
Abstract
Despite the consolidated clinico-pathological correlates of Caveolin 1 expression in non–small cell lung cancer, the available data on the role of Caveolin 1 in relation to proliferation, migration, and metastasis in lung adenocarcinoma cells is still scant. Here, we aimed to confirm whether Caveolin 1 may act as a promoter of cell growth in human lung adenocarcinoma using in vitro and in vivo models, supported by a survival analysis of Caveolin 1 expression in a series of 116 primary lung adenocarcinomas. The silencing of endogenous Caveolin 1 expression in H522 lung adenocarcinoma cells through stable shRNA transfection significantly inhibited cellular proliferation in vitro and in vivo, in a lung adenocarcinoma xenograft mouse model. The bioluminescence imaging analysis revealed that tumors derived from Caveolin 1 shRNA-transfected cells grew slower than control xenografts. However, this difference progressively diminished over time and was definitively lost after 21 days. This was consistent with a progressive Caveolin 1 re-expression, which started at day 7. The association between the restored expression of Caveolin 1 and the restart of tumor growth in vivo supports the booster role of Caveolin 1 in lung adenocarcinoma progression. To further confirm this role, Caveolin 1 expression was assessed by immunohistochemistry in a series of 116 human lung adenocarcinomas. Positive Caveolin 1 tumors accounted for 20% of cases and were associated with a significantly worse overall survival compared to Caveolin 1-negative cancers. Taken together, these data highlight that Caveolin 1 expression confers a proliferative advantage in lung adenocarcinoma cells, thus fostering increased tumor aggressiveness.
Introduction
Caveolin 1 (Cav1) is a member of the caveolin family of scaffold proteins that regulate the activity of many signaling molecules inside caveolae. Cav1 is involved in various normal cell functions, including lipid transportation, cell growth, and death regulation. Nevertheless, the role of Cav1 in cancer development and tumor progression has been widely documented with a bulk of data supporting cellular transformation, tumor growth, cell death and survival, multidrug resistance, angiogenesis, cell migration, and metastasis. 1 Of note, in various cancer models, Cav1 plays a biphasic role, as during the early stages of cancer progression it is frequently down-regulated, acting as a tumor-suppressor, whereas it becomes upregulated during more advanced cancer stages acquiring an oncogenic function, which contributes to an aggressive and metastatic phenotype. 2 Furthermore, Cav1 expression in cancer cells is tissue-type dependent, being down-regulated in ovarian and colon carcinomas, as well as in mesenchymal sarcomas, but upregulated in bladder, esophageal, pancreatic, and breast cancers. 3 In lung cancer, we have previously shown a differential Cav1 expression across distinct histological types of non–small cell lung cancer (NSCLC) and an increased expression in brain metastases, which is significantly associated with poor prognosis4,5 and radioresistance. 5
Despite the consolidated clinico-pathological correlates of Cav1 expression in NSCLCs, 6 data on the role of Cav1 in relation to proliferation, 7 migration, and metastasis8,9 in NSCLC cells are scant.
In this study, we aimed to confirm whether Cav1 acts as a promoter of cell growth in human lung adenocarcinoma using in vitro and in vivo models, supported by a survival analysis of Cav1 expression in a series of 116 primary lung adenocarcinomas.
Materials and methods
Cell line and culture conditions
The human lung cancer cell line H522 (ATCC® CRL-5810™) was obtained from the American Type Culture Collection (Rockville, MD, USA). Cells were grown in RPMI-1640 medium (PAA Laboratories GmbH, Pasching, Austria) supplemented with 10% fetal bovine serum (FBS; PAA Laboratories GmbH), 1% glutamine (Sigma–Aldrich, St. Louis, MO, USA), and 1% streptomycin, penicillin, and fungizone (Sigma–Aldrich) at 37°C in a humidified 5% CO2 atmosphere.
Western blot analysis
Cell samples were lysed at 4°C for 30 min in radioimmunoprecipitation (RIPA) lysis buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% Triton X-100, and 0.1% sodium dodecyl sulfate (SDS)) added with protease inhibitors. The bicinchoninic acid (BCA) protein assay (Thermo Fisher Scientific Inc., Rockford, IL, USA) was used to quantify the total protein extracts. Proteins (25 g) extracted from H522 non-treated (NT) cells, control shRNA, and Cav1 shRNA-transfected cells were added with 5× loading buffer, boiled for 5 min and loaded onto an 8% polyacrylamide gel. Electrophoresis was performed in tris-glycine SDS (TGS) buffer. Total proteins were separated and then transferred to a nitrocellulose membrane and stained with Ponceau red. After saturation with TBS and 5% milk, the membrane was immunoblotted overnight at 4°C either with anti-Cav1 antibody (rabbit polyclonal, Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluted 1:1000 or with anti-β-actin (Sigma–Aldrich) at 1:2000 as a loading control. Both antibodies were diluted in tris-buffered saline (TBS) with 5% bovine serum albumin (BSA). Following washes in TBS Tween, the membrane was incubated with an anti-rabbit horseradish peroxidase (HRP)-labeled secondary antibody (diluted 1:5000, Bio-Rad, Hercules, CA, USA). The reaction was developed with ECL luminol (Bio-Rad).
Cav1 stable transfection
The H522 cells were transfected with commercially available Cav1 shRNA plasmid (h) (sc-29241; Santa Cruz Biotechnology) and control shRNA (Santa Cruz Biotechnology), following the manufacturer’s protocol. Briefly, 1 day before transfection, 2 × 105 cells per well in six-well plates were seeded into RPMI-1640 medium without antibiotics and then incubated until 50%–70% confluence for optimal transfection. After the cells had been rinsed with shRNA Transfection Medium (sc-108062; Santa Cruz Biotechnology), they were transfected with shRNA Transfection Reagent (sc-108061; Santa Cruz Biotechnology) for 7 h, after which 2× normal growth medium was added (normal growth medium containing two times the normal serum and antibiotics concentration). After the 48 h post-transfection, the medium was removed and replaced with fresh medium containing puromycin antibiotic (sc-108071; Santa Cruz Biotechnology). Efficiency of protein silencing was determined by immunohistochemistry and western blot analysis for all conditions: (1) NT cells, (2) control shRNA-transfected cells, and (3) Cav1 shRNA-transfected cells.
In vitro cell proliferation assay
The Cell Proliferation ELISA BrdU colorimetric assay (Roche, Penzberg, Germany) was used to assess cell proliferation at 0, 48, and 72 h in control shRNA versus Cav1 shRNA-transfected cells, according to manufacturer’s instructions.
Luciferase cell transfection
Luciferase was cloned in the plasmid vector pCCLsin.PPT.hPGK.GFPpre at XhoI site. Lentiviruses for cell transduction were obtained as previously described. 10 Cells were analyzed for the efficiency of transduction by enhanced green fluorescent protein (EGFP) content on a FACSCalibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA). When the efficiency of infection was below 80%, cells were sorted on a MoFlo High-Performance Cell Sorter (DakoCytomation, Glostrup, Denmark) to normalize both the intensity of fluorescence and the percentages of transduced cells.
Mice and bioluminescent imaging
Animal studies were conducted in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals. The protocol was approved by the Committee on the Bioethics of the University of Turin. Mice were kept in our institutional animal facility under well-controlled conditions of temperature and humidity with access to food and water ad libitum.
SCID mice (Charles River Laboratories, Lyon, France) were injected subcutaneously with H522 cell line (5 × 105 cells) expressing luciferase. Two xenografts for mice were performed: on the left were Cav1 shRNA-transfected cells and on the right were control shRNA-transfected cells. Tumor growth was followed for 4 weeks by bioluminescence using the IVIS system 2000 series (Xenogen Corp., Alameda, CA, USA). For the bioluminescence imaging, mice were anesthetized by isoflurane inhalation and were subsequently intra-peritoneally injected with 200 mL of 15 mg/mL
Human tumor cohort
A series of 116 primary lung adenocarcinomas were retrieved from the archives of the University of Turin Pathology divisions at Città della Salute e della Scienza (Molinette) Hospital and at San Luigi Hospital of Orbassano. The clinico-pathological features of this series have been previously reported.4,5 All patients had a metastatic disease (Stage IV) involving brain (71 cases), bone (13 cases), lung (16 cases), adrenal (4 cases), and liver (2 cases). They were all treatment naive at the time of diagnosis. The study received ethical approval from the local Institutional Review Boards. Immunohistochemistry was performed as described below.
Cells and tissue processing and immunohistochemistry
Cells were grown on sterilized coverslips (22 mm × 22 mm) in Petri dishes and washed in phosphate-buffered saline (PBS 1×) for 5 min and fixed in 4% neutral-buffered formalin for 10 min.
Xenograft and tissue samples were fixed in 4% buffered formaldehyde and paraffin-embedded. Three-micrometer-thick sections from the mouse tissue samples as well as from representative tumor blocks of the human adenocarcinoma samples were collected on Superfrost Plus slides for immunohistochemistry.
Cav1 immunohistochemistry was performed using an automated slide-processing platform (Ventana BenchMark XT AutoStainer, Ventana Medical Systems, Tucson, AZ, USA) and a polyclonal antibody (rabbit anti-Cav1, diluted 1:350; Santa Cruz Biotechnology). Cav1 staining was assessed as a categorical variable (negative or positive if present in at least 10% of neoplastic cells) and analyzed independently by three of the authors (R.S., E.D., and P.C.). Vascular endothelium represented an internal positive control. In discrepant cases, slides were reviewed at a multiheaded microscope to reach agreement.
Statistical analysis
Kruskal–Wallis rank-sum test was used to analyze differences between various conditions. Kaplan–Meier curve and log-rank test were used to assess differences in survival among patient subgroups. A p < 0.05 was considered as statistically significant. Analyses were performed using the 2.12.1 version of the R statistical package (www.r-project.org).
Results
Cav1 down-regulation inhibits H522 cells proliferation
High Cav1 levels were observed in H522 cells by both western blot and immunohistochemistry, which showed a moderate to strong membranous and/or cytoplasmic expression (Figure 1(a)). To estimate the effect of Cav1 on tumor cell proliferation, Cav1 endogenous expression was abrogated through a Cav1 shRNA transfection. The successful knockdown of Cav1 was confirmed using immunohistochemistry (Figure 1(b)) and western blot analysis (Figure 1(c)), both showing Cav1 down-regulation only in Cav1 shRNA-transfected cells.

In vitro studies: stable knockdown of Cav1 in lung adenocarcinoma cells. Cav1 H522 cell expression (a) before and (b) after Cav1 stable transfection, by immunohistochemistry; (c) western blot analysis confirmed Cav1 down-regulation in Cav1 shRNA-transfected cells (shCav1), whose protein expression levels were lower compared with the control group (shCTRL) and not transfected cells (NT). Equal protein loading was determined by probing with antibody to β-actin. (d) Cav1 down-regulated cells (shCav1) showed a significant decrease in cell proliferation when compared to control group (shCTRL), by Cell Proliferation ELISA BrdU colorimetric assay.
A significant decrease in cell proliferation was observed in Cav1 down-regulated cells when compared to the control group (Kruskal–Wallis rank-sum test, time 48 h: p = 0.02; time 72 h: p = 0.05) (Figure 1(d)).
Cav1 down-regulation inhibits in vivo tumor growth
Tumors which derived from control shRNA cells showed an initial faster growth compared with those from Cav1 shRNA-transfected cells, as shown by the higher bioluminescent signal, which was significant at day 3 (p = 0.02, T-test) and still higher, although not significant, at day 7. This difference was then lost at day 13 (Figure 2). During mouse grafting, a fraction of control shRNA and Cav1 shRNA H522-transfected cells was also separately cultured to investigate Cav1 expression by immunohistochemistry in vitro, in parallel with bioluminescence imaging analysis in vivo. Cav1 expression progressively increased in Cav1 shRNA H522-transfected cells after 13 and 21 days. Tumors from both control shRNA and Cav1 shRNA-transfected cells showed an infiltrative growth pattern and were mainly composed of large cells exhibiting a high degree of cellular and nuclear pleomorphism with high mitotic counts (>20 mitoses/10HPF). In addition, in both control shRNA and Cav1 shRNA cell-derived tumors, Cav1 expression displayed a heterogeneous distribution with a predominant membranous pattern, thus confirming in vivo the re-expression progressively observed in vitro starting from day 13 after silencing. Thereafter, at 21 days, no morphological and immunohistochemical differences were observed between the two groups (Figure 2).
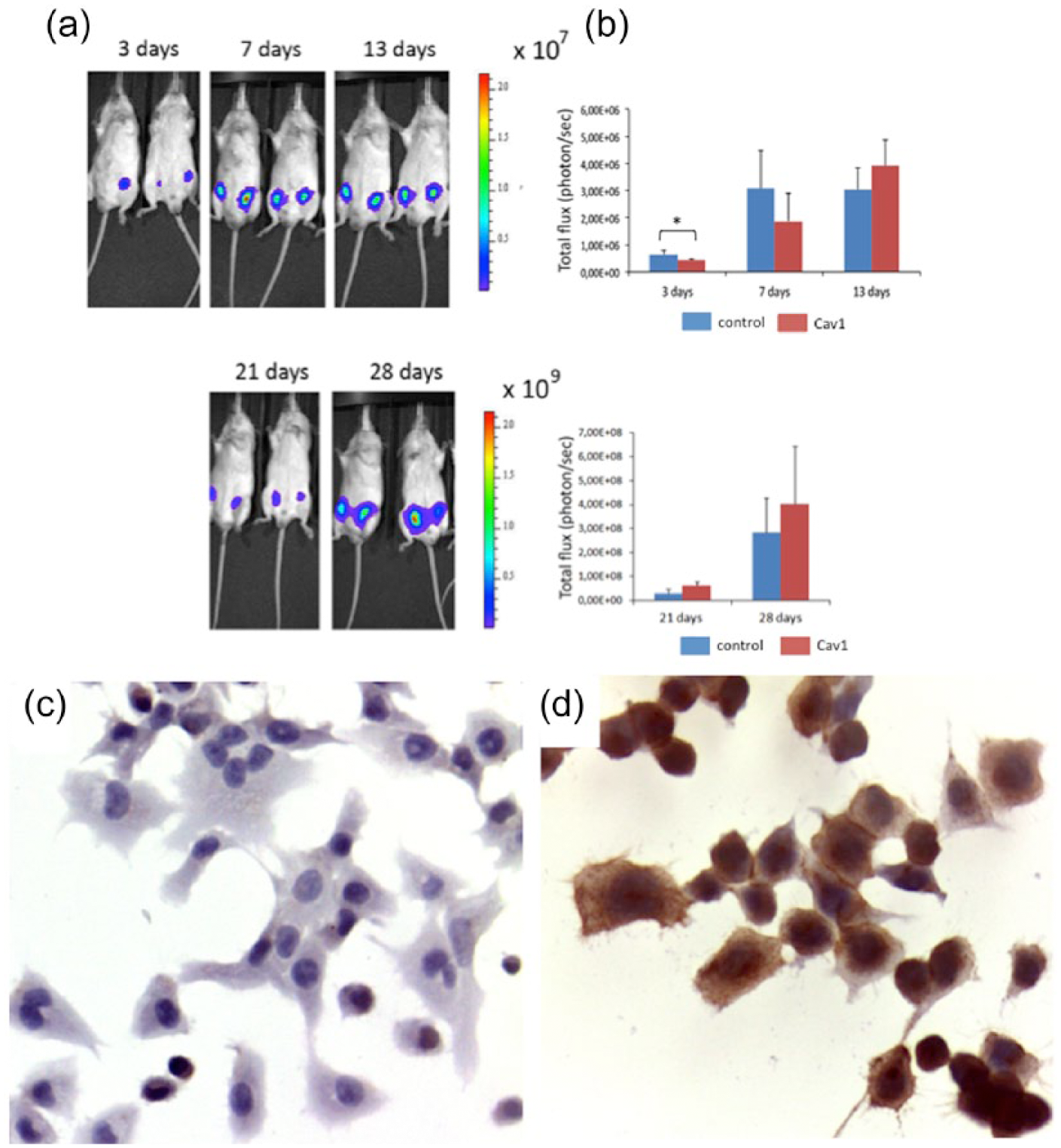
In vivo studies. Tumor growth evaluated at different time points by bioluminescence. (a) Representative images of mice analyzed by the IVIS system 3, 7, 13, 21, and 28 days after cell injection. Two xenografts for mice were performed at each stage: in each mouse, on the left side were Cav1 shRNA-transfected cells and on the right side were control shRNA-transfected cells. (b) Quantification of the bioluminescent signal expressed as total flux (photons/s) ± SD of four mice treated with control shRNA and Cav1 shRNA-transfected cells. (c and d) Cav1 immunohistochemistry in Cav1 shRNA cell-derived mice tumors at (c) 7 and (d) 21 days.
Cav1 expression is associated with a worse prognosis in lung adenocarcinomas
Cav1 expression was observed in a subset of lung adenocarcinomas accounting for 20% of the cohort (23/116). These tumors showed a mild to strong membrane or cytoplasmic staining of neoplastic cells (Figure 3). Univariate analysis for overall survival revealed that patients with Cav1-expressing tumors had a substantially shorter survival (mean, 20 months) than Cav1 negative (mean, 28 months) (log-rank test, p = 0.009).

Representative example of a Cav1-positive lung adenocarcinoma (a, hematoxylin and eosin, original magnification 400×) with strong membrane-cytoplasmic immunoreactivity for Cav1 (b, original magnification 400×).
Discussion
In this study, we explored the potential role of Cav1 in lung adenocarcinoma investigating lung cancer cells in vitro and in vivo and evaluated its prognostic impact in a cohort of lung cancer patients. We show that the silencing of endogenous Cav1 expression in H522 cells through shRNA transfection can significantly inhibit cellular proliferation in vitro. As a matter of fact, cell proliferation was reduced by Cav1 knockdown, whereas it was promoted in cells treated with control shRNA. As a matter of fact, Cav1 knockdown reduced cell proliferation, which was promoted in cells treated with control shRNA. This finding is consistent with other reports in which Cav1 silencing reduced the proliferation of cell lines from metastatic lesions of lung adenocarcinoma 11 and from breast cancer cells. 12 Meanwhile, forced Cav1 overexpression increased lung cancer cell proliferation in response to epidermal growth factor. 9
The in vivo evidence derived from shRNA-silenced H522 cells engrafted in mice (showing a significantly slower growth compared to wild-type cells) further corroborates our in vitro data. Interestingly, the difference between wild-type and Cav1-silenced grafts progressively diminished in parallel with a resumed expression of Cav1 in shRNA-silenced H522 cells, as shown by immunohistochemistry on cells cultured at day 7 following transfection. This tardive re-expression of Cav1 from silenced lung adenocarcinoma cells could be due to either the prevailing of Cav1 non-transfected clones or the progressive loss of silencing by transfected cells. To the best of our knowledge, this is the first evidence of a Cav1 expression–driven proliferative switch in in vivo grafts of lung adenocarcinoma, and the observed association between restored expression of Cav1 and restart of tumor growth in vivo may suggest a role of Cav1 in contributing to lung adenocarcinoma progression.
When tested in a series of lung adenocarcinomas, a 20% prevalence of Cav1-expressing tumors was identified, in line with previous reports.4,6,9 In addition, patients with Cav1-positive adenocarcinomas exhibited a significantly worse overall survival than Cav1-negative tumors. These data are consistent with a recent meta-analysis of NSCLC patients 6 showing that, when overexpressed, Cav1 is significantly associated with tumor progression thus conferring a higher risk of death and a reduced progression-free survival to NSCLC patients. Of note, we also recently provided evidence supporting Cav1 expression in brain metastases from lung adenocarcinoma as an independent predictor of worse outcome and radioresistance. 5
Taken together, our data validate the role of Cav1 expression in promoting lung adenocarcinoma cell proliferation, as shown by in vitro and in vivo experiments; furthermore, Cav1 may serve as a negative prognosticator in lung cancer patients, given the significant association with poor outcome. Further studies investigating the role of Cav1 in relation to other clinico-pathological features, including also the mutational status, are warranted as they may help provide a better stratification of lung adenocarcinomas.
Footnotes
Acknowledgements
E.D. and R.S. contributed equally to this manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by grants from “Fondi di Ricerca Locale ex-60%” to C.M. and P.C. and from “Dipartimento Rete Oncologica Piemonte e Valle d’Aosta” to P.C.