
Abstract
Background
Tactile stimulation (TS) is a positive experience that mimics maternal licking and grooming in the rat. Since the brain is extremely plastic in early life, TS during this period may modulate normal brain organisation and could enhance cognitive and/or motor abilities.
Purpose
This study is designed to evaluate the possible changes in exploratory behaviour, emotionality and learning and memory following repetitive TS during periadolescent period.
Methods
Male albino Wistar rats were grouped into control and TS groups. The TS group animals were exposed to repetitive TS (15 min/session), three times/day from postnatal day 28–43. After this, exploratory behaviour, emotionality and object recognition memory were tested using open field (OF), elevated plus maze (EPM) and novel object recognition (NORT) tests by a computerised video-tracking system.
Results
TS increased rearing number, rearing rate, total stretching, total stretching duration, stretching rate, mean speed and parallelism index in rats during the OF test. However, total distance travelled, global activity and total zone transitions were comparable in both groups during the OF test. TS significantly increased stretching duration and decreased entries to the closed arm during the EPM test. It slightly increased the percentage of time spent and distance travelled on the open arm in TS group. Total distance travelled, total zone transitions and global activity were comparable in both groups during the EPM test. TS mildly increased recognition index during NORT.
Conclusion
Tactile stimulation exposed animals demonstrated increased exploratory, risk assessment and decision-making behaviours on OF test suggesting a possible positive influence of TS on these behaviours. However, TS only mildly influenced emotionality and object recognition memory, and it did not alter general locomotor behaviours in rats.
Introduction
The periadolescent period is the development stage during which critical changes occur in the brain. 1 Pieces of evidence that emerged as a result of neurobehavioural, morphological and pharmacological research suggests that the brain remains under construction during adolescence. 2 Along with other factors psychological status of an individual plays a very important role in brain development and maturation during this period. 2 It is reported that consolidation of control mechanisms of the brain and nervous system, in general, occurs during adolescence through the maturation of neurocircuitry and myelination. 3 Multiple research reports demonstrate evidence to support the argument that the brain is still under construction during the periadolescent period.3, 4 Tubulinogenesis, axonogenesis and synaptogenesis are usually completely accomplished during prenatal and early postnatal life. However, research points out that myelinogenesis remains active until adolescence life suggesting any positive and negative psychological state or other related factors can potentially modulate the myelination process in the nervous system, resulting in changes in brain structure and function leading to possible long-term implications in behaviour. 5
There have been multiple efforts made to understand impulsive behaviour in young adolescents and strategies to control or overcome this behaviour. Results from neurochemical studies reveal that glutamatergic neurotransmission is almost completely accomplished during prenatal and early postnatal life. 6 However, gamma-aminobutyric acid (GABA)-ergic neurotransmission especially in the prefrontal cortex remains under development during adolescence. This delayed development of GABAergic neurotransmission is attributed to neurobehavioural excitement including novelty-seeking, euphoria and risk-taking behaviour in adolescents. 6 Perhaps, adolescents are novelty-seeking and risk-taking individuals, and they are more likely to weigh positive experiences more heavily and negative experiences less than adults. Hence, it is important to know and study the factors that can alter such behavioural patterns.
Tactile stimulation (TS) is a positive early life experience that mimics maternal licking and grooming in the rat. Since the brain is extremely plastic in early life, TS during this period may modulate normal brain organisation and could enhance cognitive and/or motor abilities. 7 Reports suggest that to generate longer-term changes in behaviour, experiences must produce stable changes in neuronal morphology and synaptic connectivity. 8 It is reported that when normal animals were exposed to TS from P3 to P21 resulted in the proliferation of neuronal parameters in mPFC (medial prefrontal cortex). 9 External TS from P8 to P21 for 10 minutes produced long-term facilitative effects in spatial working memory-related neural function in rats. 10 C-Fos is thought to have role in neuronal activity, neurogenesis, learning, and memory and TS is reported to stimulate the c-Fos expression in rat brains. 11 Recent reports demonstrate that TS offers therapeutic-like effects in case of iron-deficient disorders postnatally demonstrating the remarkable responsiveness of the brain to its interaction with the environment. 12 TS has been mainly used to compensate for known or predicted deficits, but, repeated TS effects on the normal brain and related behaviours have not been investigated. Specifically, changes in exploratory behaviour, emotionality, object recognition memory and general locomotor behaviour, following TS, have not been documented well in periadolescent rats. Hence, this study is designed to evaluate the possible changes in the aforementioned behaviours in periadolescent rats following a repetitive positive experience such as TS.
Methods
Animals and Maintenance
This experimental study was done on male Wistar rat pups. To generate this animal line, healthy male and female albino rats of Wistar strain (3 months of age), weighing 200–220 g, were used. They were maintained at the Central Animal Research Facility (CARF), Ras Al Khaimah Medical and Health Sciences University, UAE, and were placed (3/cage) in polypropylene cages (41 cm × 28 cm × 14 cm) in the CARF. The Institutional Research Ethics Committee has approved all the procedures used in the study (RAKMHSU-REC-001-2020-UG-M). Three female rats in full estrous were selected and paired with a male for a maximum of 3 hours, typically between 10:00 and 13:00 hours. Once the pregnancy was confirmed, each pregnant dam was separately maintained. After parturition, male pups were selected and separately put in a cage with the dam. They were randomly allocated to the experimental groups at PN-21 and maintained up to PN-27. The experiments commenced at PN-28. Animals were maintained in a 12:12 h L:D environment, in an air-conditioned room and fed with water and standard food ad libitum. Sample size was calculated as per the following formula; SD 2 (Zα/2 + Zβ) 2 /d 2 ). The calculated sample size was 10 per group.
Experimental Groups and Study Design
A total of 20 animals were used for the study and were randomly assigned into the following groups: Control (n = 10): Animals of this group remained in the home cage throughout the experimental period. TS Exposed (n =10): Animals of this group were exposed to TS from postnatal day 28–43 for 15 min/session, three times/day with 2 hours intervals between each session. TS was given in their home cage in the animal housing room by brushing all pups with a soft feather-like duster for 15 minutes. After the experimental period, all the animals were tested for exploratory behaviour, emotionality, object recognition memory and general locomotor behaviour. A computerised animal behaviour video-tracking system (SMART- Panlab, Version 3.0-Super Pack, Barcelona, Spain) was used for the behavioural studies.
Open Field (OF) Test
This test provides both qualitative and quantitative measurements of exploratory and locomotor behaviour in rats. A black square box made of material that is non-absorbent for odours (Square arena, LE800SC, Panlab, Barcelona, Spain) measuring 90 (W) cm × 90 (D) cm × 40 (H) cm was used for this purpose. The top is left uncovered, and it was placed on the floor to imitate an OF. The box was divided into nine imaginary zones to make it easier to count the zone transitions as well as the amount of time spent in each zone. In the testing session, the animals were brought in their house cage from their housing room into the testing room. A single rat was removed from the home cage by gently grasping the tail and placing it on the floor of the apparatus facing the wall, while concurrently activating the SMART software to record their activity. Each animal was allowed to explore for 5 minutes with free and uninterrupted movement throughout the quadrants of the apparatus during which a video camera recorded its activities. After the testing period, the animal was gently picked up and returned to its home cage. The floor and walls of the apparatus was cleaned with 70% alcohol solution using paper towels. The alcohol solution was allowed to completely dry before testing other rats. These procedures were repeated with all animals. Emotionality-related parameters like the percentage of time spent in the centre/peripheral zone, entries to centre/peripheral zones and distance travelled in the centre/peripheral zones were recorded. Exploratory and risk assessment behaviour parameters like rearing, stretching and freezing behaviour like total immobility duration were recorded. General locomotor behaviours like mean speed, total distance travelled, total zone transitions, parallelism index and global activity were also recorded and reported. 13
Elevated Plus Maze (EPM) Test
An EPM (LE840, Panlab, Barcelona, Spain) in which two of its opposite arms were closed (closed by black walls) and the other two opposite arms were open with a transparent railing to prevent the rats from falling was used for this test. The maze measured, 100 (W) cm × 100 (D) cm × 100 (H) cm. It has two arms measuring 45 cm and a centre region measuring 10 cm on all four sides which connects to four arms of the apparatus. The wall height of the closed arm was 35 cm and the entire apparatus was elevated 65 cm from the floor of the testing room. During the EPM test, each rat was placed on the centre of the maze and was allowed to explore it freely for 5 min. The path taken by the rat was recorded using a video camera. Emotionality-related parameters like percentage of time spent in the open arm, centre and closed arm, entries to various arms and distance travelled in various arms were recorded. Exploratory and risk assessment behaviour parameters like rearing, stretching and freezing behaviour like total immobility duration were also recorded. General locomotor behaviours like total distance travelled, total zone transitions and global activity were also recorded. After each trial, the number of faecal pellets and urination was noted, and all arms of the EPM were cleaned with 70% alcohol solution. 14
Novel Object Recognition Test (NORT)
A black square arena (LE800SC, Panlab, Barcelona, Spain) measuring 90 (W) cm × 90 (D) cm × 40 (H) cm was used for this purpose. It was divided into four quarters measuring 45 (W) cm× 45 (D) cm × 40 (H) cm by placing two dividers. One of the quarters was used as an experiment arena for NORT. This test is commonly used to investigate different aspects of learning and memory in rats. This test was carried out in three stages and completed in three days: familiarisation day, object recognition day and testing day. (a) For familiarisation, each rat was allowed to freely explore the testing arena for three minutes. Seventy per cent of alcohol solution was used to clean the apparatus after each familiarisation. (b) Object recognition, those objects that can be easily discriminated by the rats with similar complexities (colour, texture, brightness, shape, size) were used to rule out any object preferences that may confound the results. 15 For this assay, two similar objects were selected and placed on the corners of the same side (i.e., northwest (NW) corner and northeast (NE) corner) and the rats were allowed to explore for three minutes each. Time spent with each object, was noted. After each trial, the apparatus was thoroughly cleaned using 70% alcohol solution. (c) On the testing day, one object out of the two similar objects, was removed and replaced with a novel object. In this trial, each rat was allowed to freely explore the testing arena containing the objects for three minutes. Time spent with each object, was noted. The apparatus was cleaned thoroughly using 70% alcohol at the end of each trial. 16 Steps (b) and (c) were video recorded using a computerised animal behaviour video-tracking data acquisition system (SMART-Panlab, Barcelona, Spain). The NOR memory or cognition-related parameter such as the time spent on the novel object was recorded and by using this data the NOR index was determined and reported.
Statistical Analysis
The data is expressed as Mean ± SEM. Student ‘t’ test was used to analyse the data. A ‘p’ value ≤ 0.05 was considered as statistically significant. GraphPad Prism software (San Diego, CA, USA) was used for data analysis.
Results
Open Field Test
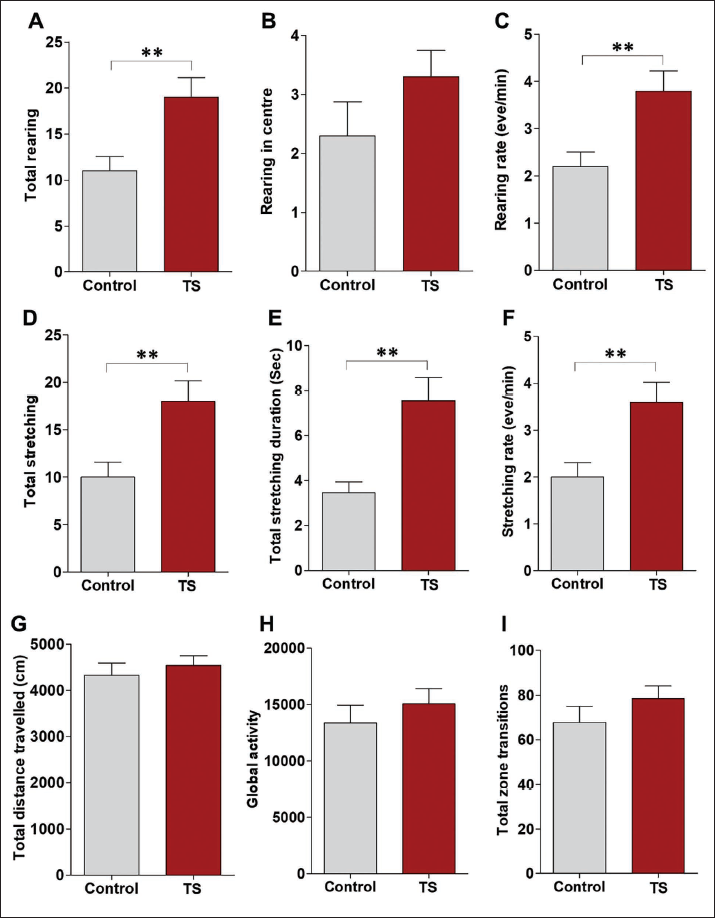
Rearing is considered to be one of the exploratory behaviours in rodents. The total number of rearing is observed to be significantly increased in the TS group (p < .001) (Figure 1a). Similarly, rearing rate is also found to be significantly increased (p < .001) in the TS group (Figure 1c). Rearing in the centre was found to be more with the TS group, but this change was not statistically significant (Figure 1b). Total stretching number (Figure 1d, p < .001), stretching duration (Figure 1e, p < .001) and stretching rate (Figure 1f, p < .001) were found to be more with the TS group compared to the control animals. Although the total distance travelled in the apparatus (Figure 1g), global activity in the OF (Figure 1h) and the total number of transitions (Figure 1i) were increased in the TS group; this difference was not statistically significant.
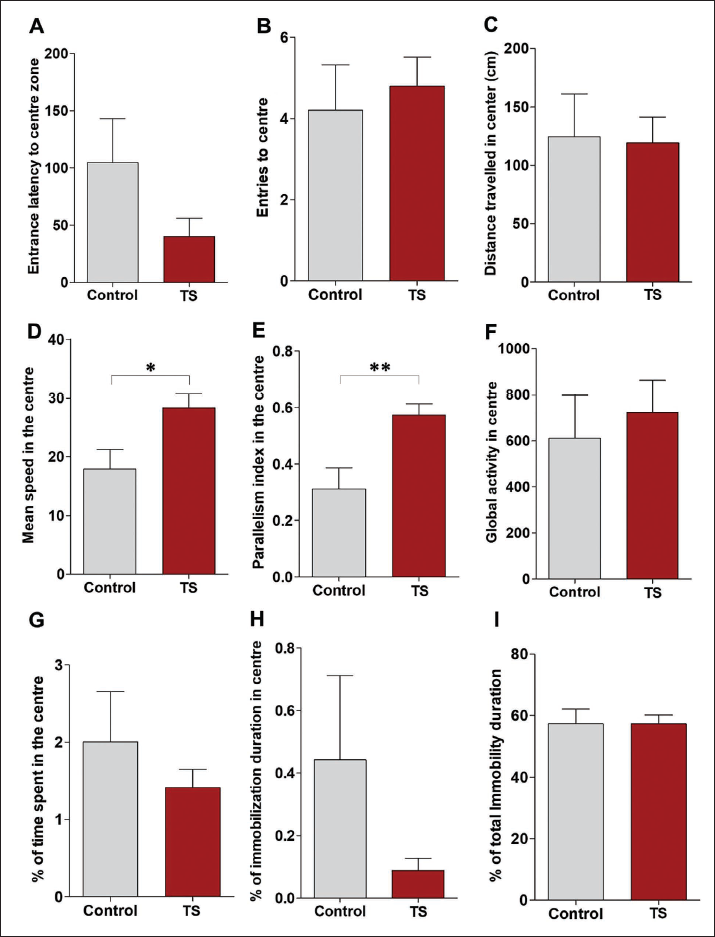
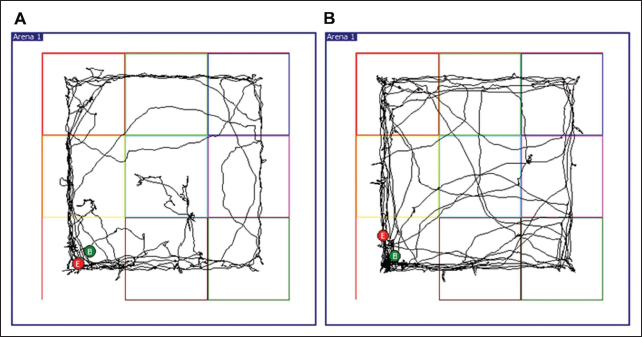
The latency to enter the centre zone was decreased in the TS group compared to the controls. However, this difference was not statistically significant (Figure 2a; p = .13). TS group animals entered the centre zone more compared to control animals, but this difference was also not found to be statistically significant (Figure 2b). Distance travelled in the centre was comparable in both groups (Figure 2c). However, the mean speed in the centre of TS group animals was significantly increased compared to controls (Figure 2d; p < .01). The parallelism index was also found to be higher with TS group animals with controls (Figure 2e; p < .001). Although global activity in the centre was increased with TS group animals, this difference was not statistically significant (Figure 2f). The percentage of time spent in the centre by TS group animals was decreased in the OF test, but this was not statistically significant (Figure 2g; p = .40). Interestingly, the immobilisation time was more with the control rats in the centre suggesting a freezing type of behaviour in these rats. Although the immobilisation time in the centre was found to be decreased in the TS group compared to the control group, this decrease was not found to be statistically significant (Figure 2h). Total immobilisation time was found to be comparable in both groups (Figure 2i). Furthermore, the real-time video tracking of control (Figure 3a) and TS group animals (Figure 3b) path during the OF test demonstrated that the TS group explored the apparatus well (Figure 3b) compared to the control group animals. The tracking analysis revealed that 20% of the control rats never visited the central square, while all TS group rats visited and crossed the centre square several times.
Elevated Plus Maze Test
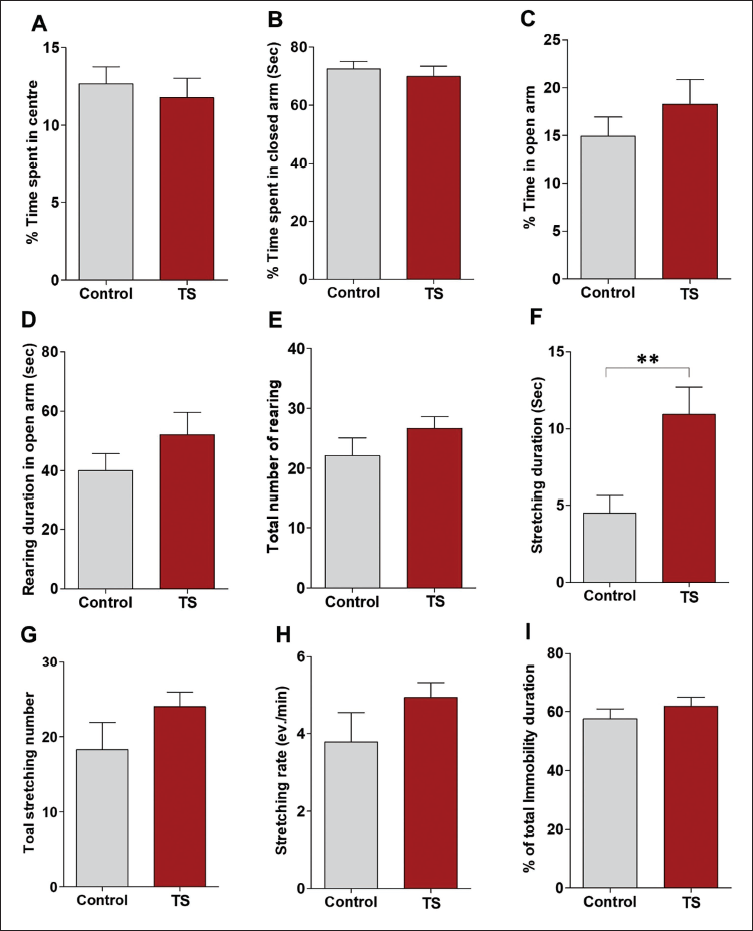
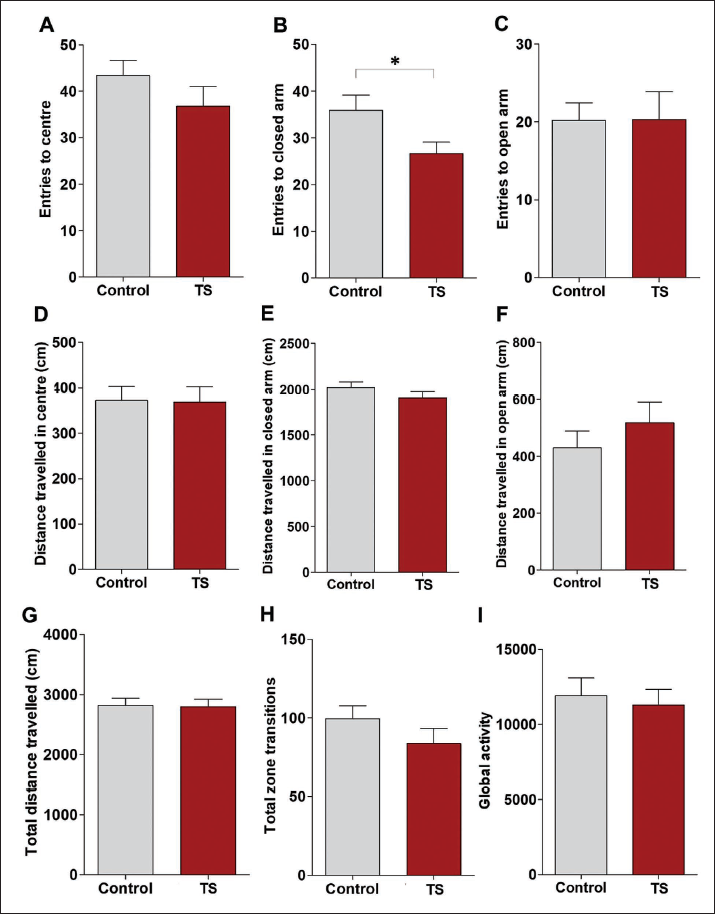
In the EPM experiment, the percentage of time spent in the centre (Figure 4a) and closed arm (Figure 4b) was slightly reduced in the TS group. However, this was not statistically significant. The percentage of time spent on the open arm was found to be more with the TS group, but the student t-test revealed no significant difference between the means of the two groups (Figure 4c). In the TS group, open arm rearing duration (Figure 4d), total rearing number (Figure 4e) and total stretching duration (Figure 4f) were observed to be increased, but only stretching duration in both the groups was found to be significantly different (Figure 4f; p < .001). Total stretching number (Figure 4g) and stretching rate (Figure 4h) were also found to be elevated in the TS group, but this was not statistically significant. The percentage of total immobility duration was comparable in both groups (Figure 4i).
Entries to the centre (Figure 5a) were decreased in the TS group compared to the control, but it was not statistically significant. However, TS group entries were significantly decreased in the closed arm (Figure 5b). Entries to open arms were comparable with both groups (Figure 5c). Distance travelled in the centre (Figure 5d) and closed arm (Figure 5e) was also comparable in both groups. However, the distance travelled in the open arm was increased in the TS group (Figure 5f), but this was not significant (p = .35). Total distance travelled (Figure 5g), total zone transition (Figure 5h) and global activity (Figure 5i) were comparable in both groups. Additionally, as depicted in Figure 6, the real-time video tracking of animal path during the EPM test revealed that both groups equally explored the apparatus well. However, the TS group explored the open arm better (Figure 6b) than the control rats (Figure 6a).
Representative Video Tracking of Control (a) and Tactile Stimulation Group (b) Animals During the EPM Test. Both Groups Have Explored the Apparatus Well. Although, the Tactile Stimulation Group Explored the Open Arm Better Than the Control Rats, This Difference Was Not Statistically Significant.

Novel Object Recognition Test
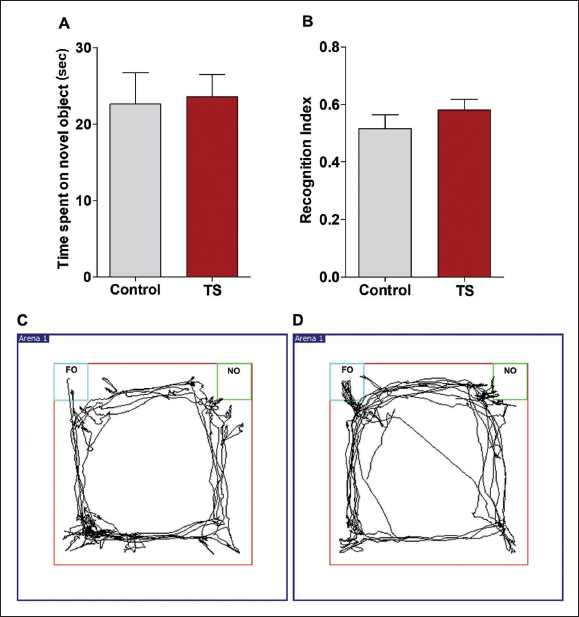
The time spent on novel object was slightly increased in TS group rats compared to control rats (Figure 7a). The recognition index was also increased in TS rats (Figure 7b). Nonetheless, these changes were not statistically significant. The real-time video tracking of animal trajectory demonstrated the higher attempts of the TS group (Figure 7d) to reach the novel object compared to the control group (Figure 7c) during the NORT.
Discussion
Based on the data acquired through the current experiments, this study has yielded four interesting findings. First, the TS modified exploratory and risk assessment behaviours such as rearing and stretching in the OF test. The general locomotor behaviours were not significantly modified except the speed and the parallelism index in the TS group. A similar finding was observed with the exploratory behaviour in the EPM test. However, the stretching duration was slightly modified in the TS group in the EPM test. The modification of the exploratory and risk assessment behaviours of the TS group in two of the behavioural analysis paradigms is an interesting observation. Second, emotionality-related parameters were not significantly modified in the TS group in the OF behavioural testing paradigm. However, it must be noted that during the OF test, while control rats showed freezing-like behaviour in the centre zone (which could be the reason for their higher times spent in the centre), the TS group did not significantly demonstrate this behaviour and explored the centre zone. Moreover, 20% of the control rats did not visit the central square while all TS group rats visited and crossed the centre square indirectly suggesting the possible decreased emotionality in these animals. Similarly, on the EPM, emotionality parameters such as the percentage of open-arm exploration time were only slightly reduced in the TS group. Third, all general locomotor-related behaviours were not modified due to TS. Fourth, although not significant TS stimulation did mildly influence the NOR memory in the TS group compared to the control group. An increased dose of TS stimulation that may modify the aforementioned behaviours needs further investigation.
Rearing behaviours involve animals standing upright on their hind paws in a vertical position. These behaviours are categorised as exploratory behaviours and also have been employed as indicators of anxiety in both the OF and the EPM by researchers. 13 Reduced rearing behaviour signifies an increase in anxiety levels in rodents. In this study, the TS group demonstrated increased rearing suggesting their possible decreased emotionality-like behaviour. Several research papers report the usefulness of including rearing in the comprehensive behaviour analysis of animals.17, 18 Studies have also demonstrated that there are two types of rearing such as supported and unsupported rearing give valuable information about an animal’s behavioural state. 19 Studies20, 21 have demonstrated that unsupported rearing loads onto the principal components similar to ‘time in centre’ a parameter typically associated with emotionality whereas supported rearing is more highly associated with a different principal component associated with distance or activity. In this study, it can be noted that unsupported rearing (rearing without contacting the walls of the arena) is mildly increased in TS exposed rats (rearing in the centre of the OF and/or on the open arm of EPM). Research has demonstrated that the hippocampus serves as a conflict resolution control centre in which it inhibits ongoing behaviour in a conflict situation and mediates exploratory behaviour if more information is required to resolve the conflict. 22 Therefore, rearing can be increased in situations of uncertainty wherein the hippocampus plays a significant role. It is argued that unsupported rearing behaviour is connected to intra- and infrapyramidal mossy fibres which may serve to transfer information via the perforant pathway to the CA3 region. To understand the modulatory role of TS stimulation on these pathways, further research is needed.
Anxiety can affect the performance of rodents in certain types of experiments. Increased anxiety diminishes attention and makes it difficult for the animal to respond properly to a novel stimulus. In similar research, studying anxiety and depression in repeatedly handled mice, it has been observed that repeatedly handling the mice improved decreased anxiety-like behaviour and increased spatial cognitive functions in the Y-maze test. 23 This study only partially corroborates with these findings as in this study recognition memory was only mildly improved in the TS group. TS is reported to have significant effects in neonatal rats. TS during the neonatal period helped rats to cope with stressful situations in adulthood and to prevent depression. 24
The object recognition test is an efficient and flexible method for studying learning and memory in rodents. While the NORT is simple and fast to use, it does have limitations. Although it is not possible to analyse potential differences in the rate of learning, it allows for studying individual phases of learning and memory, such as consolidation or recall. 16 In this study, the rats who received TS explored the novel object slightly higher than the control animals. Although this result was not statistically significant, the observation is interesting and requires further research. Reports demonstrate that unlike some of the other tests of memory, which can be attributed to one brain region, NORT appears to make use of multiple brain regions and neurotransmitter systems, including the hippocampus and perirhinal regions. 25 We believe that a positive change in NORT in TS group animals could be due to the possible improved learning and better memory consolidation. In this report, we could not categorically demonstrate our argument such as what molecular change/s in various brain regions or modification in neurotransmitter levels would have caused such a positive behavioural response in TS exposed animals. This may be considered as one of the limitations of the study.
The underlying mechanism responsible to modulates exploratory, emotional and/or cognitive behaviours following TS remains unclear. Perhaps, this could be via the modulation of the hippocampal formation. TS is conveyed and processed via serial and parallel pathways. In this study, TS was provided to the animals using a soft duster. Such types of innocuous mechanical stimuli are generally encoded by cutaneous low-threshold mechanoreceptors. They are transmitted via ascending somatosensory pathways to the primary somatosensory cortex (S1), where tactile signals are decoded and processed. 26 The tactile inputs are further processed and integrated along with inputs from other sensory modalities in higher-order association areas. 27 Reports suggest the role of the lateral entorhinal cortex (LEC), a component of the parahippocampal region is proposed to regulate the flow of both uni- and multimodal sensory information to the hippocampal formation which supports declarative memory by relational processing of spatial and non-spatial-information. 28 Evidence suggests that LEC is reciprocally connected to the primary somatosensory cortex (S1). 29 Its layer 2 reelin-expressing neurons relay processed sensory information to the hippocampal dentate gyrus (DG). 30 Presumably, DG granule cell activity can be altered by TS 31 or deprivation. 32 Memory and anxiety may be differentially modulated by DG neurons along the hippocampal dorsoventral axis, and it is possible that tactile input coverage on the DG via the LEC could be involved in shaping the cognitive and emotional processes in rats.33, 34 In order to categorically prove this happens after repetitive TS stimulation, we need to look at the histochemistry of the hippocampal formation and study the changes in dendritic arborisation which we could not include in this report is another limitation of the study. Although we could include 10 animals in each group, the behavioural variations in one group seem to be slightly higher in certain parameters (% time spent and immobility time on the centre of OF or open arm of EPM) which could have been avoided by including more animals in each group and that could be considered as another limitation of this study. A continuation of this study further evaluating the histochemistry markers, dendritic arborisation and/or neurotransmitter levels in the hippocampus or relevant other brain regions in the TS exposed group would help us to give a clearer explanation for the improved exploratory behaviour, mildly improved NOR memory and altered emotionality in TS exposed animals.
Conclusion
Tactile stimulation exposed animals demonstrated increased exploratory, risk assessment and decision-making behaviours in the OF behavioural testing, suggesting its possible positive influence on these behaviours. However, TS had only mild effects on emotionality and NOR memory in rats. The general locomotor behaviours of rats were not altered by TS. This study provides some evidence towards the positive effects of repetitive TS, a type of positive stimulation during a critical period of development which may influence exploratory and risk assessment behaviours in adolescence. However, further studies are warranted to categorically establish the underlying molecular and cellular mechanisms that occur in various brain regions as a result of repetitive TS eventually leading to behavioural changes in rats.
Footnotes
Authors’ Contribution
S.N.N. conceived of the presented idea, designed, directed the project and analysed data and edited the manuscript.
H.H.R., M.S., R.R. and A.F. performed various experiments with the help of S.N.N. and drafted the initial version of the manuscript.
All authors have contributed to and approved the manuscript’s final version.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
ICMJE Statement
This article complies with the International Committee of Medical Journal Editors (ICMJE) uniform requirements for the manuscript.
Statement of Ethics
The Institutional Research Ethics Committee has approved all the procedures used in the study (RAKMHSU-REC-001-2020-UG-M).