
Abstract
Background
Many military service members are victims of repetitive blast traumatic brain injuries (rbTBI) and endure diverse altered psychological and behavioural conditions during their lifetime. Some of these conditions include anxiety, post-traumatic stress and pain. Thus, this study attempts to fill the knowledge gap on enduring behavioural and neuroinflammatory marker alterations 1 month after rbTBI.
Purpose
Although previous rbTBI animal studies have shown behavioural and histopathological changes either a few days (acute) or many months (chronic) after trauma, knowledge related to post-traumatic changes during the intermediate timeframe, i.e. a month after rbTBI is less clear or unavailable.
Methods
Sprague–Dawley rats (male; n = 12) were assigned to either rbTBI or sham conditions. Animals assigned to the rbTBI group were subjected to 1 blast exposure per day for three consecutive days, while animals in the sham group were exposed to identical experimental conditions sans blast exposure. All animals were tested for anxiety at baseline. 30 days post-injury, animals were tested again for anxiety and paw thermal sensitivity, followed by brain harvest for immunohistochemical analyses.
Results
Animals exposed to rbTBI showed signs of anxiety-like behaviour on parameters of elevated plus-maze and behavioural signs of pain indicated by reduced thermal withdrawal latency of the forepaw. Histologically, brain sections from animals exposed to rbTBI showed a significantly increased number of microglial/macrophage and astrocytic counts in the medial prefrontal cortex.
Conclusion
Data from this initial preclinical study support the prevalence of putative anxiety-like behaviour, enhancement in forepaw thermal sensitivity and increase in the number of glial cells even 1 month after rbTBI. These findings have potential implications in the treatment evaluation of blast-exposed military and civilian populations and emphasise the need for devising protective measures for people susceptible to single or repeated exposures. A greater further understanding of rbTBI-related chronic concurrent behavioural and neuropathological sequela is warranted.
Introduction
Many service members returning from war zones in Afghanistan and the Middle East endure the long-term consequences and comorbidities following blast traumatic brain injury (bTBI).1, 2 Post-traumatic stress disorder (PTSD), anxiety, depression, substance use and sensory complications such as pain, hearing and balance problems3–6 are some common comorbidities following bTBI. About 14%-16% of those deployed to these conflicts develop PTSD or depression. 7 Compared to those with single TBIs, the frequency of anxiety, PTSD and other symptoms was shown to increase in members with multiple TBIs. 6 Furthermore, in members who experienced repeated blast TBI (rbTBI), an increased number and severity of concussive-like symptoms, 8 metabolic changes 9 and other neurological aberrations were also reported. 10
Many members also experience multifocal pain, 11 including musculoskeletal, neuropathic, and phantom pain, following blast exposure. 12 Chronic pain is a familiar complication following TBI, with a prevalence rate of 43.1% in Veterans, and can be independent of psychological disorder diagnosis. 13 Indeed, subjects referred for the management of chronic pain often have a history of symptoms suggestive of an untreated mild TBI (mTBI). 14 Additionally, compromised pain anticipation and modulation response were also reported in those with blast-related mTBI. 15
While postmortem analysis of brain tissue of blast-exposed Veterans revealed glial reactivity and degenerative changes,16–18 inflammation in the form of innate immunity modulation, inflammatory cell activation and inflammatory mediator release has also been implicated in conditions related to anxiety and PTSD. 19 Furthermore, the complex interplay between neurons, glial cells and mediators of inflammation 20 in the pathogenesis and maintenance of neuropathic pain has also been proposed.20–22
There is, however, a paucity of rbTBI-related studies showing concurrent behavioural and histopathological changes. For instance, the presence of reactive oxygen species, diffuse axonal injury, neurodegeneration and behavioural equivalents of motor impairment were shown as early as 5 days with no details on anxiety behaviour in a mouse model of rbTBI. 23 Other investigators have shown enhanced glial proliferation and altered spleen cytokine levels 7 days after rbTBI but no behavioural findings. 24 Meanwhile, Heyburn et al. 25 in a rat model of rbTBI showed differential effects of two low-level pressures and suggested neurovascular disruption in the form of enhanced glial fibrillary acidic protein (GFAP) and altered mechanosensitivity but reported no behavioural changes. Kamnaksh et al., 26 in their rat rbTBI study, showed the highest depression and anxiety-like behaviour 1 day after bTBI and suggested a potential conditioning phenomenon. Taken together, these studies support bTBI-induced acute changes but endorse limited to no knowledge of concurrent behavioural proxies of anxiety, pain, or neuroinflammation at a later time.
Meanwhile, other investigators have studied behavioural changes following rbTBI in isolation from concurrent inflammatory changes. Elder et al. 27 reported traits of anxiety and other behavioural surrogates of PTSD following repeated low-level blast overpressure exposure in rats studied for 7-25 weeks. Subsequently, at 28-35 weeks, they reported anxiety-like behaviour and altered fear response. 28 At 8 months after rbTBI, anxiety-like behaviour (open field) following exposure to a single predator scent was also reported. 29 Although these studies offer knowledge related to the anxiety-like behaviour occurring from at 7 weeks to 8 months post-rbTBI, the earlier precursors of these behaviours occurring a month after rbTBI with parallel neuroinflammatory changes remain unknown. The complex web of bTBI-related chronic behavioural changes potentially driven by cellular events highlights the need for preclinical models that can mimic the behavioural and histopathological sequelae of rbTBI. Accordingly, this initial study assessed co-occurring behavioural and neuropathological aberrations at 30-day time point after rbTBI.
Methods
All experimental procedures were approved by the Wayne State University Institutional Animal Care and Use Committee. Also, relevant guidelines in the Guide for the Care and Use of Laboratory Animals (2011) were followed. 30
All experiments were performed using twelve 55-70-day-old male Sprague–Dawley rats (Charles River Laboratories, Raleigh, NC) with an average weight of 311 ± 9.73 g before rbTBI. Upon arrival, all animals were pair-housed and remained so for the study duration. They were allowed to acclimate in a humidity and temperature-controlled vivarium room with a reverse light/dark cycle (lights off at 6:00 and on at 18:00) for 5-7 days. All rats were given ad libitum access to food and water. Following acclimation, and assessment of baseline anxiety-like behaviour on an elevated plus-maze (EPM) located in the behavioural suite that maintains identical climate conditions as their housing room they were assigned randomly to rbTBI (n = 6) or sham (n = 6) exposure conditions. The following day, they were subjected to rbTBI or sham procedures as described below. Following the injury procedure and testing for surface righting reflex, they were left undisturbed for 1 month and were then reassessed for anxiety-like behaviour on the EPM and evaluated for paw thermal sensitivity changes. All animals were also weighed at baseline and after a month. All behaviour procedures were performed during the animal’s active phase (between 9:00 and 17:00 h). After paw thermal sensitivity testing, all animals were transcardially perfused and had their brains harvested. An overview of this timeline is shown in Figure 1.

rbTBI Induction
rbTBI was induced using the shock tube system within the Wayne State University Department of Biomedical Engineering and has previously been used to study bTBI and related comorbidities.31–36 On each of three consecutive days of injury or sham procedures, they were transported in a temperature-controlled vehicle from the vivarium to the shock tube facility, a short distance of 1.3 miles. In the shock tube facility, they were allowed to habituate for at least 30 min. rbTBI animals were subjected to a single blast overpressure exposure per day for 3 consecutive days. Sham animals underwent all procedures sans blast pressure exposure. For rbTBI and sham exposures, animals were anesthetised (isoflurane 4% and 0.6 L/min oxygen) for 4 min in an induction chamber. Then, while maintaining anaesthesia for two additional minutes via a nose cone, they were harnessed to a sled without chest protection to reduce the dynamic pressure load and prevent the translation of the entire body. Rats were positioned inside the shock tube with a rostral-caudal orientation toward the shock wave. The shock front and the dynamic overpressure were generated by a custom-built shock tube (12′′ diameter, 193′′ long shock-producing tube, ORA Inc., Fredericksburg, VA). A pencil probe (PCB Piezotronics) was used to measure the static overpressure on the rat. The pencil probe was housed in an aluminium rod that supports the rat-holding platform. Desired peak static overpressures were produced with compressed helium and calibrated Mylar sheets (GE Richards Graphics Supplies Inc., Landisville, PA) to simulate a free field blast wave. A Dewetron data acquisition system (East Greenwich, RI) was used to collect pressure measurements at 100,000 Hz. The peak pencil pressure near the animal was 28.08 psi (±0.86). Such high pressures were previously shown to produce hemodynamic, 37 auditory, inflammatory and neuronal injury changes.31, 32 Immediately following the bTBI or sham procedure, rats were assessed for surface righting latency, considered an indirect indicator of loss of consciousness. All animals were returned to the vivarium following the conclusion of procedures for the day. After three consecutive days of blast TBI or sham procedures, they were left undisturbed for 30 days until post-injury anxiety and thermal sensitivity assessment.
Anxiety Assessment on the Elevated Plus Maze (EPM)
Anxiety-like behaviour before and 30 days post-rbTBI or sham procedures was assessed on the EPM for 5 min.
38
The EPM (Coulbourn Instruments, Allentown, PA) was fabricated with black Plexiglas and laid out in the shape of a plus sign with two open and two closed arms opposing each other.
39
Each open and closed arm measuring 10 cm wide and 45 cm long is surrounded by a 1 cm tall edge and a 30 cm tall wall respectively. The runway of the maze is elevated 52 cm off the ground. A ceiling-mounted overhead camera in the behavioural room connected to a computer in the anteroom allowed for a continuous undisturbed digital recording of the behaviour of the rat. Animals were placed in the centre square of the EPM always facing an open arm. An arm entry was counted when the rat’s entire body moved into an arm, and the centre square was considered a closed arm. Outcomes, including total distance travelled, entries to open and closed arms and duration of stay in open and closed arms were recorded by Ethovision™ tracking software (Noldus Information Technology, Leesburg, VA). Data were presented as average and sum pre- and post-injury time in open and closed arms and corresponding entries. Percentage change in closed arm duration and entries was also calculated [((post-closed arm duration – pre-closed arm duration)/pre-closed arm duration)
Paw Thermal Sensitivity
Paw thermal sensitivity assessment was performed following the post-rbTBI anxiety-like behaviour on the EPM after a 60-90-min period of rest in the home cage. Rats were positioned in a Plexiglas chamber on a glass platform and allowed to acclimate for 5-10 min. Then, a radiant heat light source (Model 336 Plantar Analgesia Meter, IITC Inc., Woodland Hills, CA) was positioned under the glass platform directly beneath the paw and aimed at the plantar surface until the rat exhibited a quick paw withdrawal reflex. A cut-off time of 10 s was used to avoid tissue damage. Each forepaw and hind paw were probed 3 times with a 3-min intertrial interval between each probe. The time to withdraw from the stimulus (withdrawal latency) was recorded by a built-in stopwatch. 40 Data are presented as average forepaw and hind paw thermal withdrawal latency.
Perfusion and Fixation of Brain Tissue
Approximately an hour after thermal sensitivity testing, each rat was deeply anesthetised (5% isoflurane) and transcardially perfused with cold phosphate-buffered saline (PBS) at pH 7.4, followed by cold 4% paraformaldehyde (4% PFA) in PBS. Following perfusion, the harvested brains were stored in 4% PFA in PBS for an additional 24-48 h. They were then cryoprotected by immersion in 15% sucrose followed by 30% sucrose in PBS. Forty-micron thick frozen serial sections encompassing areas of the prefrontal cortex (bregma 2.52 mm to 3.72 mm) were collected in 1 × PBS-filled multi-well plates and stored at 4°C until further processing.
Immunostaining and Glial Cell Quantification for Neuroinflammation
Representative brain sections were treated for antigen retrieval by incubation in citrate buffer (pH 6.0, C9999, Sigma Aldrich, Saint Louis, MO) at 90°C for 30 min and followed by incubation in 0.6% hydrogen peroxide (1 hr) to quench endogenous peroxidase activity. Then, they were incubated in a blocking buffer containing2% normal goat serum (Vector Laboratories, Burlingame, CA) and 2% bovine serum albumin (BSA) for an additional 1 h. Sets of sections were next incubated in the blocking solution containing anti-ionized calcium-binding adapter molecule 1 antibodies (Iba1, cat # 019–19741, Wako Chemicals, Richmond, VA; 1:20000) for detecting microglia/macrophages and in a solution with antibodies against the GFAP (cat # NE1015, Calbiochem, La Jolla, CA; 1:5000) to detect astrocytes. After overnight incubation, the sections were incubated in a solution with their respective biotinylated secondary antibodies (1:500; Vector Laboratories, Burlingame, CA), then subjected to Vectastain Elite ABC reagent exposure. Finally, the peroxidase activity was developed by incubation in 3,3’-diaminobenzidine and hydrogen peroxide solution. The sections were washed, dehydrated and coverslipped for microscopic observation (Nikon Eclipse Ni-U microscope), imaging, and quantification.
Microglial/macrophage and astrocyte cell quantification was performed using digital images (10 x images; Nikon DS-Fi2 digital camera) obtained from separate Iba1 and GFAP stained sections, respectively. Data from only five subjects were included in the blast group due to technical difficulties while sectioning for one of the subjects. A set of three sections for each stain per animal were assessed. Four digital images (20x) encompassing prefrontal cortex (PFC) areas from both hemispheres were obtained from each section (bregma 2.52 mm to 3.72 mm; ML-±1; DV 3–5.5). Forceps minor of the corpus callosum was used as a landmark to maintain the area of PFC consistent between images. The area medial to this and the midline of the section encompassing pre- and infra-limbic areas were used for imaging. Then the number of Iba1 and GFAP positive cells from each digital image was quantified using ImageJ by an independent investigator blinded to the test conditions and group assignments. First, each image was inverted, to ensure a better delineation of profiles of interest from their background. The total number of Iba1 and GFAP-positive cells were manually quantified with the aid of a multi-point tool in ImageJ and the average number of cells from the images collected per animal was analysed for group-wise differences.
Statistical Analyses
Surface righting reflex duration, outcomes from EPM (duration in open and closed arms, entries to open and closed arms and distance travelled [centimetres]), thermal withdrawal latency and Iba1 and GFAP positive cell numbers, were measured and compared for group-wise differences. These analyses employed an a priori hypothesis to assess qualitative differences between rbTBI and sham animals. A Day (3 days) by injury condition (2 conditions), repeated two-way analysis of variance (ANOVA) was performed. Then a corrected Fisher’s LSD post-hoc to assess differences in surface righting duration over the 3 days of sham and rbTBI procedures was performed. Other measures were evaluated by pair-wise comparisons using appropriate nonparametric methods (e.g., the Mann–Whitney test). This study employed a traditional significance P value threshold of <0.05 (e.g., 95% CI).
Results
rbTBI Increases the Surface Righting Reflex Duration
All animals in the rbTBI group showed prolonged surface righting duration following the injury. The surface righting duration was averaged over days for each animal and subjected to a Mann–Whitney analysis. Figure 2 illustrates the significantly higher surface righting duration (P = 0.001) in the rbTBI animals (193.1 ± 16.72 s) compared to sham animals (126.5.1 ± 4.85 s). A follow-up day by injury condition repeated two-way ANOVA revealed a significant main effect of rbTBI condition interaction between sham and rbTBI animals (F1, 10 = 14.65, P = 0.003). A Fishers’ LSD analysis that compensated for the number of comparisons (omnibus P = 0.05; per comparison P = 0.05/3 = 0.017) revealed that rbTBI animals were not significantly different (P = 0.059) on day 1. On days 2 and 3, rbTBI animals showed a significantly increased surface righting reflex duration in comparison to sham animals (P = 0.014 and P = 0.016, respectively; Supplementary Figure 1).
rbTBI Induces Prolonged Surface Righting Reflex Duration. Overall, animals in the rbTBI group showed a significantly higher surface righting duration (P = 0.001) compared to sham animals. Data were presented as mean ± SEM of 6 animals per group and were analysed by the Mann–Whitney test.

rbTBI Causes Anxiety-like Behaviour on the Elevated Plus Maze
Before injury or sham procedure, animals in both groups showed no apparent differences in preference for time spent (duration) in open arms or closed arms (Supplementary Figures 2A, B and C). However, post-injury, animals in the rbTBI group spent less time in the open arms and more time in closed arms. A decrease in percent change in open-arm duration and an increase in percent change in closed-arm duration compared to their pre-injury levels was observed but did not reach significance (Figure 3A; Supplementary Figure 2 D, E, and F). The preferential behaviour of spending more time in the closed arms by rbTBI animals post-injury was also shown in Table 1. Qualitatively, compared to their pre-injury closed arm duration, sham animals only showed a moderate +2.7% change in the sum of closed arm duration post-injury. On the other hand, rbTBI animals showed a greater +12.15% change in the sum of post-injury closed arm duration compared to their pre-injury levels.
rbTBI-related Anxiety-like Changes 30 Days Post-injury. rbTBI animals spent more time in closed arms than open arms. This is reflected by a decrease in total open arm duration and a corresponding increased percent change in closed arm duration although it has not reached significance (A; Mann–Whitney test). This preference for spending more time in the closed arms by rbTBI animals was further exemplified in the table that shows the proportion of closed arm duration by animals in both the groups and corresponding pre- to post-injury change that was higher in rbTBI animals compared to sham animals. rbTBI animals also made more entries to closed arms than open arms. rbTBI animals showed a higher preference for entering closed arms as reflected by a decrease in total open arm entries and a corresponding significant increase in the individual subject’s percent change in closed arm entries from baseline to post-injury testing (B; Mann–Whitney test).
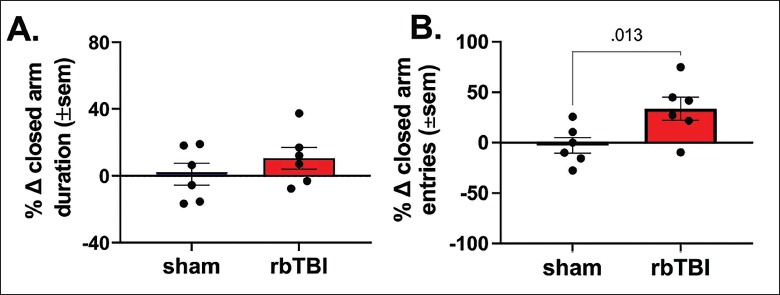
Proportion of Time Spent in Closed Arms By Animals in Both the Groups Pre- and Post-injury.
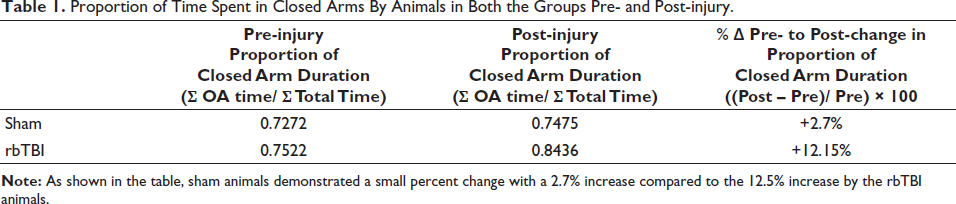
Before injury or sham procedure, animals in both groups showed no apparent differences in preference for entering open or closed arms (Supplementary Figures 3 A, B and C). However, post-injury, rbTBI animals showed a clear preference for entering closed arms as shown by a significantly increased percent change in closed arm entries (Figure 3B; P = 0.013). This is additionally exemplified by a preferentially higher number of entries to the closed arms by the rbTBI animals compared to sham animals post-injury (Supplementary Figures 3D,3E and 3F).
The distance travelled by the sham and rbTBI animals on the EPM showed no significant changes before and 30 days post procedures. A regression analysis between the average surface righting and the variables in the EPM (open and closed arm duration, open and closed arm entries) showed no significance.
rbTBI Increases Paw Thermal Sensitivity
rbTBI animals also showed a significantly (P = 0.032) shorter forepaw withdrawal latency than sham animals, suggesting increased forepaw sensitivity to noxious thermal exposure (Figure 4A). Decreases in hind paw thermal sensitivity following rbTBI were not at a significant level compared to sham controls (P = 0.090; Figure 4B). A regression analysis between the average forepaw and hindpaw) withdrawal with the average surface righting latency showed no significant relationship.
rbTBI-related Paw Thermal Sensitivity Changes. On average, rbTBI animals showed significantly (P = 0.032) lower forepaw withdrawal latency (i.e., quicker forepaw withdrawal) than sham animals 30 days post-injury (A). No significant difference (P = 0.090) was seen for hind paw withdrawal latency (B). The data were presented as the mean ± SEM of 6 animals per group. Data were analysed by the Mann–Whitney test.

rbTBI Increases Iba1 Reactive Microglia/Macrophage Number in the Prefrontal Cortex
Quantification of Iba1 reactive cells from digital images revealed a significantly higher count of positive cells (P = 0.026; Figure 5A) in PFC sections from the rbTBI group (Figure 5C) compared to the sham group (Figure 5B). A regression analysis between the microglial/macrophage number in the PFC and the average surface righting latency showed no significant relationship.
rbTBI-related Inflammation Is Marked By Increased Glial Counts. A significant increase in microglial/macrophage counts was observed (P = 0.026) in the prefrontal cortical region of sections from rbTBI animals compared to sham (A). Panels B and C show representative images of prefrontal cortical (bregma 2.52 to 3.72 mm; ML-±1; DV 3-5.5) Iba-1 positive microglial/macrophage staining from sham and rbTBI sections. A significant increase in GFAP-positive astrocytes was also observed (P = 0.015) in the prefrontal cortical region of sections from rbTBI compared to sham (D). Panels E and F show representative images of GFAP-positive astrocytes from sham and rbTBI sections. Scale bar = 100 µm. Data were presented as mean ± SEM of sections from 5 to 6 animals per group. Data were analysed by the Mann–Whitney test.

rbTBI Increases GFAP-reactive Astrocytes in PFC
Counting of GFAP-positive cells from digital images supported a significantly high number of GFAP-positive cells (P < 0.015; Figure 5D) in PFC sections from the rbTBI group (Figure 5F) compared to sham (Figure 5E). Although not quantified, astrocytic profiles were also noted in subcortical and white matter regions of the sections studied. A regression analysis between the GFAP-reactive cells and the average surface righting demonstrated a trending (P = 0.071) but ultimately a non-significant relationship between surface righting and astrocyte activity.
Discussion
Results from this initial study support the presence of prolonged surface righting duration at the time of injury, changes in behavioural surrogates of anxiety, forepaw thermal sensitivity and glial proliferation changes at 1 month in this model of rbTBI with no apparent changes in weight in both the animal groups. Our study replicates the deficit in righting response reported in female rbTBI-exposed mice 41 and suggests similarity in vulnerability between the sexes on this metric.
Although previous rbTBI studies that also used high-level blast pressures reported enhanced edema 23 and elevated glial reactivity26, 42 a few hours to days after rbTBI, they lacked anxiety-like behaviour data at 30 days. For instance, Schindler et al. 43 reported only an acute stress response marked by weight loss and elevated corticosterone levels, combined with traits of aversive behaviour 30 days post rbTBI. Meanwhile, Blaze et al. 44 reported mixed findings, with anxiety-like behaviour being observed in the open field assay but not in the elevated zero maze or light-dark box at 1-1.5 months post-rbTBI. Although the anxiety-like behaviour was reported to be diminished by 12-13 months, they indicated the persistence of anxiety-like behaviour in a subsection of animals on the elevated zero maze. Moreover, many reports on behavioural traits of anxiety or PTSD were in models at 6 to 12 months following rbTBI27, 28, 44, 45 but not a month after. For instance, Perez Garcia et al. 45 showed anxiety-like phenotypes in the EZM and light-dark box tests in animals exposed to low-level rbTBI at 1 week and 36-40 weeks post-injury, but not at 3-5 weeks. Support for the presence of behavioural characteristics of anxiety around a month after rbTBI as in this study comes from a recent study by Ravula et al. 46 who also reported anxiety-like behaviour at 25 days in their model of five successive low-level blast pressure exposures. Taken together, current literature indicates a temporal diversity in rbTBI-induced anxiety-like behaviour at earlier time points. This may be comparable to a level of non-uniform display of anxiety or PTSD-like symptoms in bTBI Veterans. 47
Many blast-exposed service members also report a cluster of musculoskeletal pain, concussion and mental health-related issues. 48 In our study, prominent forepaw thermal sensitivity in rbTBI animals 30 days following injury was observed. However, the lack of baseline data on this metric in our study limits our understanding of pre- and post-injury changes. Surprisingly, studies of pain-related behaviour in animal models of rbTBI are minimal. For example, Uddin et al. 49 showed no hind paw mechanical hyperalgesia or changes in gliosis following single bTBI. However, they reported persistent facial hypersensitivity to cold temperatures, combined with mechanical hyperalgesia and enhanced Rat Grimace Scale scores 4-5 weeks after a single bTBI (500 kPa) in awake rats. Whether or not the observed thermal sensitivity in our study is persistent and related to altered neuronal activation is not well established. While one study showed no marked changes in the spontaneous or sensory-evoked activity of posterior thalamic and spinal trigeminal nucleus neurons despite behavioural signs of pain, 49 others showed bTBI-induced neuronal activation 28 days post-injury as evidenced by manganese-enhanced magnetic resonance imaging. 50
Finally, we also show concurrent neuroinflammatory alterations as evidenced by an increase in glial cells 30 days post-rbTBI. However, the possibility of sustained glial proliferation and their putative phenotypic alterations extending to chronic conditions need to be studied. Furthermore, if these increased glial number also coincides with a corresponding increase in inflammatory mediators need to be verified. Glial proliferation changes have long been considered a marker and driver of bTBI pathology and ensuing comorbidities. Many recent rbTBI studies, despite reporting behavioural phenotypes of anxiety and PTSD,27–29, 43 provide no information on glial or inflammatory markers. Sosa et al. 51 reported chronic microglial activation related to focal haemorrhagic tears at 4 months, but not at 1.5 months in their low-level rbTBI model. The possible occurrence of such lesions in rbTBI brain tissue in our study cannot be disregarded, as high-level blast exposure was also shown to be associated with haemorrhagic lesions. 52 Given reports of disrupted blood barrier following blast pressure, 53 the increased microglia/macrophage number observed in this study may also be associated with such disruption and facilitation of peripheral immune cell infiltration. On the other hand, decreased GFAP and vascular pathology in the form of reduced vascular coverage by GFAP and degeneration of astrocyte end-feet 6 weeks after rbTBI has been noted. 54 A decrease in astrocytic end feet coverage was also described 24 h after a single blast TBI. 55 Such vascular changes may have implications for altered cerebral blood flow and can impact inflammation, 56 leading to varied chronic conditions.
Evidence of inflammation also comes from analysis of serum from service members routinely exposed to rbTBI that showed dysregulated gene expression associated with chronic inflammation and immune response. 57 Such inflammation-oriented alterations may ultimately contribute to or exacerbate the enduring neurological symptoms following rbTBI. For that matter, the association of neuroinflammatory cytokines with poor psychological outcomes in mTBI patients has been reviewed. 58 Additionally, psychological factors, such as anxiety and depression, are postulated to be associated with enhanced sensitivity to pain 59 and a role for microglia in pain and anxiety has been proposed. 59
The pressures used in our study were described as high level while descriptions of blast pressures used by investigators vary. Elder et al. described blast pressures of around ~11 psi in their studies as low-level,27, 28, 44, 45 while pressures of 14.6 psi were described as mild. 24 Some referred to pressures in the range of 20 psi as high level,23, 26 while others referred to 19.91 psi as pressures in keeping with mTBI. 60 Other investigators described air blast pressures of 30 and 50 psi as sub-concussive and concussive. 61
The modest behavioural results observed in our study may be related to limitations arising from the sample size. The lack of assessment of neuronal injury changes as part of this study can also be viewed as another limitation. Additionally, the lack of female rats could be considered another limitation of our study, with many previous observations,27, 28, 44, 45 including ours, being made exclusively in male rats. Recently, Baskin et al. 41 reported a potential dimorphic rbTBI-induced behaviour in which both male and female animals showed anxiety-like behaviour at 48 h; however, by 1 month, such changes were demonstrated in male animals alone. Similarly, only male rats have been shown to exhibit anxiety-like behaviour at both 2 and 7 days after rbTBI. 55 The lack of these behavioural changes, along with a well-maintained blood-brain barrier, in female rats was described as a potential sign of resilience. 55
Conclusion
The 30-day post-rbTBI period may be characterised by a combination of behavioural and glial changes that need to be considered in the evaluation and management of blast trauma-related pathology. Future studies including females and assessing chronic behavioural changes with concurrent glial alterations post-rbTBI are warranted.
Footnotes
Authors’ Contribution
SK and SAP contributed to the conception and design of the study. SK and NS performed the experiments.
SK and CJD performed data analysis.
SK, CJD and AG contributed to the preparation of the manuscript.
SK, CJD, AG, KB, CB, AC and SAP interpreted the results.
All authors contributed to the revision of the manuscript, and read and approved the submitted version.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This research work was supported by National Institutes of Health grants R01-DA042057 (Perrine); R21-DA052657 (Perrine); and Veterans Administration Merit Award I01-RX-002252-03 (Perrine).
Statement of Ethics
Datasets are available upon request. The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation upon reasonable request.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.