
Abstract
Lung cancer, especially the non-small-cell lung cancer, is a highly aggressive vascular cancer with excessively activated signaling pathways. Tumor-associated calcium signal transducer 2, also known as trop2, was identified to be correlated with tumor proliferation and invasion of non-small-cell lung cancer; however, the biological role of trop2 in neovascularization of non-small-cell lung cancer remained elusive. In this study, we first verified that trop2 was overexpressed in non-small-cell lung cancer tissues as well as cell lines and that the increased expression of trop2 promoted non-small-cell lung cancer cell proliferation and invasion. Then, we expanded the biological role of trop2 by in vitro and in vivo angiogenesis assay. The tubular formation analysis revealed that trop2 promoted non-small-cell lung cancer angiogenesis in vitro, and the immunohistochemistry staining of vascular markers (CD31 and CD34) provided evidences that trop2 promoted in vivo neovascularization. The results of polymerase chain reaction array revealed that trop2 promoted the expression level of two well-known angiogenesis factors MMP13 and PECAM1. By screening the trop2-related signaling pathways, we observed that excessive angiogenesis was correlated with activation of ERK1/2 signaling pathway, and ERK1/2 inhibitor (U0126) could suppress the tubular formation ability induced by trop2 expression. These results suggested that trop2 facilitated neovascularization of non-small-cell lung cancer via activating ERK1/2 signaling pathway. Targeting trop2 might provide novel anti-angiogenesis strategy for non-small-cell lung cancer treatment.
Introduction
Lung cancer, predominantly non-small-cell lung cancer (NSCLC), remains the biggest cancer-related killer worldwide with 221,200 estimated new cases and 158,040 estimated deaths in 2015. 1 Surgical resection was considered as the best possible treatment for NSCLC patients; however, only 20%–25% patients were suitable for potentially curative resection. 2 Despite the fact that chemotherapy options had been proven with promising results for the broader population in recent years, the survival status of NSCLC patients remained worrying with an overall 5-year survival less than 20%, raising an urgent need to seek more effective strategies for NSCLC treatment. 3 The past decade witnessed a revolutionary drug development based on “druggable” protein kinases including epidermal growth factor receptor (EGFR) and anaplastic lymphoma kinase (ALK).4,5 Furthermore, recent studies reported that targeting human epidermal growth factor receptor 2 (HER2), phosphatase and tensin homolog deleted on chromosome 10 (PTEN), and nuclear factor-kappa B (NF-κB) had also shown potential to promote the treatment of NSCLC in the future.6–8 Thus, a better understanding of the molecular regulatory mechanisms might contribute to improving the current therapeutics and prognosis of NSCLC patients.
The carcinogenesis of lung cancer is a multistage process involving genetic and epigenetic alterations of cancer-related genes as well as the abnormal modifications of corresponding proteins. 9 Recently, tumor-associated calcium signal transducer 2 (TACSTD2), also known as trop2, was considered to be correlated with NSCLC progression. Pak et al. 10 reported that trop2 was overexpressed in lung adenocarcinoma and squamous cell carcinoma tissues compared with the normal bronchial epithelium and pneumocyte. In addition, several studies also revealed the biological roles of trop2 in NSCLC. Trop2 overexpression promoted proliferation and invasion of lung adenocarcinoma cells, while silencing trop2 suppressed the biological behaviors. 11 These results provided oncogenic evidences of trop2 in NSCLC progression. However, the research of Wang et al. 12 revealed that loss of trop2 expression promoted the tumorigeneis, as well as the epithelial–mesenchymal transition of lung squamous cells, suggesting the tumor suppressive functions of trop2 in NSCLC. In this study, we confirmed the clinical significance of trop2 expression in NSCLC tissues, and clarified the role of trop2 in cell proliferation and invasion. Importantly, we expanded the molecular regulatory mechanisms involved in NSCLC neovascularization.
Materials and methods
Clinical specimens
The clinical research protocol was approved by the Ethical Committee of Henan Provincial People’s Hospital with written consent from all patients. A total of 107 clinical samples were collected from NSCLC patients in the Department of Thoracic Surgery of Henan Provincial People’s Hospital by routine thoracic surgery (I, II, and IIIA stage patients) or non-surgical procedures including fiberoptic bronchoscopy (IIIB and IV stage patients). Fresh tissues were stored in liquid nitrogen soon or fixed with formaldehyde for immunohistochemistry (IHC) staining. Patients who had undergone any chemo- or radiotherapy were excluded for this study.
Cell culture and lentiviral transduction
Human lung cancer cell lines A549, NCI-H520, NCI-H441, NCI-H226, and normal immortalized bronchial epithelial cell line HBE were purchased from American Type Culture Collection (Manassas, VA, USA). Cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) at 37°C in a 5% CO2 incubator. The medium was supplemented with 10% fetal bovine serum (FBS), 100 µg/mL streptomycin, and 100 µg/mL penicillin. For the stable silencing of trop2, A549 cells and NCI-H520 cells were transfected with lentivirus that expressed trop2 short hairpin RNA (shRNA; GeneChem, Shanghai, China) or the control lentivirus.
Quantitative real-time PCR
Quantitative real-time polymerase chain reaction (qRT-PCR) was conducted as previously described. 13 In brief, total RNA from clinical specimens and cell lines was extracted with Trizol reagent (Invitrogen, USA) according to the manufacturer’s instruction. The SYBR Green PCR Kit (Applied Biosystems, USA) was used for real-time polymerase chain reaction (PCR) analysis. GAPDH was used as an internal loading control for gene expression analysis. The primers used for qRT-PCR assay were trop2: 5′-GCCTACTACTTCGAGAGGGACA-3′ (sense) and 5′-CAGTTCCTTGATCTCCACCTTC-3′ (antisense); GAPDH: 5′-TTGGTATCGTGGAAGGACTCA-3′ (sense) and5′-TGTCATCATATTTGGCAGGTTT-3′ (antisense).
Western blot
Proteins were extracted with radioimmunoprecipitation assay (RIPA) buffer (Solarbio, China) with protease inhibitor and phosphatase inhibitor (Sigma, USA). Proteins from lysed cells were fractionated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. Then, the membranes were blocked with 5% bovine serum albumin (BSA) in TBST (tris-buffered saline with tween 20) and incubated with the specific antibody overnight at 4°C. The membranes were then washed with TBST three times and incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies. Proteins were visualized using an ECL Kit (Pierce, USA). Antibodies against GAPDH (5174), caspase 3 (9662), p44/42 MAPK (ERK1/2) (4695), p-ERK1/2 (4094), AKT (9272), p-AKT (4060), EGFR (4267), and p-EGFR (2244) were purchased from Cell Signaling Technology (USA). Antibodies against trop2 (ab65006), PECAM1 (ab28364), and MMP13 (ab75606) were purchased from Abcam (USA).
Transient transfection
pcDNA3.1-trop2 and the control plasmids were purchased from GeneChem. Lipofectamine™ 2000 (Invitrogen) was used for transient transfection according to the instruction. Briefly, 3 × 105 cells were seeded into six-well plates, and plasmids were transfected into cells at 70% confluence. RNA isolation, protein extraction, and cell function assays were conducted 48 h after transfection.
Cell proliferation, migration, and invasion assay
Cell Counting Kit-8 (CCK-8; Dojindo, Japan) was used for evaluating the cell viability. In brief, cells were seeded in 96-well plates, 200 µL (1000 cells) per well, and placed in the incubator for 5 days. The viability of cells was determined at 0, 24, 48, 72, and 96 h. Viable cells were quantified at each 24-h interval with CCK-8 according to the manufacturer’s protocol, and the absorbance at 450 nm was measured by the microplate reader (Epoch; BioTek, USA). Cell migration and invasion ability were evaluated according to a previously published protocol of our study. 13
In vitro angiogenesis assay
Tubular formation ability of human umbilical vein endothelial cells (HUVECs) was employed for evaluating the angiogenesis ability in vitro. A total of 3 × 104 cells were resuspended with 200-µL supernatant of corresponding tumor cells (A549/ctrl-shRNA, A549/trop2-shRNA, H520/pcDNA3.1, and H520/pcDNA3.1-trop2 with or without ERK1/2 inhibitor) and plated into 96-well plates coated with 50-µL Matrigel (BD Biosciences, USA) in advance. After 8-h incubation, the tube formation ability was evaluated by counting the tubular number, the tubular length, and intersections according to the published protocols.
Tumor angiogenesis PCR array analysis
The tumor angiogenesis PCR array, consisting of 17 genes known to be involved in angiogenesis, was used for detecting the molecular changes of A549 and H520 cells. In brief, total RNA of A549/ctrl-shRNA, A549/trop2-shRNA, H520/pcDNA3.1, and H520/pcDNA3.1-trop2 was isolated for qRT-PCR assay.
Nude mice study
All animal experiments were performed according to the guidelines for scientific use of live animals and were approved by the local ethical committee. BALB/c nude mice (4 weeks old) were housed in a specific-pathogen-free (SPF) environment and cared according to the institutional guidelines for animal care. A total of 4 × 106 cells in 100 µL of phosphate-buffered saline (PBS) were injected subcutaneously into the flanks of nude mice, and the tumor volume was evaluated every 5 days according to the following formula: V = tumor length × tumor width2 / 2. After 30 days, the subcutaneous tumors were resected, weighed, and fixed in 10% neutral buffered formalin for IHC staining. IHC staining was performed as described previously. 13 The antibodies used in this study were CD31 (ab28364; Abcam) and CD34 (ab81289; Abcam, USA).
Statistics
Student’s t test was used for analyzing quantitative data between two groups. One-way analysis of variance (ANOVA) was employed for analyzing data of more than two groups. A p value less than 0.05 was considered to be significant. All the analyses were performed using the SPSS software (version 11.0).
Results
Trop2 was overexpressed in lung cancer and correlated with tumor stage
To explore whether trop2 was associated with NSCLC progression, we examined the expression levels of trop2 in 107 paired NSCLC tissues by qRT-PCR. The results indicated that trop2 expression was significantly up-regulated in tumor tissues compared with non-tumor tissues, which was in accordance with the result of a recent study (Figure 1(a)). We next explored the relationship between trop2 level and clinicopathological parameters of NSCLC patients. The clinical and pathologic parameters of 107 NSCLC patients were presented in Table 1. Based on the cutoff value at 1.5-fold up-regulation, the samples were divided into two groups: trop2 high-expression group (n = 58) and trop2 low-expression group (n = 49). We observed that the level of trop2 was significantly higher in advanced-stage NSCLC (p = 0.004). However, we got no difference between trop2 level with gender, age, differentiation, histology type, and smoking status (Table 1). In addition, Figure 1(b) listed a more detailed relationship between trop2 level and NSCLC stage, showing that the level of trop2 increased as the stage became advanced, with stage IV NSCLCs being the highest and stage I NSCLCs the lowest (p < 0.01).

Trop2 was correlated with NSCLC progression and promoted cell proliferation and invasion. (a) expression of trop2 was analyzed in NSCLC tissues and non-tumor tissues by qRT-PCR assay. (b) the expression level of trop2 was increased as the tumor stage increased. (c, d) trop2 was overexpressed in a couple of NSCLC cell lines compared with the normal immortalized bronchial epithelial cell line HBE by qRT-PCR and Western blot analysis. (e) the silencing efficiency of trop2 in A549 cells was verified by Western blot assay. (f) silencing trop2 in A549 cells suppressed cell proliferation. (g) silencing trop2 in A549 cells suppressed cell invasion ability. (h) the level of trop2 in NCI-H520 cells was evaluated by Western blot assay after transfected with pcDNA3.1-trop2 or control plasmid. (i) overexpression of trop2 in NCI-H520 cells promoted the cell proliferation. (j) overexpression of trop2 in NCI-H520 cells promoted the cell invasion ability.
Correlative analysis of trop2 levels with clinicopathological parameters of NSCLC patients.

We then detected the expression of trop2 in a series of human NSCLC cell lines (namely, A549, NCI-H520, NCI-H441, NCI-H226, and HBE) by RT-PCR and Western blot. We observed that trop2 was up-regulated in NSCLC cell lines A549, NCI-H520, NCI-H441, and NCI-H226 compared with the normal immortalized bronchial epithelial cell line HBE (Figure 1(c) and (d)).
Trop2 promoted cell proliferation and invasion ability in vitro
We first wanted to confirm the biological roles of trop2 in NSCLC cell proliferation and invasion, which had been evaluated in a couple of recent studies. A549 cells were transfected with trop2 shRNA or stable negative control and selected with puromycin at a concentration of 5 µg/mL. Silencing efficiency of trop2 at protein level was examined by Western blot (Figure 1(e)). As presented in Figure 1(f) and (g), the proliferation and invasion ability of A549 cells were significantly repressed after trop2 down-regulation. Then we wanted to know whether overexpression of trop2 could promote these aggressive behaviors. NCI-H520 cells, which had a relatively lower trop2 expression level, were transiently transfected with pcDNA3.0-trop2 or the control plasmids. The protein level was examined at 48 h after transfection (Figure 1(h)). As shown in Figure 1(i) and (j), overexpression of trop2 promoted the proliferation and invasion ability of H520 cells, which coincided with the results of A549 cells.
Trop2 promoted in vitro and in vivo angiogenesis ability of NSCLC
Tumor growth and metastasis need glorious angiogenesis to provide enough nutrition. Thus, we first evaluated the role of trop2 on in vitro angiogenesis. As the result, silencing trop2 in A549 cells significantly suppressed the tubular formation ability of HUVECs by counting the tubular number, tubular length, and intersections (Figure 2(a)), while the overexpression of trop2 promoted the tubular formation ability (Figure 2(b)). Then, we wanted to know whether trop2 could regulate in vivo neovascularization of NSCLC. A549/trop2-shRNA and A549/ctrl-shRNA cells were subcutaneously injected into the right flank of nude mice (five mice per group). The tumor volume of each mouse was examined every 5 days. As shown in Figure 2(c), silencing trop2 in A549 cells inhibited the tumor growth by examining the tumor volume as well as the tumor weight. Then, the subcutaneous tumors were evaluated with the proliferation indicator ki67 and two common indicators of angiogenesis: platelet endothelial cell adhesion molecule-1 (CD31) and hematopoietic progenitor cell antigen (CD34). We observed that ki67 intensity and both angiogenesis marker (CD31 and CD34) expression were decreased in the A549/trop2-shRNA group compared with the A549/ctrl-shRNA group (Figure 2(d)). These results suggested that silencing trop2 inhibited in vitro tubular formation as well as in vivo neovascularization of NSCLC.
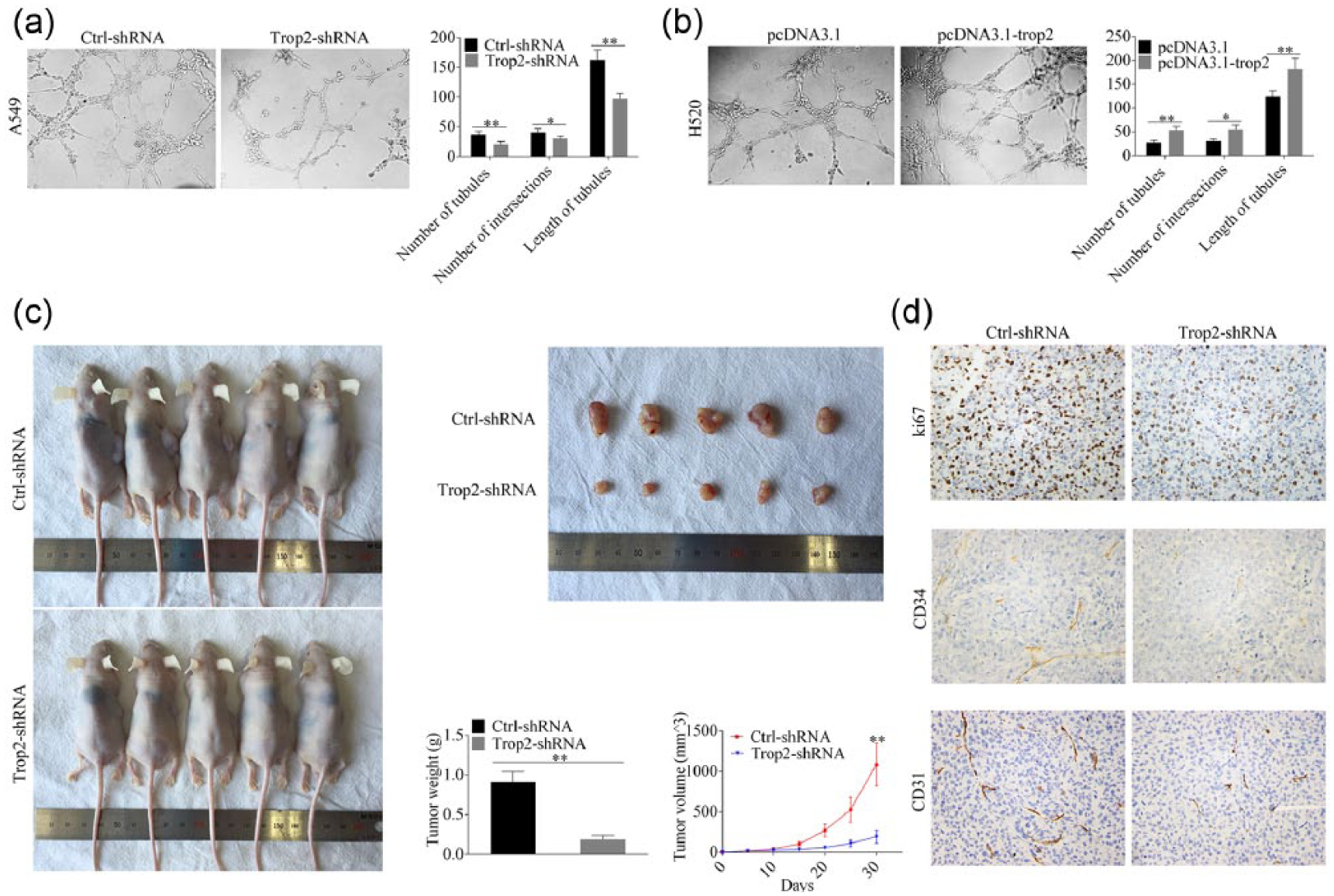
Trop2 promoted in vitro and in vivo angiogenesis ability of NSCLC. (a) tubular formation ability of HUVECs resuspended with supernatant from A549/ctrl-shRNA or A549/trop2-shRNA was assessed and number of tubules, length of tubules, and number of intersections were analyzed. (b) rich tubules were observed in the NCI-H520/pcDNA3.1-trop2 group compared with the control group. (c) A549/ctrl-shRNA and A549/trop2-shRNA cells were injected into the right axillary fossa of nude mice. The tumor volume was evaluated every 5 days and A549/trop2-shRNA cells formed smaller xenografts compared with A549/ctrl-shRNA cells. (d) ki67 intensity and both angiogenesis markers (CD31 and CD34) expression were decreased in the A549/trop2-shRNA group compared with the A549/ctrl-shRNA group.
Trop2 promoted self-renewal ability of HUVECs
Self-renewal ability, including the proliferation ability, apoptosis rate, and migration ability, determined the tubular formation ability of HUVECs. To this end, we then examined the proliferation ability of HUVECs resuspended with supernatant of each group. We observed that proliferation ability of HUVECs was decreased in the trop2-silenced group and increased in the trop2-overexpressing group compared with the corresponding control group (Figure 3(a)). In addition, the level of cleaved-caspase 3, the most important end-cleaving protease of cell apoptosis, was increased after trop2 down-regulation and decreased after trop2 overexpression (Figure 3(b) and (c)). More importantly, the migration ability of HUVECs was suppressed in the trop2-silenced group (Figure 3(d) and (e)) and enhanced in the trop2-overexpressing group (Figure 3(f) and (g)), which provided more direct evidences that trop2 could regulate neovascularization of NSCLC.

Effects of trop2 on self-renewal ability of HUVECs. The tumor supernatant from A549/ctrl-shRNA, A549/trop2-shRNA, NCI-H520/pcDNA3.1, and NCI-H520/pcDNA3.1-trop2 cells was collected to treat HUVECs, and the proliferation, migration, and apoptosis of HUVECs were examined. (a) The proliferation of HUVECs was decreased in the A549/trop2-shRNA group compared with the A549/ctrl-shRNA group, while the proliferation of HUVECs was increased in the NCI-H520/pcDNA3.1-trop2 group compared with the NCI-H520/pcDNA3.1 group. (b, c) the level of cleaved-caspase 3, the most important end-cleaving protease of cell apoptosis, was increased after trop2 down-regulation in A549 cells and decreased after trop2 overexpression in NCI-H520 cells. (d, e) the migration of HUVECs was decreased in the A549/trop2-shRNA group compared with the A549/ctrl-shRNA group. (f, g) the migration of HUVECs was increased in the NCI-H520/pcDNA3.1-trop2 group compared with the NCI-H520/pcDNA3.1 group.
Trop2 promoted expression level of multiple angiogenesis factors
To understand the molecular mechanisms of trop2-mediated angiogenesis of NSCLC cells, we conducted a PCR array containing 17 genes known to be involved in angiogenesis in A549/ctrl-shRNA, A549/trop2-shRNA, H520/pcDNA3.1, and H520/pcDNA3.1-trop2 cells. Results showed that the messenger RNA (mRNA) levels of MMP13 and PECAM1 were down-regulated after silencing trop2 and up-regulated after trop2 overexpression (Figure 4(a)). The protein levels of MMP13 and PECAM1 were in accordance with the mRNA level change (Figure 4(b)).

(a) Trop2 promoted the expression level of multiple angiogenesis factors. Tumor angiogenesis PCR array was utilized to profile A549/ctrl-shRNA, A549/trop2-shRNA, H520/pcDNA3.1, and H520/pcDNA3.1-trop2 cells. (b) expression level of MMP13 and PECAM1, which showed significant mRNA level change in PCR array, was confirmed by Western blot assay. (c) Western blot analysis for AKT, p-AKT, EGFR, p-EGFR, and β-catenin in indicated cell lysates.
Trop2 regulated neovascularization of NSCLC via activating ERK1/2 signaling pathway
Recent study reported that trop2 could regulate ERK, AKT, EGFR, and β-catenin signaling.14–17 Thus, we screened these signaling pathways by evaluating the protein level of ERK1/2, p-ERK1/2, AKT, p-AKT, EGFR, p-EGFR, and β-catenin. As shown in Figure 4(c), we just observed a slight down-regulation of p-EGFR after silencing trop2, while the expression level of ERK1/2, p-ERK1/2, AKT, p-AKT, and EGFR showed no significant change. Interestingly, however, we observed that the level of phospho-ERK1/2 was decreased in the A549/trop2-shRNA group and increased in the NCI-H520/pcDNA3.1-trop2 group compared with the control group of each cell line (Figure 5(a)). Figure 5(b) showed that the expression level of phospho-ERK1/2 was increased in a couple of NSCLC cell lines and was in accordance with the trop2 level shown in Figure 1(d). We next investigated whether trop2 could regulate tubular formation of NSCLC cells via the ERK1/2 pathway with an ERK1/2 inhibitor (U0126) at different concentrations. Figure 5(c) showed that 1-µM U0126 significantly inhibited the level of phospho-ERK1/2, and 10-µM U0126 had a more efficient inhibitory effect of NCI-H520/pcDNA3.1-trop2 cells. At the same time, inhibition of ERK1/2 signaling pathway in NCI-H520/pcDNA3.1-trop2 cells suppressed the angiogenesis by evaluating the tubular formation ability (Figure 5(d)–(g)). Therefore, trop2 regulated neovascularization of NSCLC via ERK1/2 pathway.

Trop2 regulated neovascularization of NSCLC via activating ERK1/2 signaling pathway. (a) silencing trop2 decreased the level of phospho-p44/42 MAPK, while the expression of trop2 promoted the level of phospho-p44/42 MAPK. (b) the level of phospho-p44/42 MAPK was examined in a couple of NSCLC cell lines. (c–e) after treatment with U0126 at indicated concentrations, the level of phospho-p44/42 MAPK was detected and the corresponding tubular formation ability of each group was evaluated.
Discussion
Trop2, which contained a large extracellular domain, a single transmembrane domain, and a short intracellular tail, was first discovered as a biomarker of invasive trophoblast cells.18,19 Recent studies had reported that trop2 was involved in the progression of multiple human tumors. In colorectal cancer, trop2 was correlated with poor prognosis and biological aggressiveness.20,21 As to cervical cancer, trop2 promoted cell proliferation and invasion via regulating ERK signaling pathway.22,23 The oncogenic role of trop2 was also reported in laryngeal carcinoma, pancreatic cancer, and gallbladder cancer.24–26 As to lung cancer, the biological studies were mainly focused on cell proliferation and invasion.11,19 However, the angiogenic role of trop2 was only reported in human glioma. 27 Considering the fact that tumor growth needed continuous supplement of nutrients via the blood vessel, we hypothesized that trop2 might be involved in angiogenesis of NSCLC.
Clinical data of recent studies reported a controversial relationship between trop2 expression and NSCLC progression. Trop2 overexpression was positively correlated with clinicopathological parameters, while it was negatively correlated with the survival state of NSCLC patients. 10 In this work, we first confirmed the clinical significance of trop2 expression in NSCLC tissues. We observed that trop2 was overexpressed in NSCLC tissues and correlated with late TNM stages. In addition, trop2 was also overexpressed in a couple of NSCLC cell lines compared with HBE, which was derived from the normal bronchial epithelium. As to the biological study, we observed that silencing trop2 in A549 cells suppressed the proliferation and invasion ability, while the expression of trop2 in NCI-H520 cells promoted these aggressive behaviors. These results confirmed the conclusion of recent studies and suggested the potential oncogenic role of trop2 in NSCLC progression.
Great efforts had been made to explore the mechanisms of tumor angiogenesis in the past decade; nonetheless, the critical mechanisms that truly contributed to NSCLC angiogenesis remained elusive. A recent study reported that trop2 expression was positively correlated with microvessel density in glioma specimens. 27 Given that trop2 could regulate proliferation and invasion of NSCLC cells, we hypothesized that trop2 might be involved in tumor angiogenesis of NSCLC. The in vitro tubular formation study of HUVECs and in vivo IHC staining of pivotal angiogenesis-related markers revealed that trop2 could truly regulate tumor angiogenesis in NSCLC. As an attempt to understand the events through which trop2 promoted tubular formation ability, we assessed the self-renewal ability of HUVECs cells by proliferation, apoptosis, and migration analysis. We observed that trop2 promoted the proliferation and migration ability, while it inhibited the apoptosis of HUVECs. These data strongly support the hypothesis that trop2 was a critical angiogenesis regulator of NSCLC.
Trop2 could activate several pathways that contribute to its tumor-promoting properties. Stoyanova et al. 17 reported that trop2 promoted the stem-like properties and self-renewal ability of epithelial cancers via β-catenin signaling. In another study, Cubas et al. 14 demonstrated that trop2 contributed to tumor pathogenesis by activating the ERK MAPK pathway. However, the signaling pathways activated by trop2 in NSCLC were rarely reported. In this study, we observed that the expression of phospho-p44/42 MAPK was obviously reduced after trop2 down-regulation, while the forced expression of trop2 promoted the level of phospho-p44/42 MAPK. In addition, the level of trop2 and phospho-p44/42 MAPK showed a fairly high degree of consistency in a couple of NSCLC cell lines. These results indicated that the expression of trop2 activated the MAPK signaling pathway, while the relationship between trop2-MAPK signaling and NSCLC neovascularization still needed to be verified. Then, we identified that U0126, an ERK1/2 inhibitor, reversed the pro-angiogenic effects induced by trop2 expression, suggesting involvement of ERK MAPK pathway in trop2-mediated angiogenesis of NSCLC cells. In conclusion, our data unraveled that trop2 promoted angiogenesis of NSCLC via ERK1/2 signaling pathway. Trop2 might serve as an anti-angiogenesis target of NSCLC.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.