
Abstract
Background
Ischaemic heart disease (IHD) is currently managed by drugs and minimally invasive procedures. However, the necessity for safe, eco-friendly herbal-based medications is consistently advocated to prevent the side effects. Medicinal plants, rich in cardioprotective phytochemicals, offer promising potential for developing safer therapeutics to mitigate IHD progression while reducing treatment-related risks.
Purpose
Trans-ferulic acid (TFA), a phenolic secondary metabolite recognised for its various pharmaceutical applications, has been selected for in vitro assessment of its toxicity and cardioprotective efficacy.
Materials and Methods
H9c2 cells were exposed to ischaemic conditions and treated with varying concentrations of TFA (3.1–50 µg/mL). Cytotoxicity and cardioprotective effects were assessed using MTT and lactate dehydrogenase (LDH) assays. Collagen degradation was evaluated by Sirius Red staining, while quantitative real-time polymerase chain reaction (qRT-PCR) was used to analyse the expression of nuclear factor erythroid 2-related factor 2 (NRF2) and HIF1α genes involved in oxidative stress response and hypoxia regulation.
Results
50 µg/mL TFA showed significantly (p < .001) less toxicity and exhibited 86% viability in ischaemic H9c2 cells; the protective levels were significantly (p < .001) higher (36%) when compared to the untreated group. Decreased LDH activity in the treated group demonstrated the cardioprotective efficacy of TFA, with a value of 239 U/mL compared to the untreated cells (356 U/mL). TFA inhibited the collagen degradation induced by ischaemia, hence preventing the organisation of cardiac muscle tissue, as evidenced by Sirius Red staining. TFA treatment upregulates the NRF2 and suppresses the expression of HIF1α by means of mitigating cellular damage caused by oxidative stress and inflammation, which has a significant impact on the occurrence of IHD.
Conclusion
These findings suggest TFA as a promising phytochemical candidate for developing novel therapeutic strategies against IHD.
Introduction
Ischaemic heart disease (IHD) constitutes a global health challenge and is the leading cause of morbidity and mortality, with 8.99 million deaths recorded in 2021 (Naghavi et al., 2024). IHD encompasses a range of conditions from stable angina to acute myocardial infarction, with the most severe form being coronary artery obstruction, which results from restricted blood supply to the heart muscle. IHD pathophysiology is associated with an increased risk of thrombosis, atherosclerosis and myocardial ischaemia (Severino et al., 2020). Pharmacotherapies such as beta-blockers, anti-platelet agents, ACE inhibitors and statins are frequently used, depending on the health condition, to control cholesterol and blood pressure levels and prevent clot formation (Severino et al., 2020). Numerous dietary strategies have been recommended to reduce the prevalence of IHDs, gaining increased attention among the clinical and scientific groups. Functional foods and dietary supplements containing antioxidants and other metabolites are vital for neutralising reactive oxygen radicals and inhibiting cellular damage (Chan et al., 2010; Qin et al., 2014).
Phytochemicals such as flavonoids, carotenoids, polyphenols, terpenoids and alkaloids are being explored as possible alternative agents in the management of cardiovascular diseases, including myocardial ischaemia (Arshad et al., 2025). The anti-inflammatory, antioxidant, vascular-modulatory and cholesterol-lowering properties of phytochemicals offer therapeutic effects for the treatment of cardiovascular diseases (Asao & Asaduzzaman, 2018). For instance, tannins derived from Urtica simensis (burn nettle) had prevented the risk of myocardial injury in rats by diminishing the harmful effects of cardiac biomarkers (Tesfaye et al., 2021). The risk of myocardial ischaemia can be mitigated by reducing the levels of antioxidant enzymes and the activity of membrane-bound Na+/K+ ATPase. Phytochemicals derived from Musa paradisiaca enhance the pro-inflammatory cytokines, cardiac troponin-I and malondialdehyde, hence contributing to the improvement of cardiac health (Suleiman et al., 2021).
Trans-ferulic acid (TFA), a phenolic compound abundant in plants, exhibits anti-microbial, anti-cancer, anti-angiogenic, anti-ageing, anti-inflammatory and antioxidant properties (Rahmoune et al., 2017; Rezaeiroshan et al., 2020; Zhang & Feng, 2006). Studies have shown earlier that phenolic compounds have effective pharmacological benefits (Singh et al., 2025). The cardioprotective effect of ferulic acid (4-hydroxy-3-methoxycinnamic acid), which inhibits ferroptosis, has been investigated earlier by Liu et al. (2022). Ferulic acid mitigated oxidative stress and cellular damage induced by high glucose, suggesting its potential as a therapeutic intervention for IHD (Bekheit et al., 2025). Singh and Patil (2022) have demonstrated the possible anti-diabetic effect of TFA via the activation of adenosine monophosphate-activated protein kinase (AMPK) under high glucose conditions (Singh et al., 2022). TFA increased the reactive oxygen species (ROS) levels and induced apoptotic cell death of human lung cancer cells (H1299 cells). The ability to inhibit cancer cell migration and modulate phosphorylation promises potential anti-cancer therapy (Fong et al., 2016). The possible wound healing properties of ferulic acid combined with macromolecules, including collagen, chitosan-gelatin and glycerol phosphate hydrogel, have been previously demonstrated (Cheng et al., 2011; Kumar et al., 2020). Kaczmarek et al. (2020) proved that the mechanical properties of collagen are enhanced when incorporated with ferulic acid. Also, ferulic acid may serve as a potential inhibitor of collagen fibril production in fibrotic diseases. While research on the therapeutic effects of TFA for neurological disorders, diabetes and cancer is still ongoing, studies evaluating its cardioprotective properties are few or non-existent. While ferulic acid has been reported to exert cardioprotective effects, the transcriptional modulation of NRF2 and HIF1α by its stable derivative, TFA, under ischaemic conditions remains poorly defined. Hence, we designed a study to assess the therapeutic potential of TFA, a stable derivative of ferulic acid, in managing IHD through cytotoxicity analysis, lactate dehydrogenase (LDH) assay, Sirius Red staining, and its association with NRF2 and HIF1α gene regulation in an in vitro ischaemic cardiomyoblast model.
Materials and Methods
Cytotoxicity Assay
H9c2 cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cell culture was maintained in a humidified incubator with 5% CO2 at 37°C, nourished with Dulbecco’s Modified Eagle’s Medium (DMEM; Sigma-Aldrich, USA) supplemented with 10% foetal bovine serum (FBS),
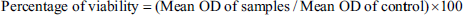
Where OD represents optical density.
Investigating the Cardioprotective Efficacy
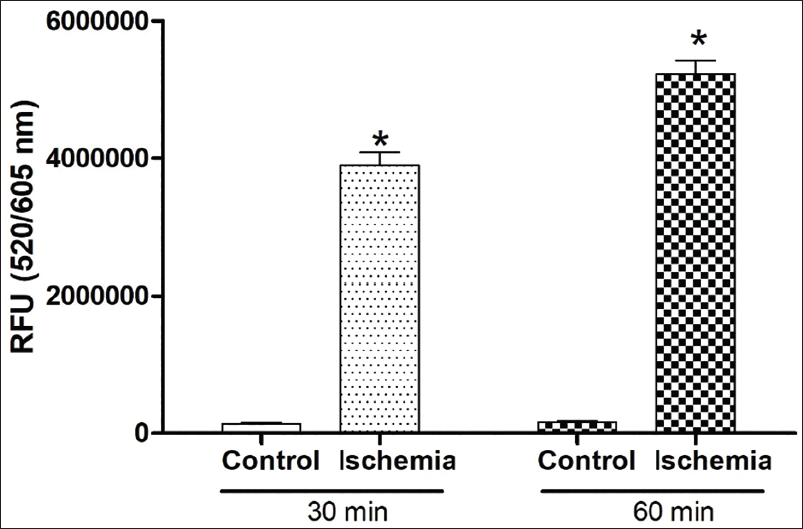
Two-day-old confluent monolayers of H9c2 cells were induced with ischaemia using the ischaemic buffer and incubated for 4 h in 5% CO2 incubator. The ischaemic buffer solution, adjusted to pH 6.6, was composed of 125 mM NaCl, 8 mM KCl, 1.2 mM KH2PO4, 1.25 mM MgSO4, 1.2 mM CaCl2, 6.25 mM NaHCO3, 20 mM 2-deoxyglucose, 20 mM sodium lactate and 20 mM HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid). The induction of ischaemia was validated using ROS generation measurement using a kit (Abcam; ab186027). Briefly, at the end of incubation, the medium was replaced with 100 µL of ROS Red Stain working solution, according to the manufacturer’s protocol. The cells were then incubated again for 30 min and 60 min, and fluorescence was measured at 520/605 nm using a SpectraMax microplate reader. The ROS values were scored as fluorescence intensity units (Figure 1).
In a separate set of experiments, following incubation, the ischaemic buffer was replaced with varying concentrations (3.1, 6.25, 12.5, 25 and 50 µg/mL) of DMEM diluted with TFA to evaluate the cardioprotective effectiveness. Cytotoxicity in the cardiac myoblast cells was assessed using the MTT assay as previously described. OD values and images were documented as described above to elucidate the cardioprotective efficacy of TFA.
Reactive Oxygen Species Assay
To measure and validate ischaemia induction, the reactive oxygen species (ROS) assay was performed using a cell-based quantitative assay using a kit (Abcam; ab186027). Briefly, H9c2 cells were plated into 96-well black plates with clear bottoms. The plate was incubated overnight for confluency, ischaemic buffer was added and incubated for 4 h in 5% CO2 incubator. Ischaemia-induced H9c2 cells were treated with 50 µg/mL of DMEM diluted TFA and incubated for 24 h. At the end of the incubation, the media was replaced with 100 µL ROS Red Stain working solution as described by the manufacturer’s protocol. The cells were then incubated again for 30 min, and fluorescence was measured at 520/605 nm using a SpectraMax microplate reader. The ROS values were scored as fluorescence intensity units.
Lactate Dehydrogenase Assay
Lactate dehydrogenase assay (LDH) is used as a quantitative marker enzyme for the intact cell, its activity providing information on cellular glycolytic capacity. The assessment of LDH release (leakage) is a critical and commonly used assay for severe irreversible cell damage. Ischaemia-induced H9c2 cells were treated with 50 µg/mL of DMEM diluted TFA and incubated for 24 h. Untreated ischaemia-induced cells were maintained as controls. The LDH activity was measured according to the Deutsche Gesellschaft für Klinische Chemie (DGKC) method. This standardised method is based on the catalytic reduction of pyruvate to lactate by LDH in the presence of nicotinamide adenine dinucleotide (NADH), during which NADH is oxidised to NAD+. Briefly, 50 µL of cell-free supernatant collected from the test wells was added with 1 mL working reagent (Bergmeyer, 2012). The decrease of OD was recorded at 340 nm in a spectrophotometer, after 5 min of incubation. The following formula can calculate the activity of LDH,
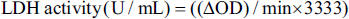
Sirius Red Staining
Sirius Red staining analysis is used to evaluate fibrosis by visualising the thin septa and collagen fibres. Sirius Red stain binds to the triple-helical structure (Gly–X–Y) found in collagen fibres, the property of which helps to assess collagen in cardiac cells under bright field. The Sirius Red Stain kit (Biognost, Croatia) was used to perform the assay (Zhu et al., 2022). H9c2 cells were cultured in a 12-well plate, and ischaemia-induced cells were treated with TFA as described in the above analysis. Control and untreated ischaemic cells were maintained to compare the efficacy of TFA on ischaemic myoblasts. The collagen degradation in the wells was determined by the Sirius Red staining method after 5 days. Supernatant media was replaced with 0.1% Sirius Red saturated in picric acid for 1 h. Excess or unreacted stain was removed by washing with 0.01 M HCl and dissolving it in 0.1 M NaOH. OD of the supernatant was read at 540 nm, and resin-mounted dehydrated slides were examined using a microscope.
Quantitative Real-time Polymerase Chain Reaction Analysis to Find Out Gene Expression
TFA (50 µg/mL)-treated ischaemic cardiac myoblasts and the control cells were subjected to real-time polymerase chain reaction (RT-PCR) analysis to determine the expression of NRF2 and HIF1α genes. Total RNA was isolated using the total RNA isolation kit according to the manufacturer’s instructions (Invitrogen, product code 10296010). The purity and the concentration of total RNA were determined. Template complementary DNA was synthesised using the iScriptc DNA Synthesis Kit (Bio-Rad, Cat# 1708891 master premix for first-strand cDNA synthesis). The thermal cycler (Eppendorf Mastercycler) was programmed to undergo cDNA synthesis. The following cycling conditions were employed: priming for 5 min at 25°C; cDNA synthesis for 20 min at 46°C; and RT inactivation for 1 min at 95°C.
qRT-PCR analysis was performed using SYBR Green Master Mix (G Biosciences, Product code 786-5062) using LightCycler 96 (Roche). The data were analysed for the triplicates according to the 2−∆∆Ct method using LightCycler 96 SW 1.1 software based on the manufacturer’s instructions. Three cycles of initial activation, each for 2 min at 95°C; 10 s of denaturation at 95°C; 1 min of annealing at 58°C; and 1 min of extension at 72°C were set for 40 cycles (Fong et al., 2016). The expression of NRF2, HIF1α and GAPDH was investigated; the primer details are summarised in Table 1. The fragments of amplified DNA formed bands in the ethidium bromide-stained 1.5% agarose gel at a specific area corresponding to the molecular weight of the tested genes. Images of bands in the stained gel were visualised using a gel documentation system (ChemiDoc Imaging System, Bio-Rad).
Forward and Reverse Primers Used to Determine the Gene Expression of NRF2, HIF1α and GAPDH, and the Required Annealing Temperatures.

Statistical Analysis
All experiments were performed in triplicate and repeated at least three times independently. Data are presented as mean ± standard error of the mean (SEM). Statistical analyses were conducted using GraphPad Prism software (version 6.0). Differences between multiple groups were evaluated using one-way analysis of variance (ANOVA), followed by Dunnett’s post hoc test for comparisons between treated groups and the corresponding control group. A p value of <.05 was considered statistically significant.
Results
Cytotoxicity Assay
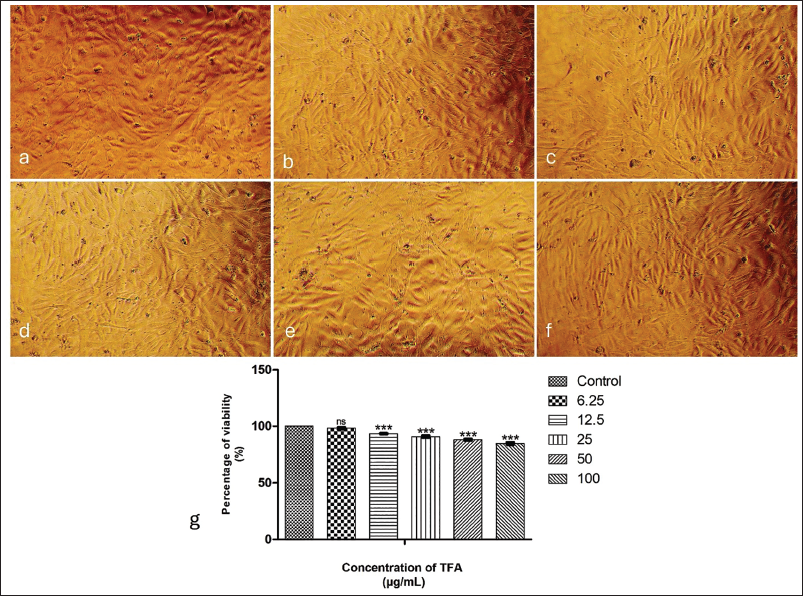
TFA demonstrated a dose-dependent increase in cytotoxicity with significant differences (p < .001) among each group, except between control and the lowest concentration (6.25 µg/mL). The images photographed using a phase contrast microscope illustrate the cytotoxicity levels recorded for different concentrations (Figure 2(a)–2(g)). The viability of healthy cardiac myoblasts exhibited changes when treated with varying concentrations of TFA. The percentages of viability ranging from 93% to 85% have been recorded for different concentrations of TFA: 12.5, 25, 50 and 100 µg/mL. The lack of significant change at 6.25 µg/mL indicates potential safety at lower concentrations. Higher doses of up to 100 µg/mL demonstrated a substantial reduction in cell viability; however, the viability levels remained at 85%, indicating the safety of using TFA for cardiac treatment (Figure 2(h)).
Cardioprotective Efficacy
Ischaemic H9c2 cells without treatment showed a significant decrease in viability, approximately 50%. Figure 3(a)–3(g) illustrates the cardioprotective efficacy of TFA in ischaemia-induced cardiomyoblasts. Treatment with TFA showed a dose-dependent increase (p < .001) in viability, ranging from 60% to 86% for the concentrations varying from 6.25 to 50 µg/mL. We observed a substantial increase (p < .01) of 54% in the viability of H9c2 cells, even at the lowest concentration of 3.1 µg/mL, compared to the untreated ischaemic group (50%) (Figure 3(h)).
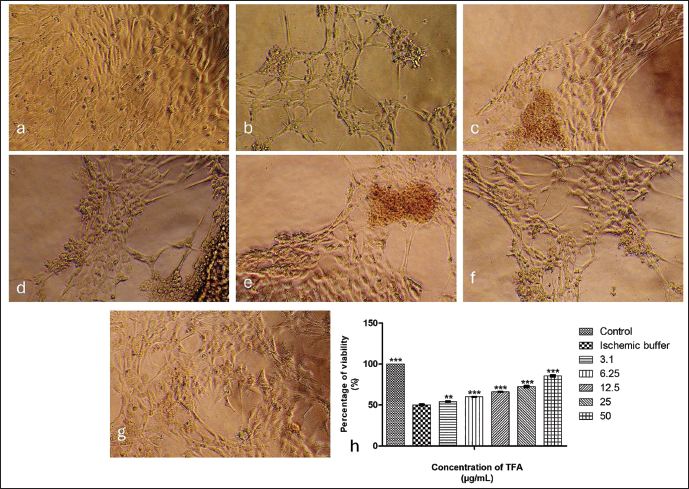
The concentration of 50 µg/mL TFA was selected for further mechanistic analyses because it demonstrated maximal cardioprotective efficacy under ischaemic conditions while maintaining minimal cytotoxicity in healthy H9c2 cells.
Lactate Dehydrogenase Assay
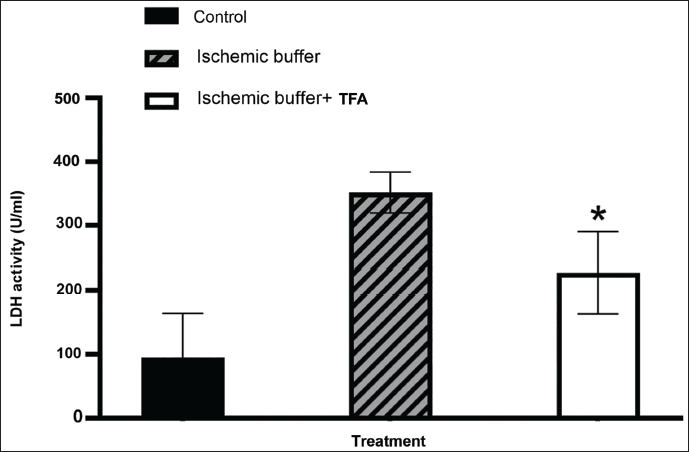
LDH assay examined the protective efficacy of TFA against ischaemia-induced cellular damage in H9c2 cells. The results demonstrate a significant reduction in LDH activity with TFA, suggesting its potential as a therapeutic agent. A considerable increase in LDH activity to 356 U/mL of activity in the ischaemia-induced H9c2 cells demonstrated cellular damage. 50 µg/mL TFA has significantly (p < .001) reduced the LDH levels to 239 U/mL, indicating the potential therapeutic efficacy against cellular damage (Figure 4). The LDH levels in healthy H9c2 cells were observed as 68 U/mL.
Graphical Representation of Lactate Dehydrogenase Assay (LDH). Y-axis: LDH Activity (U/mL); X-axis: Sample Code. All Experiments were Done in Triplicate, and Results are Represented as Mean ± SE. One-way Analysis of Variance (ANOVA) and Dunnett’s Test were Performed to Analyse Data. *p < .001 Compared to the Control Group.
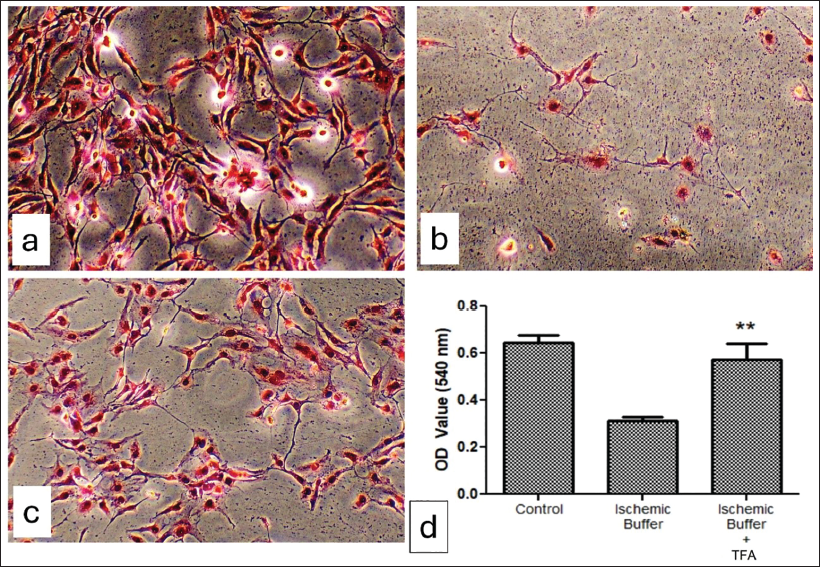
Sirius Red Staining Analysis
Microscopic examination of Sirius Red-stained H9c2 cells demonstrated inhibition of collagen degradation following the treatment using TFA. Degradation of collagen fibres in ischaemic controls caused the heart muscle cells to degenerate, indicating the possibility of ischaemic heart dysfunction. Treatment using 50 µg/mL TFA reduced the collagen degradation, resulting in a typical arrangement of muscle cells and fibrous network compared to the untreated group. OD values of supernatant confirm the inhibition of collagen degradation in the TFA-treated groups, as we recorded a value of 0.52, which is significantly higher compared to the untreated group (OD value −0.334) (Figure 5).
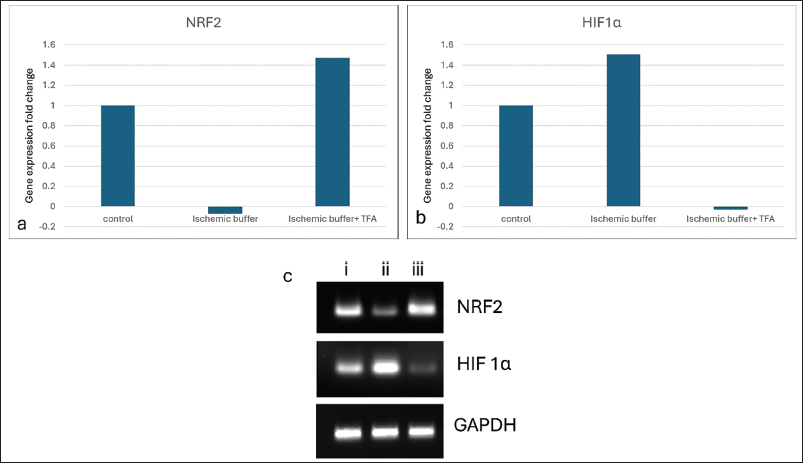
Quantitative Real-time Polymerase Chain Reaction Analysis
We adopted qRT-PCR analysis to investigate the expression profile of NRF2 and HIF1α in ischaemia-induced H9c2 cells (Figure 6). The housekeeping gene GAPDH serves to standardise the data obtained from the investigation. Ischaemic H9c2 cells demonstrated downregulation of the NRF2 gene in IHD. Conversely, the fold change exceeded 1.4 when ischaemic cells were treated with 50 µg/mL TFA. The expression level of NRF2 in the TFA-treated group was elevated compared to the control cells, confirming the cardioprotective mechanism enhanced by TFA. The expression of the HIF1α gene was inhibited in TFA-treated ischaemic H9c2 cells. HIF1α expression was raised in ischaemic cells, denoting cellular damage, which may have resulted from stress induced during the experiment.
Discussion
In recent decades, there has been a growing interest in TFA due to its significant pharmacological effects and diverse biological functions. Ferulic acid occurs naturally in fruits, vegetables, grains and a few herbs (Chaudhary et al., 2019; Frismantiene et al., 2018; Raj & Singh, 2022). Angelica sinensis, a Chinese medicinal herb abundant in ferulic acid, is frequently employed in the treatment of tumours, as well as cardiovascular and cerebrovascular disorders (Chen et al., 2024). Those features make it a potential candidate for therapeutic uses, especially in IHD, where oxidative stress and inflammation are the primary pathogenic factors. Additionally, another anti-atherosclerotic property, such as lipid-lowering, influences the management of IHD (Thapliyal et al., 2021). Consequently, we attempted to examine the anti-ischaemic potency of TFA concerning cardiac diseases and its cytotoxicity by various in vitro experiments.
Despite its considerable therapeutic potential, the clinical application of ferulic acid confronts numerous formidable challenges, including toxicity. In our study, TFA demonstrated a dose-dependent increase in cytotoxicity, although it did not exceed the threshold level. The viability of healthy H9c2 cells exhibited variation below 20% even at the concentrations of 100 µg/mL. The cytotoxicity assay performed by Choi et al. reported that ferulic acid at around 300 µg/mL concentration exhibited no significant toxicity in three types of cell lines and blood cells (Choi et al., 2018; Sun et al., 2021). Reported that 100 µM does not demonstrate toxicity to H9c2 cells in the MTT assay (Sun et al., 2021). A cytotoxicity level below 25% is considered relatively safe; therefore, the average dose of 50 µg/mL, which demonstrated 22% cytotoxicity, was used for subsequent tests.
Cardioprotective efficacy of 50 µg/mL TFA was 86%. Ischaemia-induced H9c2 cells exhibited 50% viability, the condition of which has been significantly improved by TFA at the various doses 12.5, 25 and 50 µg/mL. According to the previous studies, both pre- and co-administration of p-Coumaric acid (pCA) significantly reduced the cell toxicity induced by Dox in H9c2 cell lines. Pre-treatment of pCA helped to retain 86% cell viability, and co-administration of pCA helped to retain the cell viability of 84% (Chacko et al., 2015). Root flavonoids derived from Potentilla anserina L. improved cell viability of H9c2 cells from 70% to 80% while the dose of 52.08 µg/mL was used (Yan et al., 2025). Ferulic acid-mediated free radical scavenging shields the essential macromolecules from damage and enhances the activities of natural antioxidant enzymes, hence inhibiting cell apoptotic activities, a primary contributor to the occurrence of IHD (Purushothaman et al., 2024). Enhancement in the viability of ischaemic H9c2 cells observed in our results demonstrated the anti-apoptotic property of TFA.
LDH is found in various tissues, and it is released into the peripheral blood stream upon cellular damage. Knowledge about the impact of LDH levels in myocardial ischaemia is limited; however, it serves as an indicator of heart failure following myocardial infarction (Zhang et al., 2024). The LDH level was significantly increased to around 9,000 U/L upon the induction of ischaemia in cardiomyocytes using 200 µmol/L cobalt chloride (CoCl2) (Yue et al., 2024). IHD induced using hydrogen peroxide (150 µM) showed increased LDH levels to 600 U/L in H9c2 cells (Wang et al., 2019). In our investigation, ischaemia-induced H9c2 cells showed 356 U/mL of LDH activity. TFA treatment at a 50 µg/mL concentration showed a reduced level of LDH activity to 239 U/mL. Prior studies demonstrated that pre-treatment with 20 µmol/L Kaempferol notably increased cell viability, and so the exogenous LDH levels were lower, approximately 125 U/L. Kaempferol, a plant flavonoid, offers cardioprotective properties and demonstrates a beneficial effect on myocardial infarction (Yue et al., 2024). Araloside C (the most abundant triterpenoid compound present in Aralia elata), at a concentration of 25 µM, protects H9c2 cells against oxidative stress; the LDH levels were reduced to 400 U/L upon treatment (Wang et al., 2019). So, LDH assay results confirm that TFA treatment to ischaemic H9c2 cells inhibits cellular damage.
The disruption of the extracellular matrix (ECM) is shown to be the primary cause of myocardial damage. The composition and regulatory functioning mediated by the ECM control the maintenance of collagen (Kim et al., 2000). We detected collagen fibre degradation in ischaemic cells, indicating the possibility of ischaemic heart failure due to disorganised cardiac muscle cells, resulting in ventricular dilation and systolic dysfunction (Zuchi et al., 2020). Moreover, oxidative stress-induced muscle damage in the heart is another cause of ischaemia, leading to alterations in myocardial structure and function (Duan et al., 2024). Cardiac muscle damage may severely affect the pumping rate, which may further affect the oxygen supply and cause heart failure (Boyette & Manna, 2018). Our investigation indicates that TFA exhibits potential efficacy in reducing ischaemia-induced collagen degradation and preserving tissue integrity, as evidenced by the Sirius Red staining assay. Additional quantitative analysis and larger sample sizes are recommended to validate these findings.
NRF2 is a transcription factor that regulates the cellular defence against toxic and oxidative stress through the expression of genes involved in oxidative stress response and drug detoxification. NRF2 is involved in many other cellular processes, including metabolism and inflammation, and its functions are beyond the originally envisioned (Morgenstern et al., 2024). NRF2 gene expression in the ischaemic group was highly suppressed compared to the control. We observed elevated levels of NRF2 gene expression in the TFA-treated ischaemic H9c2 cells. Under normal conditions, NRF2, found in the cytoplasm, undergoes ubiquitination by the cullin 3 (Cul3) protein. Kelch-like ECH-associated protein 1 (Keap1) is involved in the ubiquitination process by acting as a substrate adaptor for NRF2, facilitating its degradation. Consequently, NRF2 has an indeterminate lifespan in the cytoplasm. During oxidative stress, as a defence process, elevated levels of NRF2 in the cytoplasm subsequently translocate to the nucleus, where it combines with Maf proteins to form heterodimers. The binding of heterodimers to antioxidant response elements (AREs) initiates the transcription of several antioxidative genes. Thus, the NRF2–Keap1 pathway constitutes the initial line of defence against ROS-mediated cardiac injury (Ngo & Duennwald, 2022; Singh et al., 2022). It has been reported that FA prevents neuronal degeneration by activating the NRF2 pathway. Unsaturated side chain and phenolic nucleus in the ferulic acid produce a phenoxy radical and contribute to free radical scavenging potential (Singh et al., 2022). Interestingly, we observed the fold change greater than 1.4 upon TFA treatment, which was higher than the levels in the control group, indicating the cardioprotective efficacy of TFA.
Hypoxia-inducible factor (HIF)-α, a transcription factor, plays a crucial role in myocardial ischaemia by regulating the hypoxia-responsive genes (Sato & Takeda, 2023). Accumulated HIF-α is a key sensor of low oxygen levels, because HIF-α proteins are continuously degraded by an oxygen-dependent degradation pathway in normal conditions. If oxygen depletion occurs, HIFα occurs, and its interaction with p300 and CBP transcriptional co-activators dimerises with HIF1β and translocates to the nucleus. The binding of the HIF dimer to specific DNA sequences promotes the transcription of genes involved in adaptive responses to cope with low oxygen levels. We observed elevated HIF1α expression in ischaemia-induced cardiomyocytes with a fold change of 1.5, indicating severe ischaemic injury, which might be caused by the cellular damage as suggested by Sirius Red staining analysis. A study by Sato and Takeda (2023) reported that the overexpression of HIF1α may lead to adverse outcomes, including cardiac rupture in some cases. Conversely, downregulation of HIF1α prevents P53-dependent apoptosis following myocardial infarction (Ikeda et al., 2021). We observed suppression of the HIF1α gene in TFA-treated ischaemic H9c2 cells, which denotes the therapeutic efficacy of TFA against ischaemic cellular damage. FA can suppress the HIF1 signalling pathway and inhibit ferroptosis and apoptosis, thus improving lipid metabolism and treating atherosclerosis (Wu et al., 2025). In vitro study using a foam cell model demonstrated FA-induced downregulation of HIF1α expression by upregulating GPX4 and Bcl2 pathways, which reduced the lipid accumulation and ROS content, whereas it improved the mitochondrial function and cell viability (Wu et al., 2025). The findings of the in vitro analysis conducted in this research demonstrate the anti-apoptotic and anti-ferroptotic efficacy of TFA in treating myocardial cellular damage that occurs in IHD. A dose of 50 µg/mL TFA exhibited safe and effective cardioprotective properties by preserving cardiac muscle organisation via inhibiting collagen degradation, which may lead to cellular damage.
We have observed elevated NRF2 gene expression and suppression of HIF1α in TFA-treated ischaemic H9c2 cells. These molecular changes were accompanied by improved cell viability, reduced LDH release and preservation of collagen organisation. These expressions clearly indicate a functional protective phenotype. While these findings suggest that modulation of oxidative stress and hypoxia-responsive pathways may contribute to the cardioprotective effects of TFA, the present study demonstrates an association rather than a direct mechanistic relationship. Existing literature has demonstrated the cardioprotective potential of ferulic acid and related phenolic compounds primarily through antioxidant activity, ferroptosis inhibition and metabolic regulation, often implicating pathways such as AMPK, MAPK or inflammatory mediators. However, limited studies have examined the coordinated modulation of oxidative stress and hypoxia-responsive transcription factors in ischaemic cardiomyoblasts. In this context, the present study advances knowledge by showing that TFA simultaneously upregulates NRF2 and suppresses HIF1α, two key regulators of cellular responses to oxidative stress and hypoxia, which are maintained across multiple cell types. Although these pathways represent general stress response mechanisms rather than cardiac-specific signalling, their regulation in cardiomyoblasts is highly relevant to IHD. Additional investigations using pathway-specific inhibitors, gene knockdown strategies, or in vivo ischaemia models are necessary to establish whether NRF2 activation and HIF1α suppression directly mediate the observed functional cardioprotection.
Conclusion
The therapeutic properties of ferulic acid are being investigated for the treatment of numerous disorders. TFA, a stable form of ferulic acid, has gained interest in the treatment of ischaemic stroke due to its stability and absorption properties. The preservation of cardiomyocyte structure and viability under ischaemic conditions may contribute to improved cellular resilience to hypoxic stress; however, direct effects on oxygen transport or vascular function were not assessed in this study. Overall, the current study investigated TFA, which demonstrated cellular-level cardioprotective effects by upregulating gene expression that mitigates oxidative stress and inflammation, thereby indicating a novel potential for a comprehensive evaluation of TFA. Moreover, the present study did not include transcriptional validation or pathway-specific functional assays. Therefore, additional studies incorporating protein expression analysis and targeted mechanistic assays are warranted, together with in vivo testing, to further confirm the functional significance of these gene expression changes.
Summary
This study evaluated the cardioprotective potential of TFA in ischaemia-induced H9c2 cardiomyoblasts. TFA showed minimal cytotoxicity and significantly improved cell viability to 86% at 50 µg/mL while reducing LDH activity, indicating reduced cellular injury. Sirius Red staining confirmed inhibition of collagen degradation, preserving cardiac muscle structure. Gene expression analysis revealed that TFA upregulated NRF2 and suppressed HIF1α, suggesting its role in mitigating oxidative stress and hypoxia-induced damage. Overall, TFA demonstrated strong cardioprotective effects, highlighting its potential as a natural therapeutic compound for managing IHD.
Footnotes
Abbreviations
ACE: Angiotensin-converting enzyme; AMPK: Adenosine monophosphate-activated protein kinase; ANOVA: Analysis of variance; ARE: Antioxidant response element; ATP: Adenosine triphosphate; Bcl2: B-cell lymphoma 2; cDNA: Complementary deoxyribonucleic acid; CoCl2: Cobalt(II) chloride; Cul3: Cullin 3; DGKC: Deutsche Gesellschaft für Klinische Chemie; DMEM: Dulbecco’s Modified Eagle’s Medium; DMSO: Dimethyl sulfoxide; DNA: Deoxyribonucleic acid; DOX: Doxorubicin; ECM: Extracellular matrix; FA: Ferulic acid; FBS: Foetal bovine serum; GPX4: Glutathione peroxidase 4; H9c2: Rat cardiac myoblast cell line; HEPES: 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; HIF1α: Hypoxia-inducible factor 1 alpha; IHD: Ischaemic heart disease; Keap1: Kelch-like ECH-associated protein 1; LDH: Lactate dehydrogenase; mRNA: Messenger ribonucleic acid; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; Na+/K+ ATPase: Sodium-potassium adenosine triphosphatase; NRF2: Nuclear factor erythroid 2-related factor 2; OD: Optical density; pCA: p-Coumaric acid; PBS: Phosphate-buffered saline; qRT-PCR: Quantitative real-time polymerase chain reaction; RNA: Ribonucleic acid; ROS: Reactive oxygen species; SEM (or SE): Standard error of mean; TFA: Trans-ferulic acid.
Declaration of Conflict of Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Ethical Approval
This research does not require ethical approval.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
Informed Consent
NA.