
Abstract
SIRT1 plays an important role in human malignant progression, inducing cancer cell proliferation and metastasis by regulating downstream gene expressions. However, little is known about the underlying mechanisms in which SIRT1 promotes pancreatic cancer tumorigenesis. The aim of this study is to investigate the SIRT1 expression levels and biological functions in promoting pancreatic cancer progression. We first investigated the expression of SIRT1 in a series of pancreatic cancer tissues as well as in a panel of pancreatic cancer cell lines. The effect of SIRT1 on cell activity was explored by knockdown experiments. Cell growth was measured using the MTT assay and colony-formation assay. Migration and invasion were tested using transwell assay. Our results showed that the expression of SIRT1 was significantly up-regulated both in pancreatic cancer tissues and cell lines. Knockdown of SIRT1 suppressed cell proliferation and migration of pancreatic cancer cells. This is the first report to disclose the role of SIRT1 in regulation of pancreatic cancer cell proliferation and migration, which may provide a potential therapeutic target for pancreatic cancer patients.
Pancreatic ductal adenocarcinoma, often called pancreatic cancer, is one of the most aggressive and lethal malignancies. 1 Despite new therapeutic approaches including surgery, radiotherapy, and chemotherapy for pancreatic cancer have improved, pancreatic cancer remains a disease with a dismal prognosis. 2 It is characterized by a high propensity for local tumor invasion and drug resistance, but the molecular events underlying this remain mysterious. 3 Thus, investigation of the molecular mechanisms underlying pancreatic cancer development and progression is urgently needed.
SIRT1, the human homolog of Sir2, is a member of sirtuins family. 4 SIRT1 is a NAD1-dependent class III deacetylase (HDAC) which plays a critical role in the regulation of critical biological processes such as metabolism, aging, oncogenesis, and cancer progression.5,6 Emerging evidence has suggested that SIRT1 is overexpressed in some cancers, and it contributes to cell growth, drug resistance, invasion, metastasis, and recurrence; 7 however, the expression and function of SIRT1 in pancreatic cancer progression is still unclear. In this study, we sought to determine the roles of SIRT1 in pancreatic cancer development and progression. We found a key role for SIRT1 as a tumor promoter that enhances invasive and metastatic potential in pancreatic cancer in vitro. Our findings provide a new insight into the use of sirtuin inhibitors for pancreatic cancer therapy.
Materials and methods
Human tissue specimens and immunohistochemical analysis
The tissue microarray (TMA) construction and immunohistochemical analysis were conducted. The TMA contained 108 patients with pancreatic cancer and matched tumor adjacent tissue. All samples were derived from patients who had not received adjuvant treatment including radiotherapy or chemotherapy prior to surgery. The TMA was prepared and processed for immunostaining using anti-SIRT1 (Abcam, Cambridge, MA, USA; ab32441). The staining results were scored by two investigators blinded to the clinical data. Besides, freshly frozen tissue samples and adjacent noncancerous controls were also available from 30 pancreatic cancer patients and snap-frozen in liquid nitrogen immediately after surgery. This study was approved by the legislation and ethical boards of First Affiliated Hospital, Henan University of Science and Technology. All subjects or their caregivers have written informed consent.
Cell lines
Human pancreatic ductal epithelial cell line HPDE, pancreatic cancer cells BxPC-3, CFPAC-1, CAPAN-1, SW1990, AsPC-1, PANC-1, and HS-766T were all obtained from Chinese Academy of Sciences Cell Bank (Shanghai, China). PANC-1 and HS-766T were grown in 5% CO2 saturated humidity, at 37°C, and cultured in Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 2 mmol/L glutamine and 10% fetal bovine serum (FBS) (both from Gibco, USA) and subcultured by harvesting with trypsin-EDTA. HPDE, BxPC-3, CAPAN-1, AsPC-1, CFPAC-1, and SW1990 cells were cultured in RPMI-1640 (Gibco) supplemented with 10% FBS.
RNA extraction and real-time quantitative polymerase chain reaction
Total RNA extraction, complementary DNA (cDNA) synthesis, and quantitative polymerase chain reaction (qPCR) were performed according to manufacturer’s standard procedure. Polymerase chain reaction (PCR) primers were as follows: for human glyceraldehyde 3-phosphate dehydrogenase (GAPDH), forward 5′-TGCCAAATATGATGACATCAAGAA-3′ and reverse 5′-GGAGTGGGTGTCGCTGTTG-3′; for STAT3 forward 5′-TGCTGGCCTAATAGAGTGGCA-3′and reverse 5′-CTCAGCGCCATGGAAAATGT-3′.
Western blotting assay
Cells were lysed in the cell lysates (Thermo) supplemented with protease inhibitors PMSF and Cocktail (Roche). Proteins were separated in 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose NC membranes (0.22 mm, Whatman). Membranes were blocked with blocking buffer (Li-COR), sequentially incubated in primary antibodies and secondary antibody. The primary antibodies included rabbit anti-human SIRT1 (Abcam), anti E-cadherin, anti-N-cadherin, anti-Vimentin (Santa Cruz Bio-technology, Santa Cruz, CA, USA), and anti-human GAPDH (Cell Signaling Technology, Inc., Danvers, MA, USA). The secondary antibody was Goat Anti-Rabbit IgG (Invitrogen). Protein levels were measured by gray value with Quantity One software.
Cell transfection
Cells were transduced with lentivirus vectors expressing SIRT1 ShRNA. These cells were cultured for 24 h, followed by the exposures to virus-containing supernatants (MOI520) via polybrene. Cells were selected by puromycin (2 mg/mL) (Sigma) 48 h later. ShRNA1-1 and ShRNA1-2 were the anti-SIRT1 ShRNA sequences. To turn off the effect of ShRNA, a scrambled sequence was used as a control. ShRNA sequences were as follows: ShSIRT1-1: GAAGTGCCTCAGATATTAA; ShSIRT1-2: GTTGACCTCCTCATTGTTA; ShScramble: GCGCGCTTTGTAGGATTCG.
Cell growth assay
Cell growth was determined by MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazoliumbromide; Sigma). Transfected cells were grown in complete medium. Each group was repeated in three wells for three times. After incubation for 72 h, MTT (5 mg/mL in phosphate-buffered saline, PBS) was added to each well. After 2 h of incubation with MTT, the plates were centrifuged at 500g for 5 min. After careful removal of the solution, 0.1 mL of dimethyl sulfoxide (DMSO) was added to each well and plates were shaken. The absorbance was recorded on a microplate reader at the wavelength of 540 nm.
In vitro invasion and migration assays
The invasive and migratory potential of cells were analyzed using Transwell chambers (8 µm pore; BD Biosciences, San Jose, CA USA). For invasion assays, cells in 100 µL of serum-free medium were added to the upper chamber coated with Matrigel. Medium supplemented with 10% FBS was added to the lower chamber. After incubation for 24 h, non-invading cells on the upper surface were removed with a cotton swab and cells invading to the lower chamber were fixed with methanol, stained with 0.1% crystal violet, dried, and photographed. For migration assays, cells in 100 µL serum-free medium were placed in the top chamber without Matrigel, and 500 µL of 10% FBS containing medium was added to the lower chamber. After 16 h, cells on the upper surface were removed and cells migrated to the lower chamber were fixed and stained as described above. The number of invading or migrating cells was counted under the microscope in five representative fields and expressed as the average per field.
Statistical analysis
SPSS version 19.0 (IBM SPSS, Chicago, IL) was used for statistical analysis in our study. The association of Sirt1 and clinicopathological variables was determined using chi-square test. p < 0.05 was considered statistically significant.
Results
SIRT1 expression was up-regulated in pancreatic cancer tissues
We first examined SIRT1 expression levels in clinical pancreatic cancer specimens. Immunohistochemical staining results showed that SIRT1 expression in pancreatic cancer specimens was significantly up-regulated compared with that in the adjacent non-tumoral tissue (Figure 1(a) and (b)). SIRT1 overexpression was observed in 82 of 108 (75.93%) pancreatic cancer specimens when compared with the adjacent non-tumoral tissue (40 of 108, 37.04%, p < 0.001). Then, we examined the SIRT1 messenger RNA (mRNA) levels in a series of pancreatic cancer tissues and matched adjacent non-tumor tissue using quantitative reverse transcription polymerase chain reaction (qRT-PCR). As shown in Figure 1(c), in comparison with the adjacent tissues, the pancreatic cancer tissues had a significantly increased SIRT1 mRNA expression, indicating that SIRT1 might play an oncogenic role in pancreatic cancer progression.

High expression of SIRT1 in pancreatic cancer and pancreatic cancer cells: (a) positive expression of SIRT1 in pancreatic cancer specimens (magnification, 200×), (b) negative expression of SIRT3 in corresponding nonneoplastic pancreatic tissues (magnification, 200×), (c) SIRT1 mRNA expression levels in pancreatic cancer and corresponding nonneoplastic pancreatic tissues, and (d) expression of SIRT1 in a panel of pancreatic cancer cell lines was detected by qRT-PCR. **p < 0.01.
SIRT1 promotes pancreatic cancer cell proliferation in vitro
To investigate the functional role of SIRT1 in pancreatic cancer progression, we performed qRT-PCR analysis to detect the expression of SIRT1 in a panel of pancreatic cancer cell lines. As shown in Figure 1(d), qRT-PCR results revealed that all pancreatic cancer cells expressed higher SIRT1 than human pancreatic ductal epithelial cell line HPDE. Among the pancreatic cancer cell lines, AsPC-1 and CFPAC-1 had relatively higher expression levels of SIRT1; thus, we used AsPC-1 and CFPAC-1 cells as a model to investigate the effect of SIRT1.
To further assess the biological function of SIRT1 in pancreatic cancer, we used two lentiviral shRNA vectors to specifically and stably knock down the expression of SIRT1 in AsPC-1 and CFPAC-1. A knockdown effect was observed by qRT-PCR and Western blot, and we found that SIRT1-sh1 achieved the greater efficacy in silencing SIRT1 expression compared to the negative control (Figure 2). We then analyzed the effect of SIRT1 on pancreatic cancer growth and metastasis in vitro. MTT assay was used to measure the proliferation of the cells and found that knockdown of SIRT1 expression significantly inhibited cell proliferation both in AsPC-1 and CFPAC-1 cells compared with the respective controls (Figure 3(a) and (b)), suggesting that SIRT1 may act as an oncogene involved in the promotion of pancreatic cancer.

(a) AsPC-1 and (b) CFPAC-1 cells were transfected with sh-SIRT1 or NC. SIRT1 expression level was verified by qRT-PCR; (c) and (d) SIRT1protein expression was analyzed by Western blotting; knockdown of SIRT1 reverses EMT in AsPC-1 and CFPAC-1 cells.
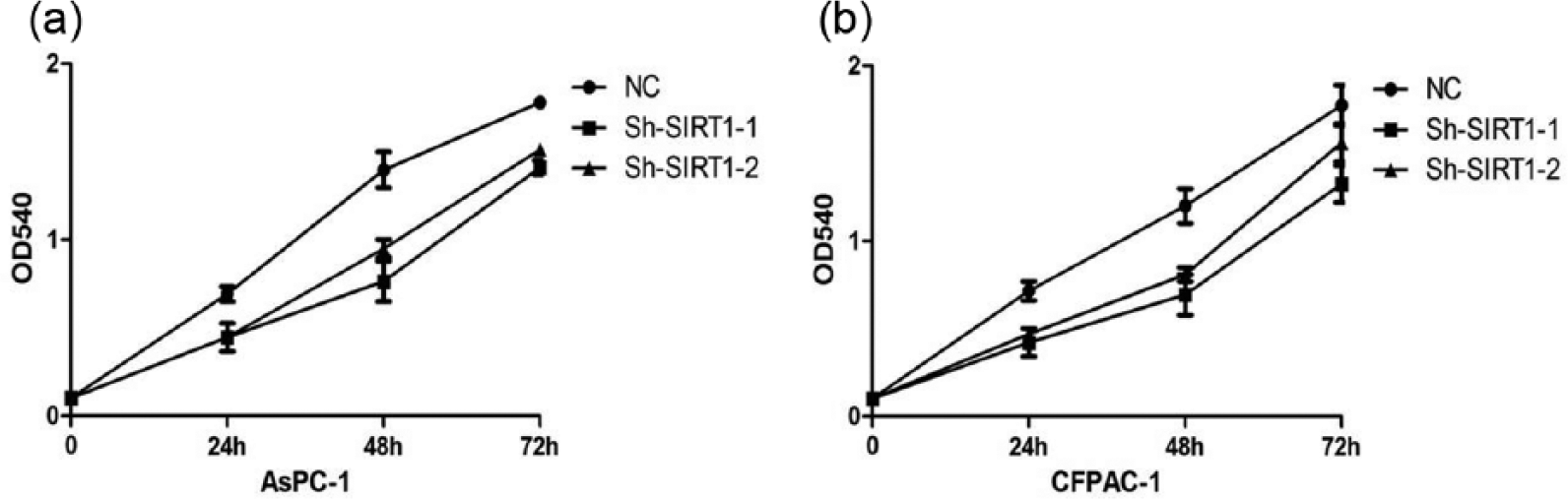
Cell proliferation was measured by MTT assay. The cell proliferation curves were based on the absorbance at 540 nm measured every 24 h. (a) Knockdown of SIRT1 inhibits the proliferation of AsPC-1 cells. (b) Knockdown of SIRT1 inhibits the proliferation of CFPAC-1 cells.
Effects of SIRT1 expression on pancreatic cancer cell migration and invasion
Then, we performed migration assays to detect the effect of SIRT1 on cell migration and invasion in AsPC-1 and CFPAC-1 cells. To assess the impact of SIRT1 expression on pancreatic cancer migration and invasion, the transwell assays were performed. The results showed that inhibition of SIRT1 significantly impaired migration and invasion capacity of AsPC-1 and CFPAC-1 cells in comparison with NC groups (Figure 4(a) and (b)). To investigate the underlying mechanisms by which SIRT1 promotes pancreatic cancer cell migration and invasion, Western blotting was performed to show expression of epithelial-mesenchymal transition (EMT)-relevant markers, and the results indicated that suppression of SIRT1 concomitantly reduced expression of vimentin and N-cadherin protein expression and restored E-cadherin expression compared to control (Figure 2).

(a) Effect of SIRT1 interference on the ability of migration and invasion of AsPC-1 cells according to Transwell assays. (b) Effect of SIRT1 interference on the ability of migration and invasion of CFPAC-1 cells according to Transwell assays.
Discussion
Pancreatic cancer is a devastating disease with a 5-year survival rate of less than 5%, and its incidence is increasing annually. 1 In the United States, pancreatic cancer is projected to become the second most common cause of cancer deaths by 2030. 2 Due to its characteristics, especially extensive local tumor invasion and drug resistance, the effect of traditional surgery or radiotherapy and chemotherapy is poor. It is urgent to explore the molecular mechanisms related to the pancreatic cancer progress to seek new treatment strategies.
SIRT1, the human homolog of Sir2, is a NAD+-dependent class III deacetylase (HDAC) which can deacetylate both histone and non-histone proteins. 8 SIRT1 was discovered to be associated with regulating multiple cellular and physiological processes, including diabetes, inflammation, neurodegenerative diseases, stress responses, cell survival, metabolism, aging, and longevity. 9 Due to its ability of promoting cell survival through regulating some nuclear proteins, such as p53, Bax, E2F1, and Ku70, SIRT1 is considered as an essential role in tumorigenesis and more and more researches focus on the role of Sirt1 in human cancers.10–12 Previous studies have demonstrated that SIRT1 is overexpressed in some cancers, including colon cancer, prostate cancer, skin cancer, and breast cancer.13,14 However, the function of SIRT1 in regulating pancreatic cancer progression and metastasis remains enigmatic.
In this study, we first demonstrated the SIRT1 overexpression in pancreatic cancer patients using a series of pancreatic cancer tissues and matched adjacent non-tumor tissues through qRT-PCR. Moreover, we observed that the patients with high SIRT1 expression were significantly associated with tumor grade. These suggested that SIRT1 overexpression may influence the progression of pancreatic cancer. Consistent with the result in patients, the expression of SIRT1 was significantly upregulated in various pancreatic cancer cell lines.
Then, we investigate the function of SIRT1 in pancreatic cancer progression in vitro. We downregulate the expression of SIRT1 in two pancreatic cancer cell lines via shRNA transfection and found that the proliferation of SIRT1 knockdown cancer cells was significantly inhibited. In addition, we also revealed the critical role of SIRT1 in migration and invasion, and inhibition of SIRT1 could weaken the pancreatic cancer cell migration and invasion, indicating that the SIRT1 may be a novel therapeutic approach against pancreatic cancer.
Although the evidence has highlighted the critical role of SIRT1 on pancreatic cancer, the molecular mechanism is still unclear. To explore the mechanisms of SIRT1 tumor progression, EMT, which has been demonstrated to be an important step in metastasis, was tested. EMT could dissolve the cell–cell junctions between non-motile cells and convert them into individual, motile mesenchymal cells by loss or downregulation of cell–cell adhesion molecules, such as E-cadherin, and many studies have suggested the role of SIRT1 in regulating EMT in various cancers.15,16 Our finding shows that knockdown SIRT1 expression could significantly reduce the vimentin expression and restored E-cadherin expression.
In conclusion, we showed that SIRT1 is overexpressed in pancreatic cancer and leads to increased cell proliferation and invasion. In addition, inhibition of SIRT1 could suppress pancreatic cancer progression in vitro, suggesting that SIRT1 may be utilized as a potential target molecule for human pancreatic cancer.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.