex structures consisting of cellular and non-cellular components such as tumor cells (different clones), the extracellular matrix, and stromal cells.1–3 The tumor microenvironment promotes tumor growth by the secretion of paracrine stimulatory factors, angiogenesis, and immune-mediated interactions. 2 Cytokines are small signaling proteins that can be produced by a broad range of cells. They can act in either a tumor-promoting or a tumor-rejecting manner and sometimes have dual roles depending on concentration and the combination of cytokines present. 4 Radiation has been shown to affect cytokine expression,4–6 thus radiotherapy could change the tumor microenvironment. This change could be part of the immune activation thought to explain the abscopal effect, in which local irradiation results in the regression of a metastasis at a distance from the irradiated site. 7
Targeted therapy is a cornerstone in personalized medicine with the aim to enhance the specificity of the treatment in order to spare normal tissue. In radioimmunotherapy (RIT), a tumor-targeting antibody is labeled with a radionuclide in order to achieve local irradiation of tumor cells. The effects depend on several factors including target antigen expression and the properties of the radionuclide. In this study, a monoclonal antibody (mAb) was labeled with the beta-emitting radionuclide lutetium-177 (177Lu) and evaluated in a syngeneic rat colon carcinoma model. This treatment has previously resulted in complete response (CR, defined as non-palpable tumor for at least 1 week) in 17 out of 19 rats. 8 Analysis of tumors during regression has shown that tumors transformed from dense growth of tumor cells to the formation of granulation tissue with ongoing cell death located mainly in the peripheral parts of the tumor. 9 The tumors were also infiltrated by neutrophils and lymphocytes. 9 Analysis of immune cell markers demonstrated that T cells and macrophages augmenting tumor rejection were present to a higher extent than those associated with immune tolerance, and that the number of immune cells decreased as a result of the treatment. 10 After treatment, approximately half of the animals in the syngeneic rat model developed metastases within the follow-up period of 100 days. The number of animals with metastases increased after the depletion of CD8-positive cells, 11 indicating that the immune system may be important in preventing the development of metastases. This motivated us to examine the tumor microenvironment prior to and during RIT in the same immunocompetent rat tumor model. The aim of this study was to evaluate the cytokine profile of untreated tumors and of tumors treated with RIT in our immunocompetent rat model to further characterize the effects of 177Lu-mAb on the immune system.
Methods
The syngeneic rat model
BN7005-H1D2 is a cell line established from a 1,2-dimethylhydrazine-induced colon carcinoma in a Brown Norway (BN) rat. 12 The radiosensitivity of the cell line is similar to that of human colorectal carcinoma cell lines. 13 We have determined the survival fraction of BN7005-H1D2 cells after 2 Gy irradiation with a 137Cs source to be 0.5 (unpublished data). The cells were cultured in RPMI-1640 medium supplemented with 10% fetal calf serum, 1 mM sodium pyruvate, 10 mM HEPES buffer, and 14 mg/L gentamicin (all from Gibco, Invitrogen, Carlsbad, CA) at 37°C, in a humidified environment containing 5% CO2. The cells were washed in phosphate-buffered saline (PBS) and detached by treatment with trypsin/ethylenediaminetetraacetic acid (EDTA) (PAA Laboratories GmbH, Pasching, Austria).
BN rats are immunocompetent and express the target antigen (Lewis Y) in normal tissues, mainly in the epithelium of the gastrointestinal tract, 14 which is similar to humans. 15 The animals were inoculated with 3 × 105 tumor cells between the abdominal wall and the peritoneum under general anesthesia (isoflurane; Abbott Scandinavia AB, Solna, Sweden). Tumor volumes were calculated as tumor length × tumor width2 × 0.4.
The animals were housed under standard conditions and fed with standard pellets and fresh water ad libitum. All experiments were conducted in compliance with European legislation on animal welfare and were approved by the Regional Animal Ethics Committee.
The radioimmunoconjugate
The mouse/human chimeric monoclonal IgG1 antibody BR96 (Seattle Genetics Inc., Seattle, WA) was used. This binds to the tumor-associated antigen Lewis Y that is expressed on the majority of human epithelial tumors. The dissociation constant between BR96 and the cell line used is 4 nM, 16 illustrating the strong binding affinity of the mAb to the tumor cells.
Conjugation of BR96 and S-2-(4-isothiocyanatobenzyl)-1,4,7,10-tetraazacyclododecane tetraacetic acid (DOTA) was performed according to Forrer et al. 17 Briefly, BR96 was transferred to 0.2 M sodium carbonate buffer, pH 9.5, by repeated centrifugation using an Amicon Ultra-15 filter (molecular weight cut-off 30 kDa; Millipore, Billerica, MA). All empty vials were pretreated with 1% HNO3, and all buffers were pretreated with Chelex-100 (BioRad, Hercules, CA) to remove metals. The DOTA chelate (2 mg/mL H2O; Macrocyclics, Dallas, TX) was added to the BR96 antibody (100 mg/mL) at a molar ratio of 3:1 (DOTA:BR96) and incubated for 1 h at 37°C. The conjugate was purified by repeated centrifugation using an Amicon Ultra-15 filter and transferred to 0.25 M ammonium acetate buffer, pH 5.3, and the final concentration was adjusted to 10 mg/mL BR96. Matrix-assisted laser desorption/ionization–mass spectrometry (MALDI-MS) was used to determine the number of DOTA moieties per BR96 molecule, by desalting the sample to 18 MΩ · cm H2O using a centrifugation filter device, and dividing the increase in molar mass by 688 (the molecular mass of the DOTA chelate).
The immunoreactivity (i.e. the antigen-binding properties) of DOTA-BR96 in relation to BR96 was determined from a saturation binding curve, using BN7005-H1D2 cells as the target antigen. Briefly, increasing concentrations of BR96 and DOTA-BR96 (20 ng/mL–40 µg/mL) were added to the cell plate in triplicate and incubated for 90 min. The bound BR96/DOTA-BR96 conjugate was detected with rabbit anti-human IgG-horseradish peroxidase (HRP) (Dako, Glostrup, Denmark), and the equilibrium binding constant (Kd) was calculated using Prism 5.02 software (GraphPad Software Inc., binding saturation; one site total, non-specific binding and background constrained to a constant value of zero). The immunoreactivity was given by the ratio of the binding constants: Kd(BR96)/Kd(DOTA-BR96).
Both the DOTA-BR96 conjugate and the 177LuCL3 solution (MDS Nordion, Vancouver, Canada) were incubated at 45°C for 10 min. The DOTA-BR96 solution was added to the vial containing the radionuclide and incubated at 45°C for 15 min. The reaction was quenched with an excess of DTPA (diethylenetriaminepentaacetic acid) for 5 min. Human serum albumin (HSA; Baxter Medical AB, Kista, Sweden) was added to a final concentration of 1% to prevent radiolysis from affecting the immunoreactivity. High-performance liquid chromatography (HPLC) (7.8 × 300 mm molecular sieving column; Phenomenex SEC S3000, eluted in 50 mM sodium phosphate at 1.0 mL/min) was used to determine the radiochemical purity and to detect signs of aggregation or fragmentation.
RIT with 177Lu-DOTA-BR96
A total of 38 tumor-bearing male BN rats (Harlan, Horst, Netherlands) were included in the study. Their median weight on the day of administration of RIT was 276 g. In total, 28 rats were treated with 400 MBq/kg body weight of 177Lu-DOTA-BR96 (150 µg DOTA-BR96 in 0.4 mL saline with 1% HSA) by intravenous injection in the tail vein 13–14 days after cell inoculation. Our previous study showed that 400 MBq/kg body weight resulted in CR in 17 of 19 animals, 8 and that the maximum tolerable activity was 600 MBq/kg body weight. 18 The treated animals were sacrificed and dissected 1, 2, 3, 4, 6, and 8 days p.i. in groups of 4 animals per day (8 animals on day 8), while 10 untreated animals used as controls were sacrificed and dissected on the day of treatment. The tumors were cut into half, one half was cut into small pieces and placed in vials with protease inhibitor solution (containing 10 µg/mL aprotinin, 10 µg/mL leupeptin, and 10 µg/mL pepstatin from Sigma-Aldrich, St. Louis, MO) and homogenized (DISP 25, from InterMed, Roskilde, Denmark). Triton X-100 (Sigma-Aldrich) was added to the homogenates (final concentration of 1%) before storage at −80°C. The other half was fixed in 4% paraformaldehyde and embedded in paraffin for histological evaluation. In cases where the remaining tumor was small or only scar tissue remained, the whole tumor was used for cytokine analysis.
Cytokine evaluation
The tumor samples with protease inhibitors were centrifuged before determining the protein concentration with a bicinchoninic acid assay. In short, tumor samples were diluted to an expected concentration of 200–1000 µg/mL and added in triplicate to Nunc-Immuno™ Polysorp 96-well plates. Increasing concentrations of protein standard (100–1000 µg/mL, Bovine IgG1; BioRad) were also added in triplicate. A total of 50 parts of bicinchoninic acid solution were mixed with one part copper(II)sulfate pentahydrate solution (both from Sigma-Aldrich) and added to the plates (200 µL/well), which were then incubated for at least 2 h. The plates were read in an enzyme-linked immunosorbent assay (ELISA) spectrophotometer at 562 nm (VERSAmax microplate reader, Molecular Devices, Sunnyvale, CA). The concentrations of the tumor samples were calculated using a linear standard curve obtained from the protein standard.
The Proteome Profiler™ Rat Cytokine Array Kit, Panel A (R&D Systems, Minneapolis, MN) was used according to the manufacturer’s recommendations. Briefly, the cytokine array membrane was blocked with Array Buffer 6 for 1 h on a rocking platform shaker. The tumor sample (400 µg) was added to 0.5-mL Array Buffer 4, and the final volume was adjusted to 1.5 mL with Array Buffer 6. Detection Antibody Cocktail (15 µL) was added to the prepared samples, which were then incubated for 1 h at room temperature. The sample–antibody mixture was added to the membrane, which was incubated at 2°C–8°C on a rocking platform shaker overnight. The membrane was washed three times with wash buffer, for 10 min each. The streptavidin–HRP was diluted in Array Buffer 6 and added to the membrane, which was incubated for 30 min at room temperature on a rocking platform shaker. The membrane was washed three times with Wash Buffer as before. The Chemi Reagent mix was prepared and added to the membrane for 1 min. The Chemi Reagent mix was then removed from the membrane by drying, and the membrane was placed in a plastic sheet protector. The cytokines present were detected and analyzed using a chemiluminescence detector system (FluorChem FC2; Alpha Innotech, San Leandro, CA). The mean luminescence was normalized to reference spots from the same membrane after background correction.
Analysis and evaluation of biomarkers
The paraffin-embedded tumor samples were sectioned (4 µm) and consecutive sections were used for the evaluation of biomarkers. The antibody administered and fragmented DNA were stained as described below. In addition, overall histology was visualized by hematoxylin and eosin staining.
For immunological staining, the sections were incubated at 60°C prior to storage at 4°C. Sections were rehydrated and heat-mediated antigen retrieval was performed using a 2100 Antigen Retriever pressure cooker (Aptum, Southampton, UK) with acidic Target Retrieval Solution (pH 6). The sections were blocked with Peroxidase-Blocking Solution (Dako), the Biotin-Blocking System (Dako), and VECTASTAIN® goat normal serum (from VECTASTAIN Elite ABC kit (human IgG); Vector Laboratories, Peterborough, UK). The sections were incubated with VECTASTAIN anti-human IgG biotinylated antibody for 30 min at room temperature. After washing, the sections were incubated with VECTASTAIN ABC reagent (prepared 30 min in advance) for 30 min at room temperature. Finally 3,3-diaminobenzidine (DAB; Dako) was added after washing, followed by counterstaining with hematoxylin, dehydration, and mounting.
The fragmented DNA was visualized with the TUNEL assay (TACS® 2 TdT-Blue Label In Situ Apoptosis Detection Kit, Trevigen, Gaithersburg, MD). The TUNEL assay was performed according to the manufacturer’s recommendations. Briefly, after storage at 4°C, the sections were incubated at 60°C for 5 min, rehydrated, and treated with Proteinase K Solution for 15 min at 37°C. Sections were washed and incubated with Labeling Buffer for 5 min. The sections were washed again and incubated with Labeling Reaction Mix for 1 h at 37°C and washed before incubation with Stop Buffer for 5 min. After washing, the sections were incubated with streptavidin–HRP solution for 10 min at 37°C, and after additional washing, with Blue Label Solution for 3 min. Finally, the sections were counterstained with Nuclear Fast Red for 1 min and dehydrated before mounting.
Results
The radioimmunoconjugate
The immunoreactivity was determined to 0.8 and the average number of DOTA molecules per BR96 antibody was 2.6. The radiochemical purity of 177Lu-DOTA-BR96 was analyzed using HPLC and found to be 96.2% and there was 3.8% radiolabeled aggregation.
Tumor sampling
All 38 rats developed local tumors between the abdominal wall and peritoneum before the day of treatment (12–13 days after cell inoculation). The mean tumor volume on the day of treatment was 839 mm3 (standard deviation (SD) = 339 mm3). Scar tissue from tumors was collected from rats showing CR on day 4 (one rat), day 6 (one rat), and day 8 (three rats). The rapid treatment response resulted in four additional tumors that could only be used for cytokine analysis (one from day 5 and three from day 8) due to the small size of the tumor.
Cytokine evaluation
A total of 18 of the 29 cytokines evaluated were detected in the tumor samples, listed together with their functions in Table 1. The relative amount of interleukin (IL)-1α, IL-1β, and IL-1ra increased the first days after treatment, while the cytokine-induced neutrophil chemoattractant (CINC)-2 level was elevated between 3 and 6 days p.i., as seen in Figure 1. RANTES initially decreased, with nadir on day 4, before recovering to initial levels. Vascular endothelial growth factor (VEGF), CINC-1, LIX, and macrophage inflammatory protein (MIP)-3α decreased on day 8. CINC-3 and ciliary neurotrophic factor (CNTF) were only detected in a few samples. The relative amounts of the other cytokines detected did not change after treatment with 177Lu-mAb.
Cytokines present in tumors, cytokine function, and response to ionizing radiation.

CXCL: Chemokine (C-X-C motif) ligand; sICAM: soluble intercellular adhesion molecule-1; IL: interleukin; TIMP: tissue inhibitors of metalloproteinases; VEGF: Vascular endothelial growth factor.
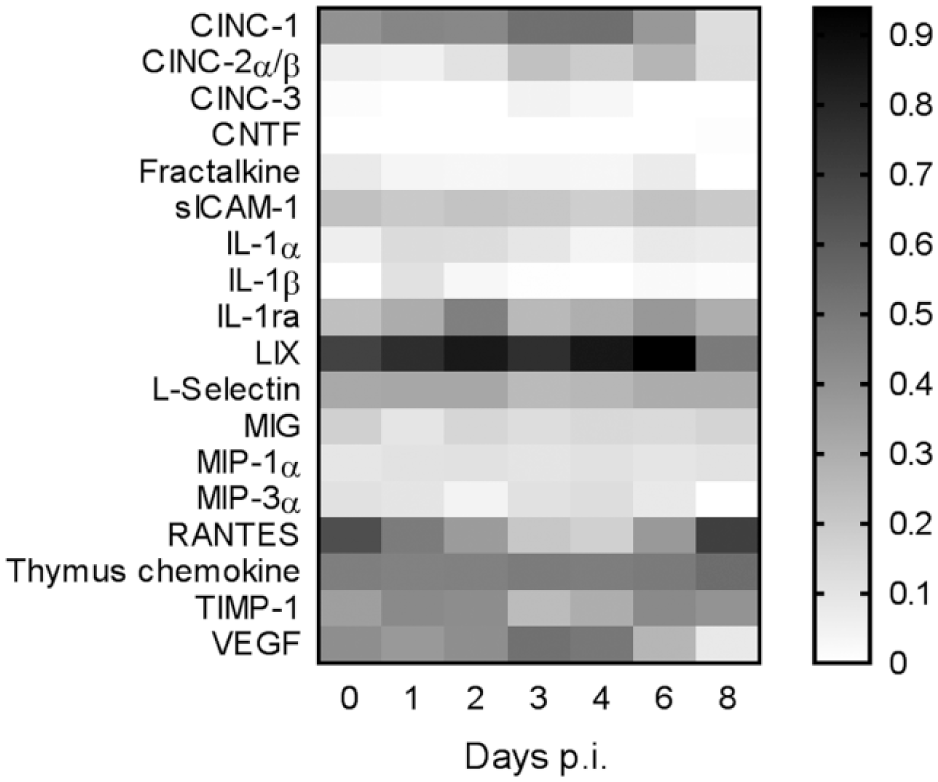
Change of relative amount of detected cytokines. All values are normalized to reference spots.
The cytokines not detected and their function are presented in Table 2. Neither interferon (IFN)-γ nor tumor necrosis factor (TNF)-α was detected. Only minor differences were seen when comparing remaining tumor and scar tissue from day 8 after treatment. Representative cytokine membranes from untreated tumor tissue, RIT-treated tumors, and scar tissue are shown in Figure 2.
Cytokines not detected in tumors, cytokine function, and response to ionizing radiation.

GM-CSF: granulocyte-macrophage colony-stimulating factor; IFN: interferon; IL: interleukin; TNF: tumor necrosis factor; NK cells: natural killer cells.

Representative cytokine membranes from an (a) untreated tumor, (b and c) RIT-treated tumors, and (d) scar tissue.
Biomarkers within the tumor
Untreated tumors consisted of dense growth of tumor cells and often areas of necrosis in the central part of the tumor (Figure 3(a)). DNA fragmentation was detected (TUNEL staining) in a few cells distributed randomly over the section and in close proximity to necrosis. After the injection of 177Lu-BR96, ongoing cell death was seen in the peripheral parts of tumor cell areas 1 day p.i., both as single cells and in small clusters. Cell death was mainly found in the same areas as the mAb BR96 (Figure 3(b)). An increase in granulation tissue was also seen as a result of the treatment. These effects were enhanced 2 days p.i. The granulation tissue increased over time, while DNA fragmentation either remained at the same level or decreased to the background level seen in untreated tumors. The few tumors that were excised 8 days p.i. and were sectioned and stained consisted mainly of granulation tissue and showed ongoing cell death in tumor cell areas (Figure 3(c)).

Tumor histology and distribution of therapeutic antibody 177Lu-BR96. (a) Untreated tumor tissue consisted of dense tumor cell growth. (b) Ongoing cell death was seen in areas with targeting antibody 1 day p.i. (c) Remaining tumors consisted mainly of granulation tissue 8 days p.i. Scale bar denotes 50 µm.
Discussion
The tumor immune contexture impacts the prognosis as seen in patients with colorectal cancer, and tumors can be divided into three groups based on this finding: (1) “immunogenic tumors” with anti-tumor immune cells result in a good prognosis, (2) “inflammatory tumors” with production of angiogenic and immunosuppressive molecules result in poor prognosis, and (3) “escaping tumors” with downregulation of antigen-presenting molecules leading to avoidance of an immune attack result in intermediate prognosis. 19 The tumor microenvironment contains a large variety of cells, and cytokines can be secreted by all cells present in the tumors, including cancer-associated fibroblasts. 20 Cytokines in the tumor microenvironment plays an important part in stimulating tumor growth and orchestrating the immune contexture. 4 These can be divided into functional groups: pro-inflammatory (TNF-α, IL-1α/β, and IL-17), anti-inflammatory (IL-4, IL-10, and transforming growth factor (TGF)-β), angiogenic (VEGF, TNF-α, and fibroblast growth factor (FGF)), pro-fibrotic (IL-6 and TGF-β), immune (IL-2, IL-4, and IL-7), and hematopoietic (colony-stimulating factor 1 (CSF1), granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-3, and erythropoietin (EPO)). 4 Some cytokines have been shown to have dual roles depending on dose and other cytokines present in the tissue,4,21 adding to the complexity of the cytokine environment and making the assessment of their true function difficult.
Cytokine analysis showed that 18 of the 29 cytokines evaluated were detected in the tumor samples. These were mainly involved in processes of infiltration of immune cells and angiogenesis. The presence of chemokines promoting the recruitment of leukocytes corresponded with earlier studies showing infiltration of neutrophils, 9 T cells, and macrophages 10 in this tumor model. The cytokine pattern was considered to be neither pro-inflammatory (described as consisting of IL-6, IL-17, IFN-γ, TNF-α, IL-1α/β among others) nor anti-inflammatory (containing cytokines like IL-1ra, IL-14, IL-5, IL-10, IL-11, IL-13). 4 Simultaneous immune stimulation and suppression has been seen in patients with advanced-stage tumors (consisting of increased levels of macrophage migration inhibitory factor (MIF), TNF-α, IL-6, IL-8, IL-10, IL-18 and TGF-β, 49 which has been interpreted as an inflammatory environment with infiltrating TH17 cells that are linked to chronic inflammation and autoimmune disease. However, this pattern does not correspond to the detected cytokines in this study.
The differences in cytokine content between untreated and RIT-treated tumors were small, indicating that the cytokine profile of untreated tumors was not transformed during tumor regression induced by RIT, especially since other cytokines present are thought to be involved in the same processes, that is, chemoattraction of immune cells and angiogenesis. RANTES was the chemokine with the most pronounced decrease after RIT, thus indicating a reduction in infiltration of T cells and macrophages. However, other chemokines with proposed similar functions were unaffected. Thus, it is not possible to draw any detailed conclusions until the roles of cytokines have been further specified. IFN-γ and TNF-α have been shown to promote the immune response of radiotherapy, 5 and thus the lack of both of these cytokines after RIT implicates that no such immune stimulation was initiated.
The cytokine array was chosen as it enabled simultaneous screening of many cytokines using a small amount of tumor sample; however, it does not provide any information on concentration. Thus, although the analysis showed higher density of IL-1ra than IL-1α and IL-1β, it is not clear whether the levels are high enough to block signaling, especially since abundant levels of IL-1ra are needed to functionally inhibit the signaling. 58
The histological evaluation demonstrated ongoing cell death in peripheral parts of the tumor, while central parts remained unaffected, as seen previously in another study with the same tumor model. 9 Staining of the injected mAb in this study followed the same pattern as seen previously when evaluated by autoradiography and could thus be used as a marker for radioactivity when autoradiography cannot be performed. The response to RIT in the tumor microenvironment would most likely follow the same heterogeneous pattern as the activity, implying that local changes in cytokine expression can be at least partly masked due to the homogenization of the tumor samples for cytokine analysis. In addition, at later time points in this study, there is a risk of selection of tumors that do not respond or respond slower to RIT than the tumors that have been rejected.
Radiation has been shown to be able to change cytokine expression, which could lead to remodeling of the stromal and vascular tissue, and changes in the function of the immune cells present in the tumor. 59 Both the innate and adaptive arms of the immune system can be triggered by radiation, but the optimal radiation regimen to activate the immune response remains to be defined. 32 Studies of tumor cell lines indicate that the secretion of cytokines after radiation seems to be cell-line specific. 60 In the in vivo situation, the tumor contains different tumor cell clones and normal cell types, making it difficult to compare the results of in vitro and in vivo studies. In addition, studies of external beam radiotherapy have shown that different radiation doses induce different cytokine profiles,4–6 without any linear correlation. 4 For example, TH1- and TH2-related cytokines decreased after a low absorbed dose, but increased after a high absorbed dose. 61 A local high dose (20 Gy) has also been shown to trigger the production of type I IFNs and upregulate CXCL9 and CXCL10, 5 leading to recruitment of cytotoxic T cells. Low-dose irradiation (<2 Gy) tends to promote anti-inflammatory responses associated with reduced production of NO and IL-1β and an increase in IL-10 and TGF-β. 5 However, these patterns were not found in our tumor model, in which the expression of CXCL9 did not change after irradiation and neither CXCL10 nor IL-10 was detected (TGF-β was not included in the cytokine array). The expected mean absorbed dose rate in this study is in the range 0.2–0.6 Gy/h at 24-h p.i. 9 and the total dose >25 Gy (based on unpublished data). However, since dose rates and particle energy differ between external beam radiotherapy and radionuclide therapy such as RIT, it is difficult to make any general comparisons between the two radiation modalities. It might be that the low dose rate cannot trigger an immune response or that the response is delayed due to the slow distribution of the radiolabeled mAb and the low dose rate. In the latter case, the lack of significant differences in cytokine expression between untreated and treated tumors in this study could be due to the short follow-up time. However, a longer follow-up time was not possible due to the rapid tumor regression after RIT in this syngeneic rat model.
Conclusion
The cytokine profile of untreated tumors was not transformed during tumor regression by RIT with 177Lu in this colon carcinoma model.
Footnotes
Acknowledgements
The authors would like to thank Dr Peter Senter (Seattle Genetics Inc.) for kindly providing the monoclonal antibody BR96.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed. This article does not contain any study with human participants performed by any of the authors.
Funding
This study was funded by the Swedish Cancer Society, Mrs Berta Kamprad’s Foundation, Governmental Founding of Clinical Research within the Swedish National Health Service, King Gustaf V’s Jubilee Foundation, The Lund University Medical Faculty Foundation, and The Lund University Hospital Fund.