
Abstract
MicroRNAs are emergent players of epigenetics that function as oncogenes or tumor suppressors and that have been implicated in regulating diverse cellular pathways. MiR-10b is an oncogenic microRNA involved in tumor invasion and metastasis in various cancers. Our data have shown that miR-10b is overexpressed in colorectal cancer samples in comparison with non-tumorous adjacent mucosa (p = 0.0025) and that it is associated with severe features such as tumor size >5 cm (p = 0.023), distant metastasis (p = 0.0022), non-differentiated tumors (p = 0.016), and vascular invasion (p = 0.01). Regarding the regulation of its expression, positive correlation between the loss of miR-10b and aberrant DNA methylation (p = 0.02) as well as a loss of TWIST-1 messenger RNA (p = 0.018) have been observed. Furthermore, expression analysis of the downstream miR-10b targets has shown that there are associations between low HOXD10 messenger RNA and E-cadherin protein levels (p < 0.0001, p = 0.0008, respectively) and overexpression of miR-10b. Our data suggests that overexpression of miR-10b results from high levels of TWIST-1 and may induce a decrease of E-cadherin membranous protein levels, thus contributing to the acquisition of metastatic phenotypes in colorectal cancer.
Introduction
Colorectal cancer (CRC) is the third most diagnosed cancer with about 1.2 million new cancer cases and over a half million deaths per year. 1 In Tunisia, the incidence of CRC is 2.5–4.5/100000. 2 The prognosis for CRC has been improved over the past decade, especially by treatment advances; however, only 30%–40% of patients are diagnosed at early stages. 3 The progression of normal colonic mucosa to invasive cancer requires several molecular changes affecting oncogenes as well as tumor suppressor genes. 4 Although chromosomal and microsatellite instability pathways constitute the major genetic alteration events in CRC,5,6 the epigenetic alterations, such as DNA methylation, play a key role in the colorectal carcinogenesis. 7 In fact, aberrant methylation of cancer-related gene promoters is very frequent and is often responsible for transcriptional silencing of several genes in tumor tissues. 8
Latterly, many studies have highlighted the role of microRNAs (miRNAs), as major players of epigenetics, in the establishment and progression of CRC. MiRNAs are recently discovered class of abundant non-coding single-stranded RNA, which are small, endogenous, and regulatory. 9 They play a fundamental role in the modulation of messenger RNA (mRNA) translation and post-transcriptional regulation, mainly via RNA–RNA interaction. 10 In fact, miRNAs interfere with the target mRNAs, leading to translational repression or mRNA decay. 11 MiRNAs participate in various physiological and pathological processes, including CRC metastasis by causing many phenomena such as angiogenesis and epithelial–mesenchymal transition (EMT). 12 In CRC, several studies have reported that miRNAs are involved in tumor progression and metastasis and are therefore suggested as potential prognostic biomarkers.13,14
It has been demonstrated that the transcription factor TWIST-1 induces miR-10b which in turn inhibits the translation of a member of the homeobox (HOX) gene family, HOXD10. The expression of HOXD10 is lost during the malignant progression of breast cancer, resulting in the increased levels of the pro-metastatic gene RHOC. 15 The transcriptional silencing of miR-10b decreases its levels and boosts the levels of HOXD10, which leads to metastasis inhibition. 16 This miRNA has been previously found to promote tumor metastasis in many malignancies, such as breast, 17 oesophageal, 18 ovarian, 19 and lung cancers. 20 While most studies report the oncogenic effect of miR-10b in gastric cancer,21–23 some others show that it is downregulated mainly through CpG DNA methylation and that it may act as tumor suppressor gene in tumor tissues.24,25
Although recent studies have reported an overexpression of miR-10b in CRC, through targeting the HOXD10 translation and thus enhancing the RHOC expression, its effect on metastatic CRC remains unclear.26–28
In addition, it has been demonstrated that miR-10b exerts its metastatic effect on breast cancer and cell lines through targeting E-cadherin protein expression.29,30 Furthermore, miR-10b promotes the migration of mouse bone marrow-derived mesenchymal stem cells and downregulates the expression of E-cadherin. 31 More recently, it has been reported that miR-10b regulates EMT by modulating the expression of a bunch of proteins, such as E-cadherin, in breast cancer malignant tissues as well as laryngeal and nasopharyngeal carcinomas cell lines.32–34
This work aims to determine the expression level and the mechanisms involved in the regulation of miR-10b in Tunisian patients with CRC. We have focused on analyzing the TWIST-1 expression and on determining the methylation patterns of miR-10b promoter in tumor tissues. We have also investigated its downstream effects on HOXD10 and E-cadherin expression.
Materials and methods
Patients and specimens
Human clinical samples were collected between 2003 and 2009 from surgical specimens from 50 patients with CRC at the Department of Digestive Surgery of Habib Bourguiba University Hospital (Sfax, Tunisia). The corresponding adjacent non-neoplastic tissues from the macroscopic tumor margin were isolated at the same time from 10 samples and were used as controls. The study was performed in accordance with the ethical standards of the revised Declaration of Helsinki (October 2013), and all patients gave informed consent prior to specimen collection according to institutional guidelines. None of the patients had preoperative or postoperative chemotherapy. At the time of surgery, the age of patients ranged from 25 to 85 years (mean 63 years), and the sex ratio was 1:1. Tumors were staged according to the tumor-node-metastasis (TNM) (2010) classification criteria of the Union for International Cancer Control (UICC).
5-Azacytidine treatment of HT-29 cells
The HT-29 (American Type Culture Collection (ATCC)) colon cancer cell line was maintained in Dulbecco’s modified eagle medium (DMEM) supplemented with 10% fetal bovine serum, 1% kanamycin, streptomycin-penicillin, and incubated at 37°C and 5% CO2. Three replicative 150 mm cultures were treated with (a) dimethyl sulfoxide (vehicle alone, 0 µM 5-Azacytidine (5-Aza)), (b) 1 µM 5-Aza, (c) 5 µM 5-Aza, and (d) 10 µM 5-Aza for 5 days. These 5-Aza concentrations were similar and greater than the 5-Aza concentration previously reported to increase apoptosis and alter genome methylation as well as mRNA gene expression in HT-29 cells. 35 The HT-29 cells were washed with phosphate buffered saline (PBS) on the plate prior to scraping and centrifuging the cells.
RNA extraction and complementary DNA synthesis
Total RNA was isolated from tissues and cell lines by TRIzol reagent (Invitrogen) and conserved at −80°C until use. RNA concentration and purity were measured using the NanoDrop ND-1000 UV/Vis spectrophotometer (Thermofisher Scientific).
First strand complementary DNA (cDNA) synthesis was performed on 500 ng of total RNA, treated with DNaseI (Fermentas), in a final volume of 20 µL containing 0.1 pmol random hexamer primers, 10 nmol of each deoxynucleotide triphosphate (dNTP), 0.5 U/µL RNase inhibitor (Fermentas), 4 µL of 5× reverse transcriptase (RT) buffer (Invitrogen), and 200 U of moloney murine leukemia virus (MMLV) reverse transcriptase (Invitrogen). The reaction mixture was incubated at 37°C for 50 min, followed by 70°C for 10 min. The cDNA (2 µL) was used as a template in polymerase chain reaction (PCR) using specific primers for TWIST-1 and HOXD10, with RNA 28S and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as endogenous controls. The primer sequences, product sizes, and annealing temperatures are shown in Table 1. The PCR products were analyzed on 2.5% agarose gel, stained with ethidium bromide, and visualized under ultraviolet (UV) light.
Primer sequences and amplification conditions of miR-10b, U6, TWIST-1, HOXD10, RNA 28S, and GAPDH.

Stem-loop reverse transcription-quantitative polymerase chain reaction
MiRNA reverse transcription (RT) was performed as described previously. 36 In brief, miR-10b was reverse-transcribed using a stem-looped primer (5′-CTCAACTGGTGTCGT GGAGTCGGCAATTCAGTTGAGCACAAATT-3′) which binds to six nucleotides at the 3′ side of the mature miR-10b molecule. The small nuclear RNA (snRNA) U6 was also reverse-transcribed using U6-R specific primer (5′-CGCTTCACGAATTTGCGTGTCAT-3′) and considered as an internal control.
RT reaction was performed on 300 ng of total RNA in a final volume of 10 µL containing 0.1 pmol of specific primer, 0.5 unit/ µL RNase inhibitor (Fermentas), 2 µL of 5× RT buffer (Invitrogen), 10 nmol of each dNTP, and 100 U of MMLV reverse transcriptase (Invitrogen). The reaction was incubated for 30 min at 16°C followed by 50 min at 37°C, and then stopped by incubation for 15 min at 70°C. MiR-10b expression level was examined by RT-quantitative PCR (RT-qPCR) using U6 as an endogenous control. The reaction mixture contained 1 µL of RT product as template, 5 pmol of each primer (Table 1), and 10 µL of 2× SYBR Premix Ex Taq II (Tli RNaseH Plus) (TAKARA). The amplification conditions performed on Biorad-CFX96 were as follows: 95°C for 30 s, then 40 cycles of 94°C 10 s and 61°C 45 s. The specificity of the PCR products was confirmed by melting curve analysis, from 65 to 95°C, with a heating rate of 0.5°C/s.
DNA extraction and methylation specific PCR
Genomic DNA was isolated from tissues sections by phenol/chloroform procedure as described previously, 37 and the quantity was checked by NanoDrop ND-1000 UV/Vis spectrophotometer (Thermofisher Scientific). DNA samples were stored at −20°C for further use. For Methylation Specific PCR (MSP), 1µg of DNA was treated by sodium bisulfite which converts the unmethylated cytosine to uracil using the EZ Methylation Kit according to the manufacturer’s recommendations (ZymoResearch). The bisulfite-treated DNA was amplified using specific primers for methylated and unmethylated miR-10b alleles. 25 The MSP conditions were as follows: initial denaturation at 95°C for 5 min, 40 cycles of 30 s at 94°C, 30 s at 53°C for the unmethylated allele (U) and 50°C for the methylated allele (M), followed by 30 s at 72°C, and a final extension at 72°C for 10 min. The reactions were performed in a total volume of 25 µL, containing 2 µL of bisulfite-treated DNA, 0.2 µM of each primer, 200 µM dNTP, 1× PCR buffer, and 15 µL of ZymoTaq™ PreMix (ZymoResearch). The amplified products were analyzed by electrophoresis on 2.5% agarose gel, stained with ethidium bromide and visualized under ultraviolet light.
Immunohistochemical analysis of E-cadherin protein localization
The immunohistochemical (IHC) analysis of E-cadherin protein membranous location was described previously. 38 In brief, hematoxylin and eosin-stained slides were reviewed, and blocks containing adenocarcinoma were selected by the pathologists. IHC staining was performed using the streptavidin–biotin peroxidase system. Succinctly, E-cadherin antibody (Clone NCH-38, Dako) diluted at 1:400 was incubated for 30 min at room temperature, and then washed with PBS. Primary antibody binding was visualized with biotin-labeled secondary antibodies (goat anti-rabbit, Dako) and a streptavidin–peroxidase complex using diaminobenzidine as a chromogenic substrate (LSAB system, Dako). The immune-stained slides were scored as described previously. Briefly, the initial scoring was graded on the basis of the approximate percentage of positive tumor cells, and the relative immunostaining intensity was as follows: 0: no staining or less than 5%, 1: 5%–25% staining, 2: 26%–75% staining, and 3: 76%–100% staining. In addition, the intensity of staining was also evaluated as follows: 0: negative; 1: weak; 2: moderate; and 3: intense. The sum of the intensity and staining extent scores was used as the final immune-reactive score (0–12). The final scores were regarded as negative (0–1 score) and positive (2–12 score). Only membranous immunostaining was considered as positive.
Statistical analysis
Statistical analysis and graphs were performed using the GraphPad Prism 5.0 software. Gene expression ΔCt values of miRNA from each sample were calculated by normalizing according to internal control U6 expression, and relative quantification values were plotted. The non-parametric Mann–Whitney U test was used for the statistical evaluation of the differences between tumor and normal tissues groups, and correlation analyses were performed using Spearman’s correlation test. The analysis of miR-10b levels according to clinical parameters or expression status of TWIST-1, HOXD10, and E-cadherin was also performed using Mann–Whitney test.
Student’s t test was used to compare the RT-PCR data from HT-29 cells. Using Mann–Whitney test, we compared miR-10b expression levels in dead patients to the surviving ones. Also, the correlation with overall survival was performed using Kaplan–Meier survival plots, and the significance was tested with the log rank test.
The SPSS software Version 20 was used for the statistical analysis of the association between miR-10b methylation and clinicopathological features, and the correlations were assessed by chi-square test. Statistical significance was set up to p ≤ 0.05 in each test.
Results
Expression of miR-10b in primary CRC tissues and non-cancerous adjacent mucosa
In this study, we have analyzed the expression of miR-10b in 50 CRC patients and 10 non-tumorous control tissues. The results have shown that miR-10b is overexpressed in tumors compared to the control samples (p = 0.0025, Figure 1). Overexpression of miR-10b correlated with aggressive behavior such as size of tumors >5 cm (p = 0.023, Figure 2(a)), poor differentiated tumors (p = 0.016, Figure 2(b)), distant metastasis (p = 0.0022, Figure 2(c)), and vascular invasion (p = 0.01, Figure 2(d)). Our data suggest that the increased expression of miR-10b constitutes a marker of poor prognosis in CRC. Furthermore, the association of miR-10b with overall survival has revealed that patients who lost the miR-10b have significant longer survival rates than those showing overexpression of this miRNA (p = 0.0018, Figure 3(a) and (b)).

The normalized levels of miR-10b in each tumor and non-tumor samples: miR-10b is significantly upregulated in tumor samples compared to the adjacent non-tumor tissues.

Correlation of miR-10b normalized expression with (a) tumor size, (b) differentiation, (c) distant metastasis, and (d) vascular invasion.

(a) MiR-10b expression levels in dead patients compared to the surviving ones and (b) Kaplan–Meier survival curves showing the correlation of miR-10b expression with overall survival.
MiR-10b CpG methylation analysis and TWIST-1 expression in colorectal tissues
To explore the effects of the methylation profile on miR-10b expression in tumor tissues, we have performed MSP. The methylated allele of miR-10b was detected in 46% of cases (23/50). A representative example of the MSP results is shown in Figure 4(a). The aberrant CpG DNA methylation strongly correlates with the loss of expression of the miR-10b (p = 0.02, Figure 4(b)). In control tissues, seven out of the 10 tested samples showed unmethylated profile. To confirm the role of this CpG DNA methylation on the loss of miR-10b expression, we analyzed its expression in the HT-29 cell line. Cells were treated with 5-Aza, then the methylation pattern and the expression level of miR-10b were compared between the treated and untreated cells. In untreated cells, miR-10b promoter was fully methylated with a loss of expression (p = 0.035, Figure 4(c)); however, the addition of the 5-Aza restores the unmethylated profile, and, consequently miR-10b expression. In fact, in treated cells, miR-10b was upregulated by a mean factor of 261,984 (standard error (SE) range is 93,925–1 382,239), in comparison with untreated ones (p < 0.0001).
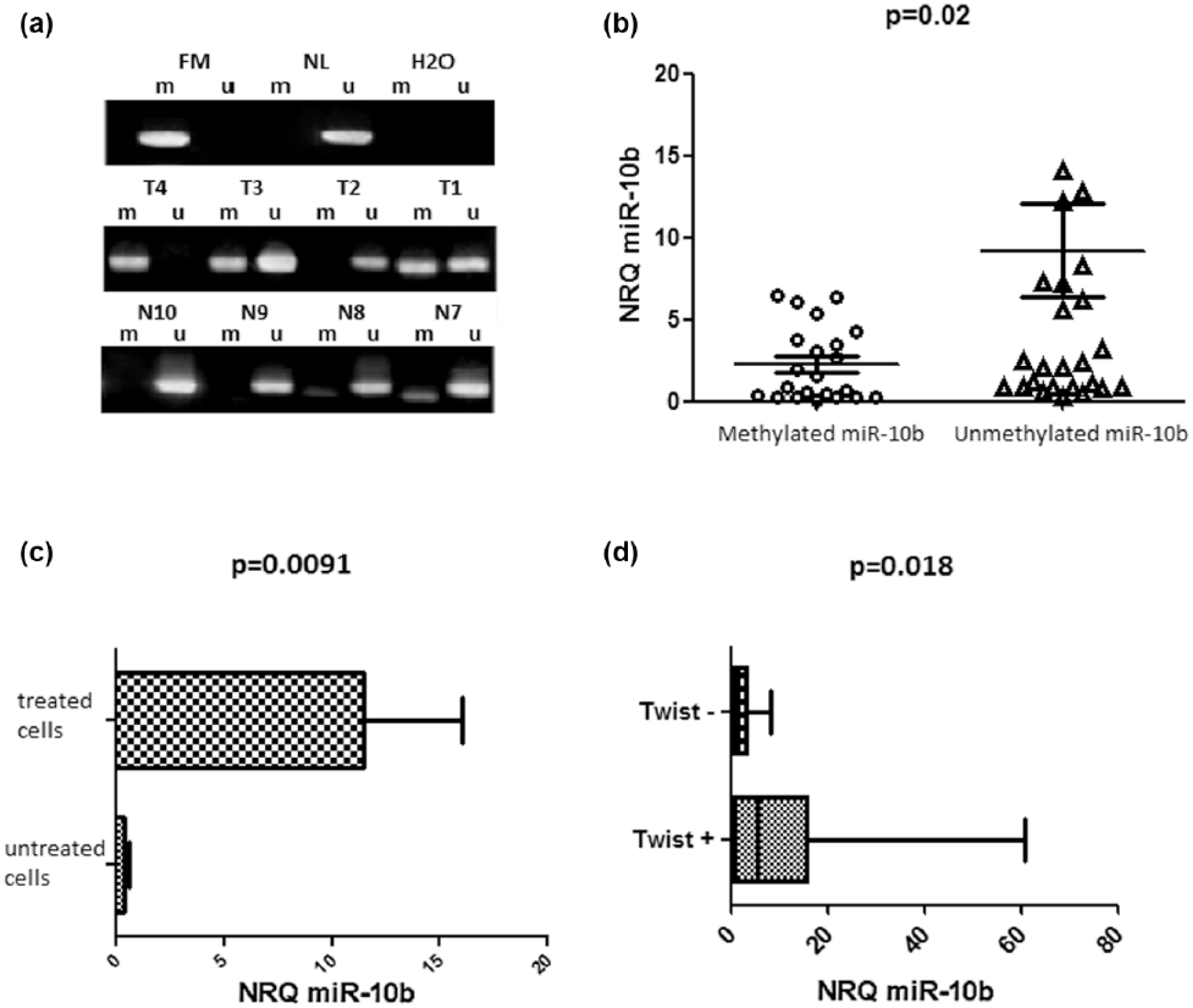
MiR-10b regulation in CRC samples: (a) representative results of MSP of miR-10b methylation status in CRC cases. H2O: negative control for MSP. T1–T4: tumor DNAs; N7–N10: normal mucosa DNAs; FM: fully methylated DNA; NL: normal lymphocytes DNA, (b) expression of miR-10b in sporadic colon cancer tumors with methylated profile in comparison to unmethylated samples, (c) difference in miR-10b expression between treated and 5-Aza treated HT-29 cell lines, and (d) correlation of miR-10b downregulation with TWIST-1 loss of expression.
Concerning the regulation of miR-10b by TWIST-1, we noticed a loss of TWIST-1 mRNA, associated with miR-10b downregulation (p = 0.018, Figure 4(d)).
Expression of HOXD10 in CRC
In attempt to validate the effect of miR-10b expression on its downstream target, we have performed RT-PCR analysis on the available sporadic tumors specimens. We found that 70% of CRC samples are positive for HOXD10 mRNA, which strongly correlated with the loss of miR-10b (p < 0.0001, Figure 5(a)). Associations of HOXD10 expression with different clinicopathological features have shown positive correlation with early TNM stages (I and II) (p = 0.044), absence of vascular invasion (p = 0.018) and well differentiated tumors (p = 0.002, Figure 5(b)).

HOXD10 expression in colorectal cancer tumor samples: (a) miR-10b normalized expression compared to HOXD10 mRNA presence and (b) significant correlation between HOXD10 expression and clinicopathological features. Red line refers to tumors differentiation, green line to vascular invasion, and blue line to TNM stage. r: Pearson correlation coefficient; p: significance.
Loss of miR-10b leads to E-cadherin expression in CRC
In our cohort, the E-cadherin expression was observed in 58.82% of samples (Figure 6(a) and (b)). Furthermore, we have demonstrated that increased levels of miR-10b was accompanied by downregulation of E-cadherin protein in tumor tissues (p = 0.0008, Figure 6(c)).

E-cadherin immunohistochemical staining and expression of miR-10b: (a) E-cadherin immune-expression in tumor tissues showing strong membranous accumulation in samples with miR-10b loss of expression, (b) E-cadherin immune-expression in normal adjacent mucosa showing membranous localization, and (c) miR-10b normalized expression compared to E-cadherin protein membranous expression.
Discussion
MiRNAs have been implicated in regulating diverse cellular pathways, and they can function as oncogenes or tumor suppressors. 17 MiR-10b has been described as an oncogenic miRNA that is overexpressed in different types of cancer and involved in tumor invasion and metastasis.17–23 In this study, increased levels of miR-10b have been detected in tumor samples compared to non-tumorous adjacent mucosa. Recently, Wang et al. 28 have reported that greater concentrations of miR-10b have been detected in CRC tissues compared with adjacent non-tumor tissues and normal tissues from healthy controls. The expression levels of miR-10b increased in line with aggressive behavior such as size of tumors >5 cm (p = 0.023), poor differentiated tumors (p = 0.016), metastasis (p = 0.0022), and vascular invasion (p = 0.01). Our data are concordant with previous studies showing that miR-10b promotes invasion in CRC and is overexpressed in patients at an advanced stage of the disease.27,28
MiRNA silencing by promoter hypermethylation may represent a mechanism by which cancer develops and progresses. To better understand the mechanism responsible for the overexpression of miR-10b in CRC, we investigated DNA CpG methylation of the miR-10b promoter. We have demonstrated that the miR-10b promoter is methylated in 46% of tumors suggesting that this epigenetic change is involved at least in part in the regulation of miR-10b in CRC. This DNA methylation has correlated significantly to the loss of miR-10b expression (p = 0.02). The CpG DNA methylation of miR-10b has been investigated in gastric cancer and renal carcinoma and high methylation rates, associated with miR-10b downregulation, have been observed.24,25,39 Moreover, in HT26 cell lines, we confirm that DNA methylation is responsible on regulating the miR-10b expression since the treatment with 5-Aza restores the expression of miR-10b. Besides the CpG DNA methylation involved in regulating the expression levels of miR-10b in cancer cells, Ma et al. 17 have identified TWIST-1 as transcription factor which binds directly to an E-box sequence proximal to its putative promoter and therefore regulates the miR-10b expression. In this study, we have found out that TWIST-1 mRNA has been lost in about 70% of samples, but it was positively correlated with the miR-10b loss of expression (p = 0.0018), which is in agreement with previous reports. 40
As one of the target genes of miR-10b, HOXD10 is considered as the main effector that negatively regulates tumor metastasis. 16 In our series, the HOXD10 mRNA has been detected in 35 out of 50 CRC cases (70%) and correlated with TNM stage (p = 0.043), vascular invasion (p = 0.018), and tumor differentiation (p = 0.002). In addition, increased levels of miR-10b has been detected in CRC samples negative for HOXD10 mRNA supporting that this miRNA regulates negatively HOXD10 and inhibits its translation. Those results are consistent with previous findings, suggesting that miR-10b could generate tumor metastasis,23–25 as the present study found that increased miR-10b also contributed to metastasis in CRC. On the contrary, and among key proteins that are involved in migration and invasion of tumor cells, the E-cadherin is a major player. In fact, using metastatic breast cancer cells, Liu et al. 30 have demonstrated that miR-10b regulates E-cadherin expression and that silencing of miR-10b restores E-cadherin expression and therefore suppresses the potential for tumor invasion/metastasis.
In addition, the regulation of E-cadherin by the miR-10b has been reported in non-small cells lung cancer (NSCLC). The authors have shown that the E-cadherin mRNA and protein were overexpressed in miR-10b-suppressed cells compared with controls, suggesting that miR-10b might be necessary for driving the expression of this protein in NSCLC. 31
In our study, the expression of E-cadherin had inversely correlated with the expression levels of miR-10b in CRC. In fact, miR-10b expression levels have markedly reduced in tumors displaying E-cadherin immunostaining (p = 0.0008).
In conclusion, our study has shown high levels of miR-10b detected in tumor tissues compared to the adjacent normal mucosa, suggesting its role as onco-miR in CRC. The high expression of miR-10b was associated with aggressive clinical behavior such as metastasis, vascular invasion and tumor differentiation. These results indicate that miR-10b may play a role in invasion in CRC which partially passed through E-cadherin. These data support the involvement of miR-10b in CRC and suggest that it might be a potential candidate for novel therapeutic targets in the treatment of metastatic CRC.
Footnotes
Acknowledgements
The authors thank Professor A. Abdelmaksoud for editing the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by a grant from the Tunisian Ministry of Higher Education and Scientific Research.