
Abstract
Lactate dehydrogenase C is an isoenzyme of lactate dehydrogenase and a member of the cancer–testis antigens family. In this study, we aimed to investigate the expression and functional role of lactate dehydrogenase C and its basic mechanisms in renal cell carcinoma. First, a total of 133 cases of renal cell carcinoma samples were analysed in a tissue microarray, and Kaplan–Meier survival curve analyses were performed to investigate the correlation between lactate dehydrogenase C expression and renal cell carcinoma progression. Lactate dehydrogenase C protein levels and messenger RNA levels were significantly upregulated in renal cell carcinoma tissues, and the patients with positive lactate dehydrogenase C expression had a shorter progression-free survival, indicating the oncogenic role of lactate dehydrogenase C in renal cell carcinoma. In addition, further cytological experiments demonstrated that lactate dehydrogenase C could prompt renal cell carcinoma cells to produce lactate, and increase metastatic and invasive potential of renal cell carcinoma cells. Furthermore, lactate dehydrogenase C could induce the epithelial–mesenchymal transition process and matrix metalloproteinase-9 expression. In summary, these findings showed lactate dehydrogenase C was associated with poor prognosis in renal cell carcinoma and played a pivotal role in the migration and invasion of renal cell carcinoma cells. Lactate dehydrogenase C may act as a novel biomarker for renal cell carcinoma progression and a potential therapeutic target for the treatment of renal cell carcinoma.
Keywords
Introduction
Renal cell carcinoma (RCC) is the most common type of malignant kidney tumour with an incidence that continues to rise. Clear cell renal cell carcinoma (ccRCC), which comprises 83% of RCC, is insensitive to conventional chemotherapy and radiation therapy. 1 Although surgery can cure early-stage RCC, postoperative recurrence and metastasis are common. In addition, molecular targeting agents are the prior therapy form when RCC develops into late clinical stage. 2 Therefore, the early diagnosis of RCC and identification of effective targets for treatment are becoming extremely necessary. Cancer–testis antigens (CTAs) are expressed only in the germinal epithelium of the testis and in various malignant tumours, which are silenced in the somatic tissues. 3 Recent articles have proposed that CTAs play critical roles in various malignant cancer cells. 4 Because of these specific characters, CTAs are considered promising targets for early diagnosis and immunotherapy for cancers. However, the potential that RCC patients might benefit from CTA-targeted therapies remains to be evaluated.
Lactate dehydrogenase (LDH) is an important glycolytic enzyme that catalyses the transition of pyruvate to
The epithelial–mesenchymal transition (EMT) has been illustrated as potentially playing a pivotal role in RCC like other cancers.15–18 EMT is a transdifferentiation process in cancer cells that causes adherent epithelial cells to convert to motile mesenchymal cells and increases cell motility,19,20 thus increasing cancer cells’ metastatic potential. It was reported that the loss of E-cadherin and the increased expression of N-cadherin are associated with tumour progression and metastasis.21,22 Vimentin is also regarded as a marker of the EMT that was upregulated during this process. 23
In this study, to determine the functional role of LDHC in RCC cells, we utilized LDHC overexpression plasmids and small interfering RNAs (siRNAs) to overexpress or silence LDHC in RCC cells and investigated the potential of LDHC on RCC metastasis. Furthermore, we also found that LDHC induced EMT in RCC cells. All these results suggested that LDHC may be a novel biomarker and a potential therapeutic target for the treatment of RCC.
Materials and methods
Patients and Tissue Microarrays
In total, 133 RCC tissues were obtained from patients who were treated by partial or radical nephrectomy between 2008 and 2011 at the First Affiliated Hospital of Nanjing Medical University (Nanjing, China). All patients were recruited following informed consent. The clinical information of the 133 patients included in these tissue microarrays (TMAs) and their pathological characteristics were collected. The follow-up deadline was April 2016. The protocols used in the study were approved by the ethics committee of the hospital. From each RCC tissue, triplicate tissue cores with diameters of 0.6 mm were represented. This study was approved by the medical ethics committee of the hospital.
Immunohistochemistry
Serial sections from TMA blocks were deparaffinized in xylene and rehydrated through an ethanol gradient, and then were blocked in hydrogen peroxide in methanol for 10 min. Antigen retrieval was performed by incubation for 2 min in a steam pressure cooker containing citrate buffer 10 mM, pH 6.0. Then, samples were blocked for 5 min and incubated overnight with primary antibodies against LDHC (Abcam, Cambridge, UK; 1:100) at 4°C. After having been washed with phosphate-buffered saline (PBS) for 10 min, slides were cultured in the secondary antibody for 30 min. After a 10-min wash in PBS, the antibody reaction was visualized with a fresh substrate solution containing diaminobenzidine (DAB). The sections were counterstained with haematoxylin, dehydrated and coverslipped.
Evaluation of staining
The immunohistochemical staining was assessed by two experienced pathologists without knowledge of the clinical data separately. To evaluate the expression of LDHC in RCC tissues, a semi-quantitative scoring system (0–3) was used based on the staining intensity of the tumour tissue: 0 = negative, 1 = weak positive, 2 = moderate positive, and 3 = strong positive. LDHC positive refers to scores 1–3 and LDHC negative refers to score 0.
Cell culture and human clinical samples
The human skin metastasis renal clear cell carcinoma cell (CAKI-1), human renal clear cell cancer cell (CAKI-2) and human renal tubular epithelial cell (HK-2) were purchased from the Chinese Academy of Sciences (Shanghai, China). CAKI-1 and CAKI-2 were cultured in McCoy’s 5A Medium (Gibco, Waltham, MA, USA) and HK-2 was cultured in Dulbecco’s modified Eagle’s medium (DMEM) Medium (Gibco Waltham, MA, USA). The medium was supplemented with 10% foetal bovine serum (FBS; Gibco) in a humidified air atmosphere at 37°C and 5% CO2.
Eighteen pairs of ccRCC and corresponding adjacent renal tissues were obtained from patients who underwent partial or radical nephrectomy with informed consent at First Affiliated Hospital of Nanjing Medical University. Their histologic characteristics were confirmed by the pathologist.
RNA extraction, RT-PCR and quantitative RT-PCR
The RNA was extracted from cell lines and tumour samples using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Total RNA was reverse transcribed into complementary DNA (cDNA) using PrimeScript RT Master Mix (TaKaRa, Kyoto, Japan). Reverse transcription polymerase chain reaction (RT-PCR) was performed using GoTaq Green Master Mix (Promega, Fitchburg, WI, USA) according to the manufacturer’s instructions. The PCR products were analysed on 1% agarose gel and visualized. Quantitative RT-PCR was performed using FastStart Universal SYBR Green Master (Roche, Branford, CT, USA). β-actin was used as internal controls for LDHC. For PCR, we used the following primers (Realgene, Nanjing, China): LDHC forward, 5′-TCCCTGTGGGTCATCTGTACT-3′, and reverse, 5′-GGGCAAGTTCATCAGCCAAA-3′; and β-actin forward, 5′-TGACGGGGTCACCCACACTGTGCCCATCTA-3′, and reverse, 5′-CTAGAAGCATTTGCGGTGGACGATGGA GGG-3′.
Construction of LDHC overexpression plasmids and siRNA synthesis and cell transfection
LDHC overexpression plasmids were conducted and purchased from GenePharma (Shanghai, China). In addition, two siRNAs and one scrambled siRNA (negative control) were designed and chemically synthesized by GenePharma. The two sequences of siRNA for LDHC were as follows: 5′-GGCAUGGCUUGUGCUAUUATT-3′ and 5′-GUGGCUAUAAUGAAAUCAATT-3′. The sequence of scrambled siRNA that did not cause specific degradation of any known cellular messenger RNA (mRNA) was 5′-UUCUCCGAACGUGUCACGUTT-3′. RCC cell lines were seeded onto the six-well plates in McCoy’s 5A (m5A) medium without antibiotics in three groups (negative control, siRNA1 and siRNA2). Once the cells were 50%–70% confluent, CAKI-2 cells were transfected with LDHC-siRNA or negative control, and CAKI-1 cells were transfected with LDHC overexpression plasmids or vector controls, using Lipofectamine 2000 (Invitrogen) and opti-MEM medium (Gibco, Carlsbad, MA), according to the manufacture’s instructions. To alleviate toxicity, the medium surrounding the cells was replaced with m5A 4–6 h after transfection.
Lactate concentration determination assay
After transfection for 48 h, cells in the six wells were washed twice with PBS and each well was replaced with fresh m5A medium. After 24 h of incubation, the medium surrounding the cells was collected and centrifuged to remove fragments of cells. The concentration of the lactate in each group was determined using a Lactic Acid Assay Kit (Jiancheng, Nanjing, China). Meanwhile, the cells in each well were collected and counted using cell counting chamber. The lactate concentration per 105 cells was lactate concentration divided by cell number and multiplied by 105.
Wound healing assay
RCC cell lines were seeded onto the six-well plates and were grown to confluence of 80%–90%. After transfection for 48 h, the confluent monolayer was scratched with a 100 µL pipette tip. Then, the scratched cells were washed three times with PBS and replaced with serum-free m5A medium. The wounded monolayer was observed and photographed under a light microscope. Percentage of cell-free area was calculated as (cell-free area48h / cell-free area0h) × 100, and cell migration rate was displayed as the percentage of cell covered area (100% cell-free area). Each experiment was performed in triplicate.
Transwell migration and invasion assays
The in vitro migration and invasion assays were carried out using 24-well chambers with 8 µm pore size polyethylene terephthalate (PET) track-etched membranes (Corning, Inc., Corning, NY, USA). A total of 5 × 104 transfected cells were suspended in the upper chamber with serum-free medium, while the lower chamber was filled with 20% FBS-containing medium. In the invasion assays, the membranes were coated with Matrigel (BD Biosciences, San Diego, CA, USA) to form matrix barriers. After incubation at 37°C for 48 h (migration assay) or 72 h (invasion assay), the cells on the upper surface were removed by a cotton swab, while the cells invaded on the bottom membranes were fixed in 4% methanol solution and stained with 0.5% crystal violet and counted under a microscope.
Western blot assay
After 48 h of transfection, cells were washed twice with PBS and dissolved in protein lysis buffer (Beyotime Institute of Biotechnology, Nantong, Jiangsu, China). The cell supernatants were extracted after centrifugation for 15 min at 14,000 r/min. The concentration of the protein was determined using a BCA Protein Quantification Kit (Beyotime Institute of Biotechnology). Equal amounts of protein were subjected to electrophoresis on a 10% sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS-PAGE) gel. The protein was transferred to polyvinylidene difluoride (PVDF) membranes in transfer buffer at 300 mA for 2 h. The membranes were incubated with Tris-buffered saline (TBS) containing 5% non-fat milk powder for 2 h at 4°C. The membranes were then washed three times with TBS containing 0.2% Tween-20 (TBST). The membranes were incubated in an antibody solution containing the primary antibodies at 4°C overnight. After that, the membranes were washed three times with TBST and incubated in a secondary antibody solution for 2 h at room temperature. The indicated proteins were detected by the enhanced chemiluminescence western blotting detection system.
Statistical analyses
The associations between protein expression values and clinicopathological factors were assessed using the chi-square test. We used the Kaplan–Meier curve and log-rank tests to assess the association of LDHC expression with overall survival. Results were expressed as mean ± standard deviation (SD). Differences between two or three groups were compared with Student’s t-test. A value of p < 0.05 or less was considered statistically significant. All of the statistical calculations were performed using SPSS software (Version 13.0 SPSS, Inc., Chicago and Delaware).
Results
LDHC is upregulated in RCC clinical specimens
We examined LDHC mRNA expression levels in 18 pairs of frozen samples from ccRCC patients (18 cancers and 18 corresponding adjacent tissues) by RT-PCR. Our results showed that 4 of the 18 (22.2%) tumour tissues aberrantly expressed LDHC, while none of adjacent tissues expressed LDHC (Figure 1(a)). To further determine the expression of LDHC protein in renal tissues, we examined the expression of LDHC protein in 133 RCC TMAs by immunohistochemical analysis. As shown in Figure 1(b), LDHC was mainly expressed in the cytoplasm of cancer cells. Table 1 showed 27 of the 133 tumour tissues expressed LDHC protein. We found LDHC positive expression was significantly associated with advanced tumour stage (p = 0.042) and survival (p = 0.011). However, no significant differences were revealed between negative and positive expression of LDHC in age, gender, tumour size, histology and histological grade. In addition, univariate Kaplan–Meier/log-rank analysis also indicated that positive LDHC protein expression was significantly associated with an increased risk for poor clinical prognosis in RCC patients (log rank p = 0.0043; Figure 1(c)). Patients with positive LDHC expression had a shorter overall survival. These results suggest that positive LDHC expression may act as an important prognostic biomarker for RCC.

LDHC expression in RCC tissues correlates to the poor prognosis. (a) RT-PCR was used to evaluate the expression of LDHC in RCC tissues. β-actin was used as an internal control. (b) Immunohistochemical images of LDHC expression in RCC TMAs. (c) Kaplan–Meier/log-rank analysis for overall survival from 133 PCa patients according to LDHC expression in tumour tissues.
Relationship of LDHC expression and clinicopathological characteristics of the 133 RCC patients involved in this study.
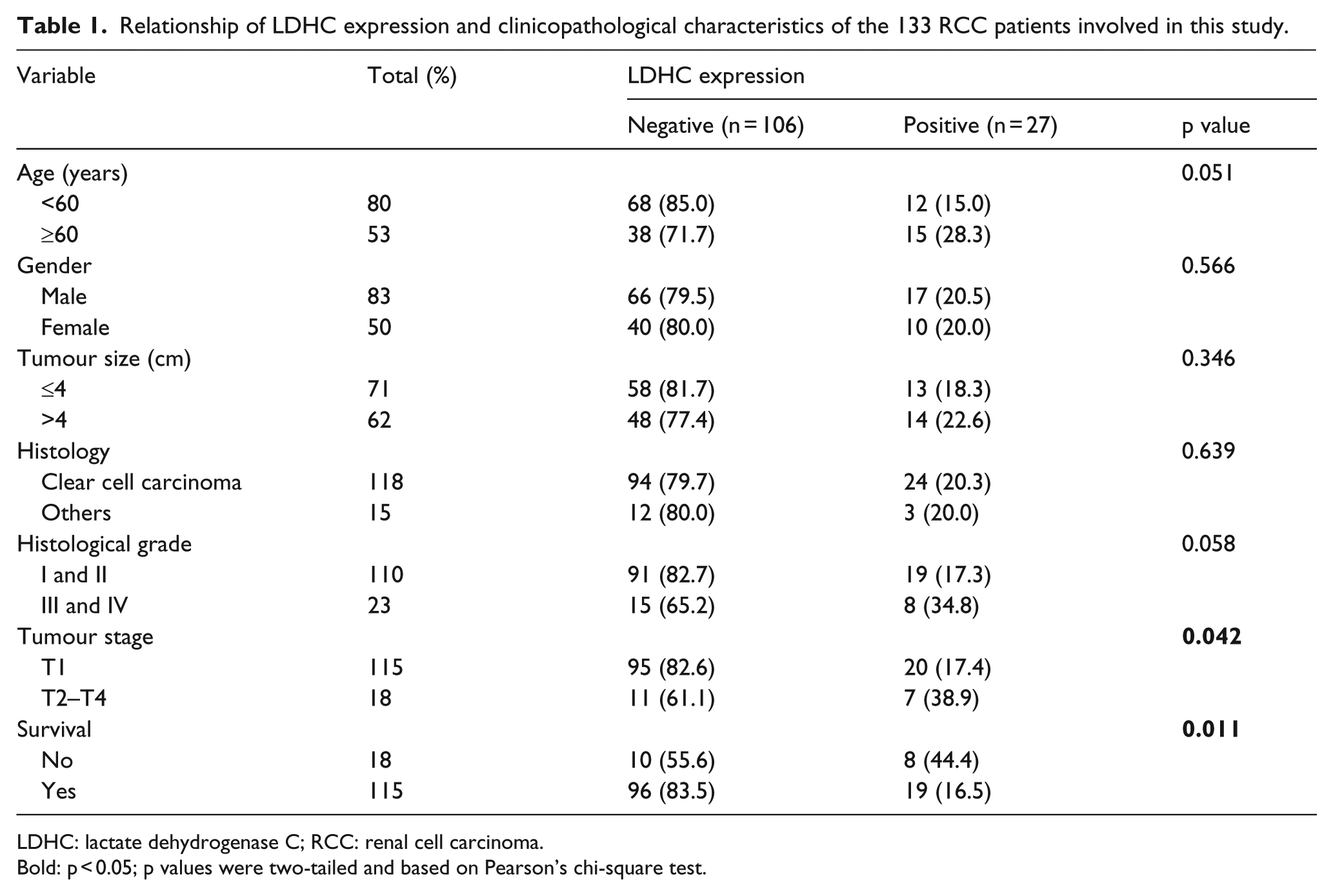
LDHC: lactate dehydrogenase C; RCC: renal cell carcinoma.
Bold: p < 0.05; p values were two-tailed and based on Pearson’s chi-square test.
LDHC is expressed in RCC cell lines
We used quantitative RT-PCR and western blot assay to determine whether LDHC was expressed in RCC cell lines (CAKI-1 and CAKI-2) and renal tubular epithelial cell (HK-2). As displayed in Figure 2(a) and (b), the mRNA and protein expression level of LDHC was highest in CAKI-2, while it was relatively low in CAKI-1. The renal tubular epithelial cell HK-2 hardly expressed LDHC. Furthermore, to study the functional role of LDHC in RCC cells, LDHC was blocked down in CAKI-2 by using siRNAs and overexpressed in CAKI-1 by transfecting overexpression plasmids. LDHC mRNA and protein expression were significantly downregulated in CAKI-2 (Figure 2(c) and (e)) and markedly upregulated in CAKI-1 (Figure 2(d) and (f)) after transfection.

LDHC expression in RCC cell lines and the transfection efficiency of siRNA and expressed plasmids. The expression of (a) LDHC mRNA and (b) protein level in RCC cells (CAKI-1 and CAKI-2) and renal tubular epithelial cells (HK-2). The expression of (c) LDHC mRNA and (e) protein level in CAKI-2 cells after transfection with siRNAs. β-actin was used as an internal control. The expression of (d) LDHC mRNA and (f) protein level in CAKI-1 cells after transfection with overexpressed plasmid. β-actin was used as an internal control. All data are presented as mean ± SD.
LDHC induces RCC cells to produce lactate
We performed lactate concentration determination assay in CAKI-1 and CAKI-2 to assess the role of LDHC in RCC lactate metabolism. As shown in Figure 3, the lactate concentration was significantly reduced after blocking the endogenous LDHC in CAKI-2. By contrast, overexpression of LDHC increased the lactate concentration in CAKI-1. This finding indicated that LDHC may play an important role in lactate metabolism of RCC cells.

Effect of LDHC knockdown or overexpression on lactate metabolism of RCC cells. (a) The lactate concentration of CAKI-2 cells with knockdown of LDHC. (b) The lactate concentration of CAKI-1 with LDHC overexpression.
LDHC is involved in RCC cells migration and invasion
Wound healing assay and transwell assay were applied in CAKI-2 and CAKI-1 to study the significance of LDHC during tumour metastasis. As shown in Figure 4(a), transwell assays revealed that knockdown of endogenous LDHC significantly inhibited the migration and invasion of CAKI-2 cells. Conversely, overexpression of LDHC enhanced the migration and invasion of CAKI-1 cells (Figure 4(b)). As shown in Figure 4(c) and (d), wound healing assays also demonstrated that low expression of LDHC reduced migration rate of CAKI-2, while overexpression of LDHC increased migration rate of CAKI-1. These results suggested that LDHC may promote RCC cells metastasis.

Knocking down the expression of LDHC inhibited the migration and invasion, while overexpression of LDHC promoted the migration and invasion of RCC cells. (a) Transwell assays to analyse the effects of LDHC knockdown on the migration and invasion of CAKI-2 cells. (b) Transwell assays to analyse the effects of LDHC overexpression on the migration and invasion of CAKI-1 cells. (c) Wound healing assays to analyse the effect of LDHC knockdown on the migration of CAKI-2 cells. (d) Wound healing assays to analyse the effect of LDHC overexpression on the migration of CAKI-1 cells.
LDHC induces EMT and increases the expression level of MMP-9
As previously described, tumour metastasis is closely associated with the activation of EMT. To examine whether LDHC induces EMT in RCC cells, western blot assay was performed to assess the EMT-associated protein expression. As shown in Figure 5, knockdown of LDHC in CAKI-2 significantly induced the expression of E-cadherin and reduced N-cadherin and vimentin. On the contrary, upregulation of LDHC in CAKI-1 reduced the expression of E-cadherin and increased N-cadherin and vimentin. The results demonstrated that LDHC may induce the EMT process in RCC.
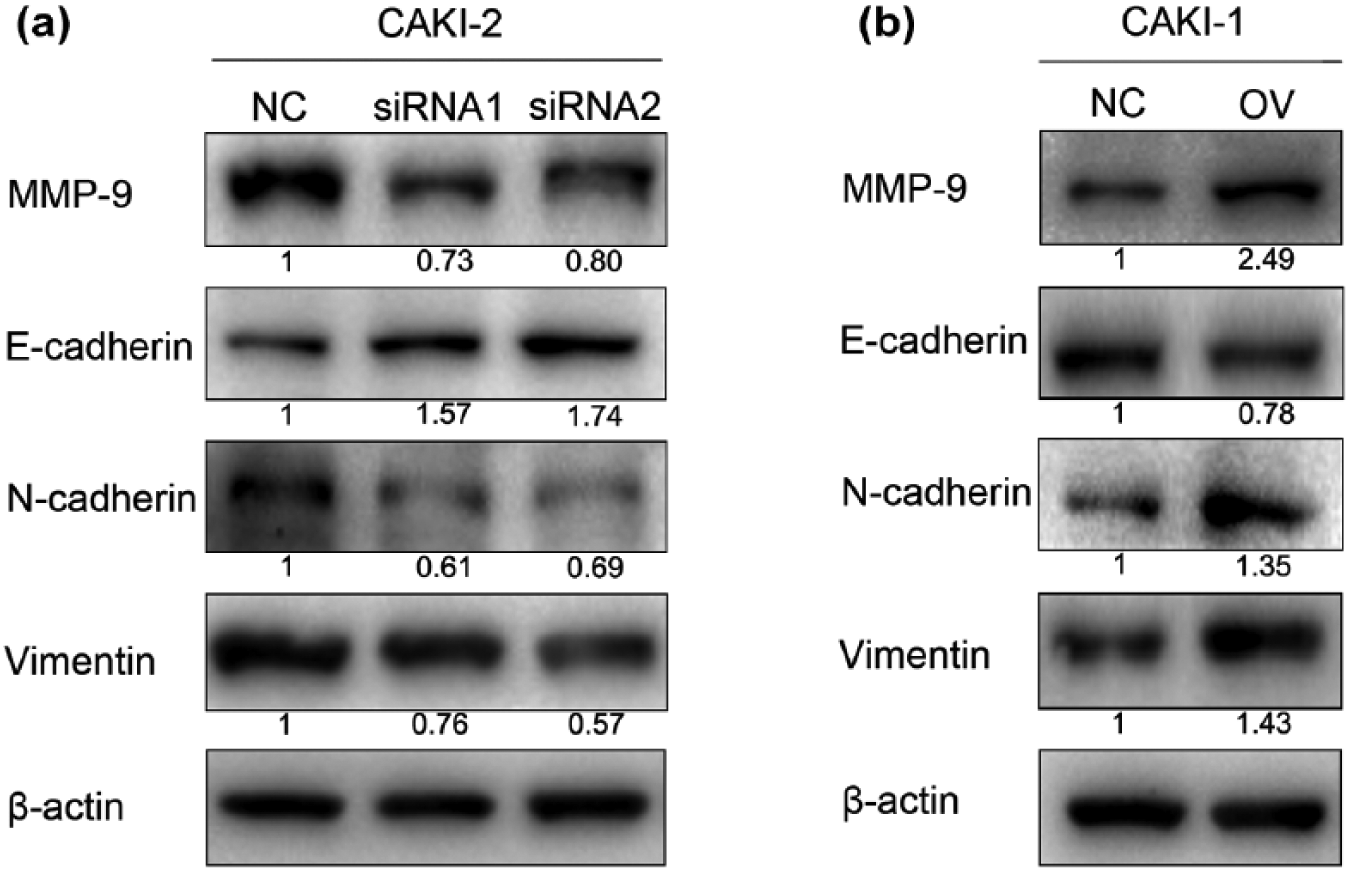
Effect of LDHC knockdown or overexpression on EMT and MMP-9 in CAKI-1 and CAKI-2. (a) N-cadherin, vimentin and MMP-9 were reduced, while E-cadherin was induced after knockdown of LDHC in CAKI-2. (b) N-cadherin, vimentin and MMP-9 were induced, while E-cadherin was reduced after overexpression of LDHC in CAKI-1. β-actin was used as an internal control.
In addition, the expression level of matrix metalloproteinase-9 (MMP-9) in CAKI-2 was markedly decreased following treatment of siRNA, and it was upregulated in CAKI-1 after transfection with LDHC plasmids.
Discussion
LDHC is the first testis-specific isozyme discovered in male germ cells, and it is essential for lactate metabolism during spermatogenesis. Compared with LDHA and LDHB, LDHC has high heat stability properties and has a broader range of substrates.8,24 Recently, researchers have begun to pay attention to the significance of LDHC in tumours. Although previous studies showed that LDHC is expressed in various tumours, the expression and the function of LDHC in RCC have not been reported. Our study revealed that LDHC expression could be an important indicator of RCC progression and involved in RCC metastasis.
In this study, we demonstrated that LDHC was not expressed in normal renal tissues but was aberrantly expressed in RCC tissues. We also identified LDHC was expressed in RCC cell lines, especially in CAKI-2. More notably, positive LDHC expression showed a significant correlation with high tumour stage and poor prognosis. Moreover, to investigate the potential biological effect of LDHC on RCC cytological behaviours, two independent siRNAs effectively downregulated the endogenous expression of LDHC in CAKI-2, and the overexpressed plasmid significantly increases LDHC expression in CAKI-1. Our data revealed that LDHC played an essential role in lactate metabolism of RCC cells, and this gene could positively regulate the ability of migration and invasion in RCC cells. In conclusion, we suggested that the aberrant expression of LDHC may act as a novel prognostic and therapeutic target for treating RCC.
This study preliminarily revealed the potential functional role of LDHC in RCC progression and metastasis. Even under well-oxygenated conditions, cancer cells gain a large amount of energy and produce a large amount of lactate through aerobic glycolysis, which is named the Warburg effect.25,26 The Warburg effect was identified as a common phenomenon in most cancers, and it was related with tumour proliferation and provided suitable conditions for tumour metastasis.27–29 Our data showed that LDHC could prompt RCC cells to produce lactate. Therefore, we suggested the expression of LDHC might accelerate RCC cells metastasis by promoting aerobic glycolysis, 30 and provided acid tumour microenvironment which was suitable for local invasive growth and metastasis.31,32 In addition, previous studies reported that lactate itself could induce the migration of cancer cells through multiple molecular mechanisms.33,34
Our data also suggested that LDHC could induce EMT process in RCC cells. EMT is a complicated multistep process, which has been shown to induce fully differentiated cells to lose their cell polarity and cell adhesion features and to obtain an invasive mesenchymal phenotype. Recent studies have showed that EMT acts a pivotal role in RCC pathogenesis, tumour aggressiveness and therapeutic effect.35,36 EMT is also correlated with an advanced recurrence risk and poor overall survival in patients with RCC. 37 Moreover, our data demonstrated that LDHC could upregulate the expression of MMP-9. This protein was a member of the family of zinc-dependent endopeptidases, the central role of which was the degradation and remodelling of the extracellular matrix, leading to tumour invasion and metastasis. 38 Recent studies revealed that MMP-9 could participate in regulating various tumour metastasis and might be a novel direction for tumour treatment.39,40 In the progression of RCC, MMP-9 could be a critical molecule for invasion and metastasis and is closely associated with poor survival. 41 Our study suggested that LDHC might have a substantial correlation with EMT and increased the expression level of MMP-9, thus enhancing the migratory ability of RCC cells. However, the exact mechanisms by which LDHC induces EMT and MMP-9 expression need further investigation.
In conclusion, our results showed that positive LDHC expression was significantly associated with tumour stage in RCC. This study was the first to demonstrate that endogenous LDHC was expressed in CAKI-2 RCC cells and could play a crucial role in RCC migration and invasion. Further studies are necessary to elucidate the mechanism by which LDHC induces EMT and MMP-9 overexpression. In vivo studies are also recommended to obtain deeper understanding of the roles of LDHC.
Footnotes
Acknowledgements
Z.W. and C.Q. helped in the conception and design. Y.H., C.L., J.Z., C.M., Y.Y., A.X., P.L., S.L., J.Y., C.Q. and Z.W. contributed to the acquisition of data. J.Z. and M.B. helped in the analysis and interpretation of data. Y.H. and Z.W. assisted in drafting the manuscript. All authors approved the final version. Y.H., C.L. and J.Z. contributed equally to this study.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Ethics approval and consent to participate
All patients gave informed consent to the study (pathological characteristics and immunohistochemical staining of renal cell carcinoma (RCC) tissues obtained after curative resection and study of statistical associations with clinicopathological data). This study was approved by the medical ethics committee of the hospital and conducted in accordance with the Chinese laws and regulations.
Funding
The study was supported by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), Program for Provincial Initiative Program for Excellency Disciplines of Jiangsu Province, National Natural Science Foundation of China (81302217) and graduate international exchange and cooperation projects of Nanjing Medical University (201601C006) and Six talent peak project in Jiangsu province (WSN-011).