
Abstract
Osteonecrosis of the femoral head (ONFH) is a leading cause of mobility impairment which may lead to a total hip replacement. Recent studies have found tendon derived stem cells (TDSCs) might be an ideal cell source for musculoskeletal tissue regeneration. And our previous study has shown Sox11 could promote osteogenesis of bone marrow-derived MSCs. However, the effect of TDSCs or Sox11 over-expressing TDSCs (TDSCs-Sox11) on bone regeneration in ONFH has not been investigated. In the present study, TDSCs were infected with AAV carrying Sox11 or empty vector. We showed that Sox11 could promote the proliferation and osteogenic differentiation of TDSCs, as well as angiogenesis in vitro. The western blot analysis showed that Sox11 could activate the PI3K/Akt signaling pathway to promote osteogenesis of TDSCs. Finally, using a rabbit model of hormone-induced ONFH, our result demonstrated that local administration of TDSCs or TDSCs overexpressing Sox11 could accelerate bone regeneration in necrotic femoral heads, and TDSCs overexpressing Sox11 showed better effects. TDSCs over-expressing Sox11 might be a promising cell source for stem cell therapy to promote bone regeneration, such as ONFH, fracture, bone defect, and so on.
Introduction
Osteonecrosis of the femoral head (ONFH) is a painful disease that may lead to a total hip replacement 1 , while the etiology and pathology are not completely clarified 2 . Administration of corticosteroids is one of the most important causes leading to ONFH. Five to twenty-five percent of patients with atraumatic ONFH are caused by steroid administration 3 . The pathophysiology of steroid-associated ONFH (SAON) remains controversial. Multiple factors are involved in the process of SAON, including lipid metabolism, oxidative injury, the impaired balance between bone rebuilding and resorption and vascular impairment, and so on 4 .
Steroids can inhibit the osteogenic differentiation of bone marrow stem cells (BMSCs) while inducing differentiation of BMSCs into adipocytes, which is an important pathophysiological mechanism of SAON 5 . The abnormality of the bone marrow stem cell pool has been verified in patients with SAON clinically 6,7 , as well as in animal models 8,9 . BMSCs have been well known as promising cell sources for therapeutic use in regenerative medicine and tissue engineering. They are characterized by the multiple-lineage differentiation potentials, as well as the ability to be easily expanded ex vivo while retaining their original lineage differentiation commitment 10 –13 . Recent studies have found tendon derived stem cells (TDSCs) might be a better cell source than other cells for musculoskeletal tissue regeneration, in terms of multilineage differentiation and proliferation 14 . In addition, TDSCs also showed a better capacity to directly contribute to de novo formation in critical-sized bone defect 15 .
Sox11 is a clade of the Sox C group of the Sox (Sry-related HMG box) transcription factors. The Sox proteins are characterized by HMG box which is a highly conserved DNA binding domain. The previous study has found that Sox11 knockout mice show craniofacial and skeletal malformations, asplenia, and hypoplasia of the lung, stomach and pancreas 16 , implying that Sox11 may be associated with bone development and bone diseases. Recently, Sox11 has been identified as one of the MSC-characteristic transcription factors involved in stemness regulation by DNA microarray analysis 17 . And Knocking down Sox11 suppressed MSCs’ self-renewal capacities, reduced their osteogenic and adipogenic differentiation potential. Furthermore, our study also demonstrated that Sox11 overexpression could promote the multi-lineage differentiation potentials of MSCs, and could be used to accelerate fracture healing as well as cartilage regeneration 18,19 . However, the effect of TDSCs or TDSCs with Sox11 overexpression on bone regeneration during the progress of ONFH has not been investigated. With the potential capacities of TDSCs and the characteristic effects of Sox11, we hypothesized that the transfer of Sox11 to TDSCs using AAV might improve the repair of osteonecrosis in the rabbit model of ONFH. AAV is one of the safest viral vectors applied in clinical trials, which is supported by recent studies showing no cellular immune responses or tissue disturbance 20 –22 .
In the present study, we used recombinant AAV carrying the Sox11 gene to infect TDSCs in vitro. And mainly investigated the effects of Sox11 over-expressing on osteogenesis and angiogenesis, as well as the application of Sox11 over-expressing TDSCs to promote bone regeneration in a rabbit model of ONFH.
Materials and Methods
Isolation and Culture of Rabbit TDSCs
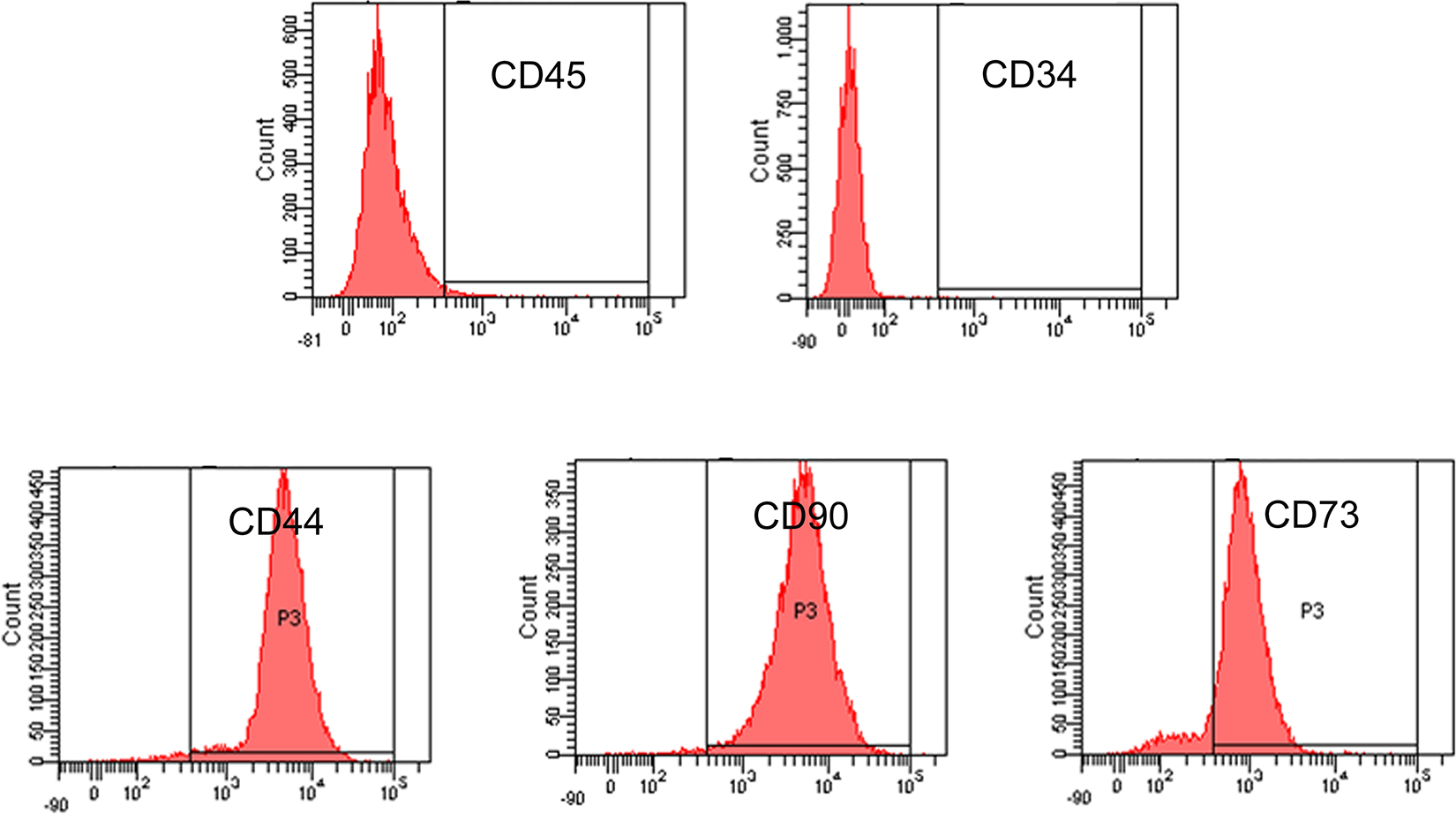
TDSCs were isolated from the rabbit patellar tendon by digesting with type I collagenase (3 mg/mL; Sigma-Aldrich, St. Louis, MO, USA) as previously described 23 . The tissue enzyme solution was filtered with a 100-mm cell strainer. Single cells were collected from the filtrate, washed twice with PBS, and plated into a 100-mm culture dish at 500 cells/cm2. TDSCs were grown in LG-DMEM (Invitrogen GIBCO, New York, USA) with 10% fetal bovine serum (Invitrogen GIBCO, New York, USA) and antibiotics (penicillin 100 U/mL, streptomycin 100 g/mL) at 37°C with 5% CO2. The next day, the cells were washed with PBS to remove non-adherent cells and trypsinized when they reached 70–80% confluency. The surface antigens of TDSCs were analyzed by flow cytometry with antibodies against CD34, CD44, CD45, CD73 and CD90. Cells of passage 2-8 were used in this study.
Gene Cloning and AAV Production
The gene encoding rabbit Sox11 was amplified and cloned into an AAV viral vector by in vitro recombination. Viruses were prepared by amplification in 293 cells. The virus suspension was purified by three consecutive CsCl gradients. The viral titers were estimated by standard plaque assay. For infection of TDSCs, 5 × 105 cells were seeded into 10 cm dishes and incubated with AAV in the incubator for 12 h.
Cell Proliferation Assay
TDSCs infected with AAV-Sox11 or empty vector were plated at 2,000 cells/well in the complete culture medium in a 96-well plate and incubated at 37°C, 5% CO2. At day 3, cell proliferation was checked using the BrdU assay kit (Roche Applied Science, Indianapolis, IN) according to the manufacturer’s instruction. The absorbance at 450 nm was measured and reported.
Osteogenic Differentiation
TDSCs or TDSCs-Sox11 were plated at 4 × 103 cells/cm2 in a 12-well plate and cultured in the basal medium until the cells reached confluence, respectively. They were then incubated in osteogenic induction medium (OIM), which is basal medium supplemented with 50 μM ascorbic acid (Sigma-Aldrich, USA), 1 nM dexamethasone (Sigma-Aldrich, USA) and 20 mM β-glycerolphosphate (Sigma-Aldrich, USA). At day 10, the mineralization was stained with 0.5% Alizarin red S (pH 4.1, Sigma-Aldrich, USA) for 5 min.
Quantitative Real-Time RT-PCR (qRT-PCR)
The cells were harvested and homogenized for RNA extraction with RNeasy mini kit (Qiagen, Hilden, Germany). The mRNA was reverse-transcribed to cDNA by the PrimeScript First Strand cDNA Synthesis Kit (Takara, Dalian, China). Each sample was diluted and 5 µl of total cDNA was used for amplification in reaction mixture containing Platinum SYBR Green, qPCR SuperMix-UDG ready-to-use reaction cocktail and specific primers. The expression of the target gene was normalized to that of β-actin gene. Relative gene expression was calculated with the 2-
△CT formula. The sequences of the primers were shown in
In Vitro Tube Formation Assay
The twelve-well plates were coated with Matrigel (Becton Dickinson, USA). Then TDSCs or TDSCs-Sox11 (5000 cells/well) and HUVECs (1 × 104 cells/well) were cocultured and seeded on the 12-well plates, respectively. TDSCs (1 × 104 cells/well) alone served as blank control. DMEM and F12 basal medium (Thermo Fisher Scientific, USA) containing 2% FBS (Thermo Fisher Scientific, USA) was used. Plates were incubated at 37°C, 5% CO2 for 8 h. Bright-field images were captured for analysis of network formation.
Western Blot
Equal proteins were loaded onto 10% Tris/glycine gels for electrophoresis and then transferred to a PVDF membrane (Millipore, USA). All the antibodies used were purchased from Beijing Biosynthesis Biotechnology Co., LTD, Beijing, China. Anti-Sox11 (bs-1025 R, 1:1000), anti-VEGF (bs-1313 R, 1:1000), anti-Runx2 (bs-1134 R, 1:1000), anti-p-JNK (bs-17591 R, 1:1000), anti-JNK (bs-10562 R, 1:1000) or anti-β-actin (bs-0061 R, 1:1000) antibodies were used in this study. After washing in TBST, the membrane was incubated with horseradish peroxidase-linked secondary antibodies (anti-mouse or anti-goat) for 1 h at room temperature. Following TBST washes, protein was detected with the enhanced chemiluminescence blotting reagents (Amersham Biosciences, USA) according to the manufacturer’s instruction.
ONFH Model and Cell Transplantation
The experimental design was approved and performed following the Animal Care and Use Committee guidelines of PLA General Hospital. Eighteen 12-weeks old male New Zealand rabbits were used. Animals were anesthetized with an intravenous injection of pentobarbitone sodium. As previously described, methylprednisolone acetate (MPSL, 20 mg/kg body weight) was used to establish the steroid-induced osteonecrosis model, the physiologic saline (1ml/kg body weight) was used as control 24,25 . Four weeks after the injection, the rabbits were anesthetized and core decompression of the femoral head was performed on the right hip. The femoral head was punctured once. After then, 1 × 106 TDSCs or TDSCs-Sox11 in 100 µl PBS were delivered into the necrotic femoral head. The control group just received 100 µl PBS.
Micro-CT Examination
For microCT analysis, the femurs were scanned by vivaCT 40 (Scanco Medical AG, Bassersdorf, Switzerland) using our previously established protocols 26 . Transect scanning of the entire hip joint was performed, including the upper and lower edges. The scanning parameters were 100 kV and 220 mA.
Histology and Immunohistochemistry
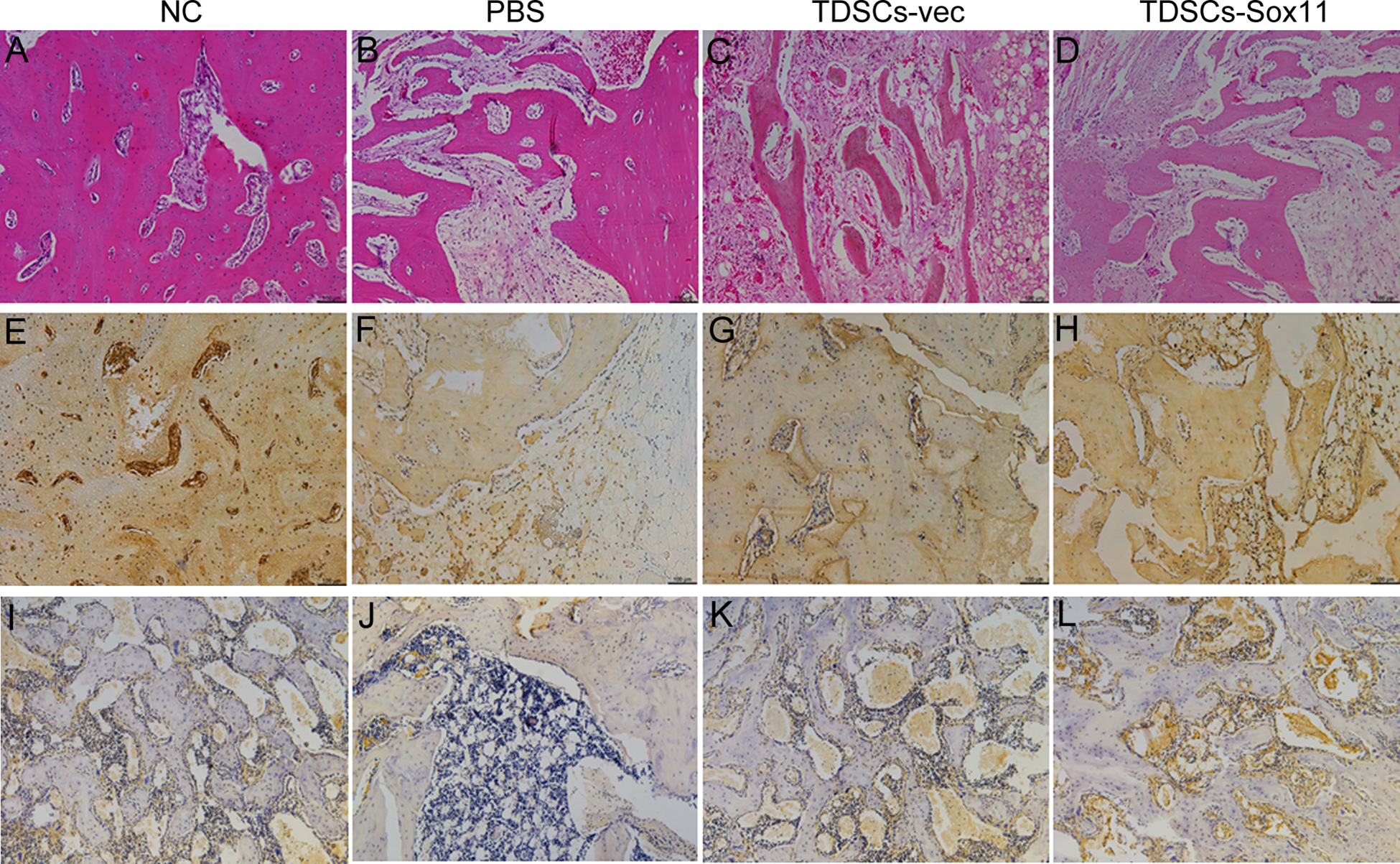
Histological staining was performed as previously described 27 . Generally, the femoral heads were collected and washed in PBS, fixed in 4% paraformaldehyde, decalcified, dehydrated and embedded in paraffin. Sections were cut at a thickness of 5µm and stained with hematoxylin and eosin (H&E) after deparaffination. As for immunohistochemical (IHC) staining, endogenous peroxidase activity was quenched with 3% hydrogen peroxide at room temperature. Antigen retrieval was then performed with citrate buffer at 80°C for 10 minutes. Primary antibody against OCN (1:100; Beijing Biosynthesis Biotechnology Co., China) was used. The horseradish peroxidase (HRP)-conjugated secondary Goat anti-mouse antibody (1:100, Santa Cruz) was then added for an hour, followed by 3, 3’-diaminobenzidine tetrahydrochloride (DAKO, Glostrup, Denmark) for signal detection of OCN. Then the sections were rinsed, counterstained in hematoxylin, dehydrated, and mounted with p-xylene-bis-pyridinium bromide (DPX) permount (Sigma-Aldrich, St. Louis, MO, USA). Primary antibody was replaced with blocking solution in the negative controls. All incubation times and conditions were strictly controlled.
Data Analysis
Data were presented as mean ± SD. All the data analysis was done using SPSS (version 16.0; SPSS Inc). Two-sided t-test was used for comparison of mean values with P < 0.05 as considered statistical significance.
Results
Characterization of Rabbit TDSCs with Flow Cytometry
First, the rabbit TDSCs were isolated and cultured. At passage 2, the surface antigens of TDSCs were analyzed by flow cytometry. The result showed that rabbit TDSCs were negative for CD45 and CD34, while positive for CD44, CD90, and CD73 (Fig. 1), which was similar to the typical markers of mesenchymal stem cells as defined by the International Society for Cellular Therapy standard criteria.
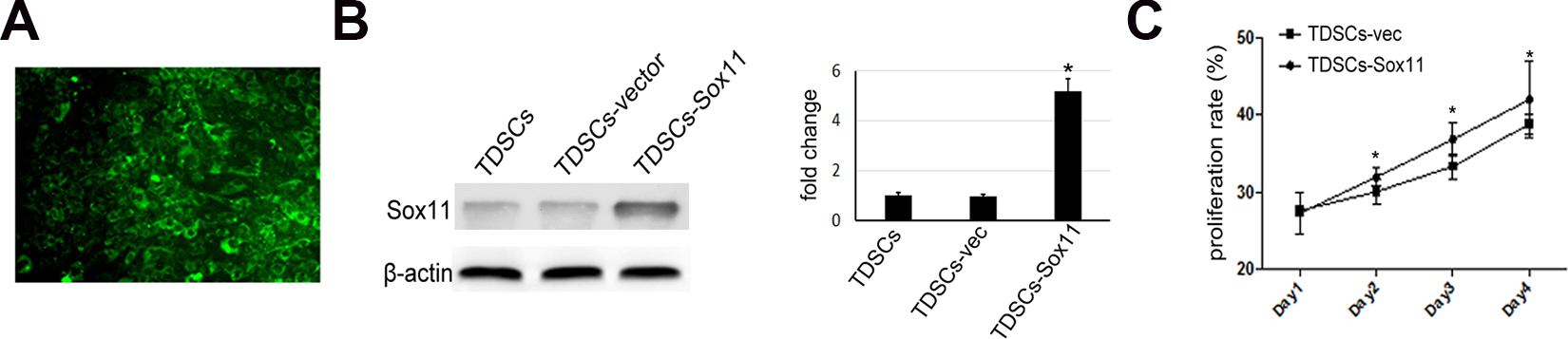
Sox11 Overexpression Increased Proliferation of TDSCs
In order to investigate the effect of TDSCs and Sox11-modified TDSCs on bone regeneration in a rabbit model of osteonecrosis, TDSCs were infected with recombinant AAV virus ( Fig. 2A ). The transfection efficiency was above 80% as estimated by the GFP signal visualized under fluorescent microscopy. Sox11 expression in TDSCs was confirmed at the protein level by western blot analysis ( Fig. 2B ). Cell proliferation analysis demonstrated that Sox11 overexpression could significantly promote the proliferation rate of TDSCs ( Fig. 2C ).
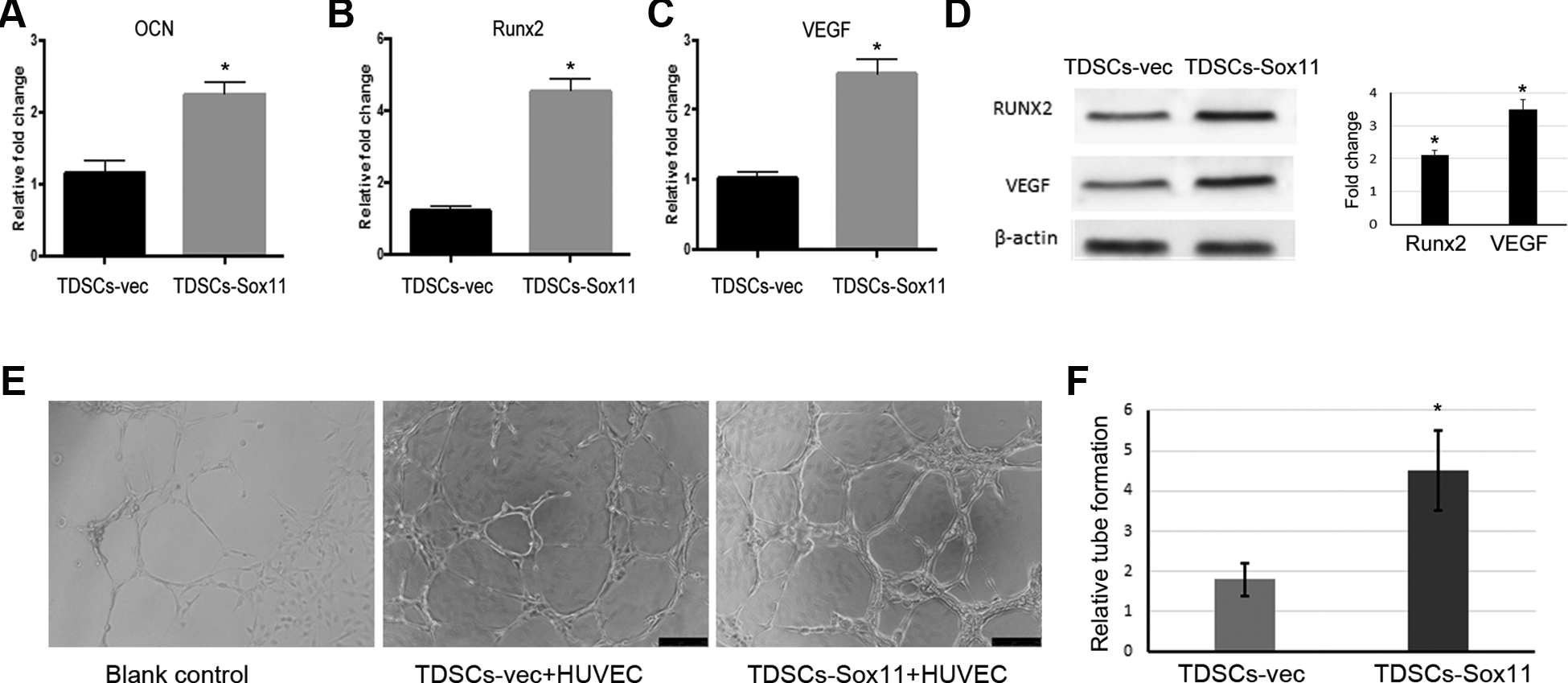
Sox11 Overexpression Promoted Osteogenesis and Angiogenesis
Next, we checked the influence of Sox11 on osteogenic differentiation and angiogenesis in TDSCs or TDSCs and HUVEC cells in vitro. It was found that Sox11 gene modification could increase the expression of osteogenic differentiation-related genes including OCN and Runx2, as well as angiogenesis-related genes such as VEGF
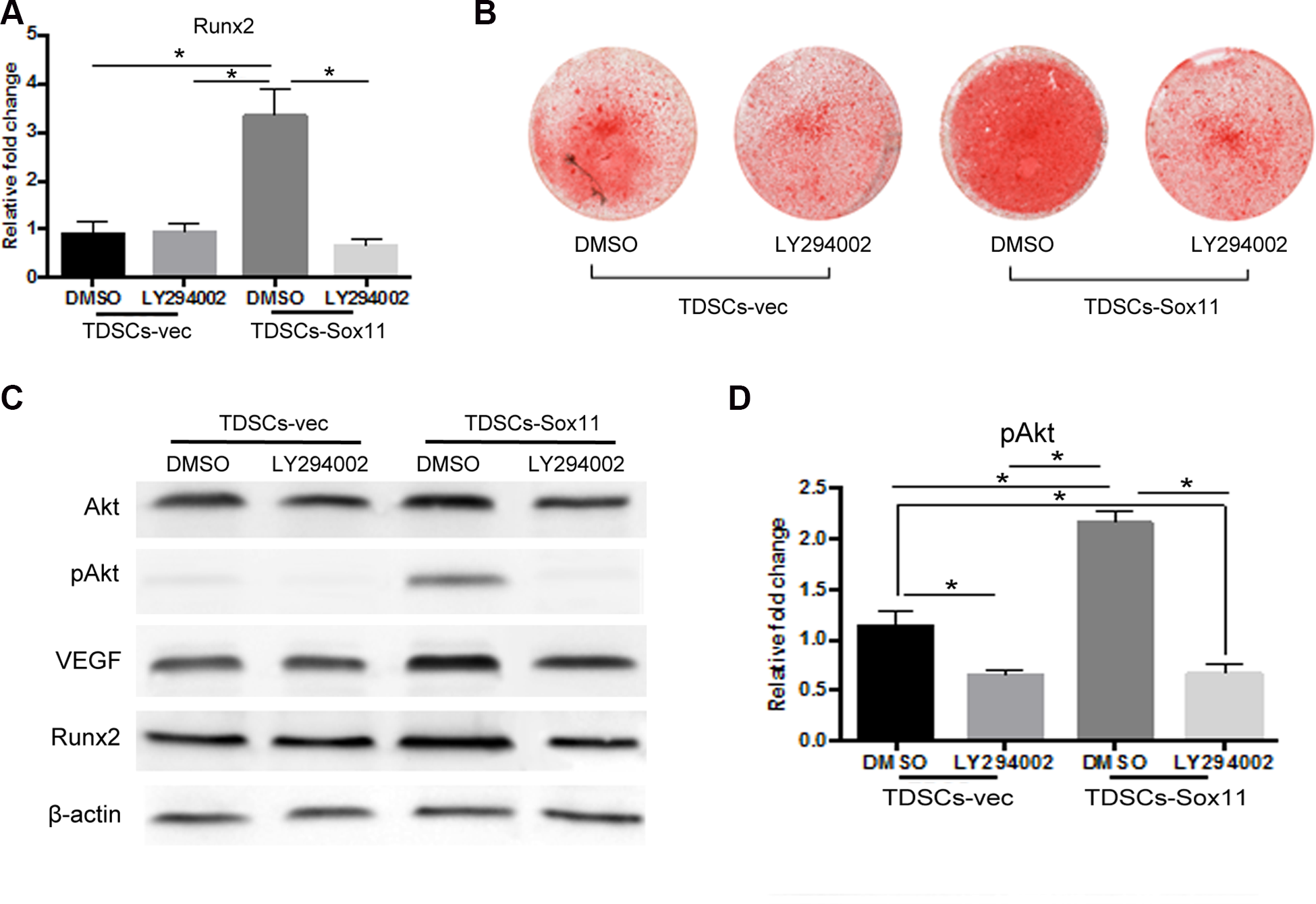
Sox11 overexpression Enhanced Osteogenesis via PI3K/Akt Signaling Pathway
Furthermore, to clarify the underlying mechanism, TDSCs transduced with Sox11 or empty vector were treated with LY294002 which was an inhibitor of PI3K/Akt signaling pathway. It was found that the promoting effect of Sox11 overexpression on Runx2 expression in TDSCs was blocked after treated with PI3 K inhibitor for 3 days
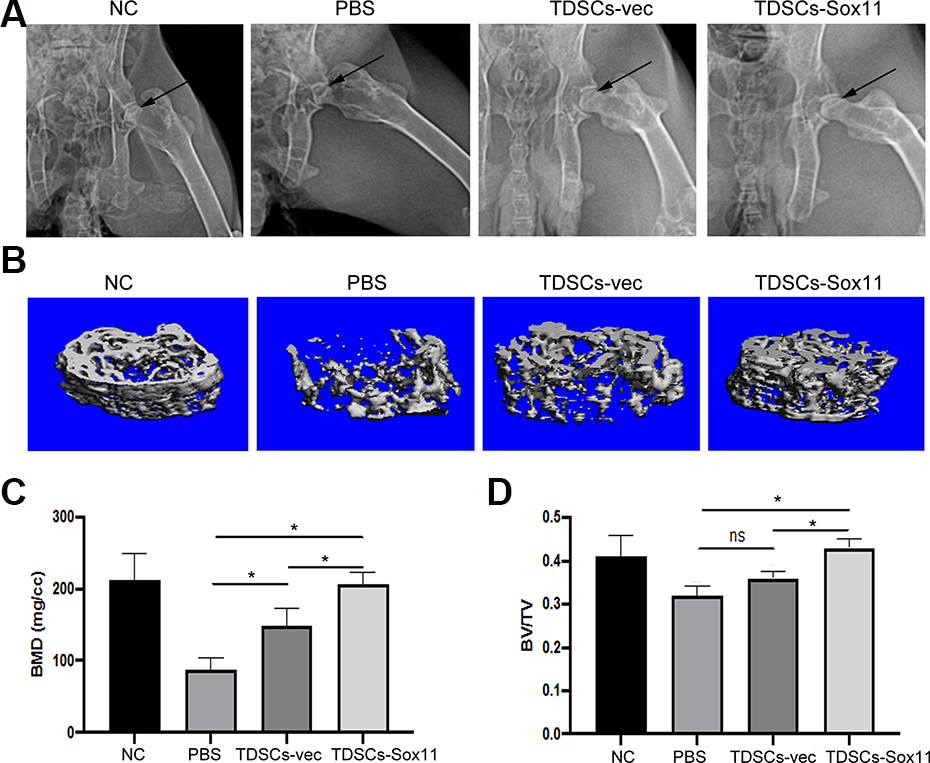
TDSCs Overexpressing Sox11 Enhanced bone Regeneration in the ONFH Model
To determine whether transplantation of TDSCs or TDSCs-Sox11 could enhance bone regeneration in a rabbit model of ONFH, the cells were delivered into the bone tunnel using a pipette after the core decompression procedure. The microCT analysis result showed that the femoral heads of the TDSCs and TDSCs-Sox11 groups had increased BMD and BV/TV compared with that of the PBS group
Discussion
In the present study, we showed that Sox11 overexpression mediated by AAV could promote the proliferation and osteogenic differentiation of TDSCs, as well as tube formation in vitro. The underlying mechanism is that Sox11 could activate the PI3K/Akt signaling pathway to promote the osteogenesis of TDSCs. Finally, using a rabbit model of hormone-induced ONFH, our result demonstrated that local administration of TDSCs overexpressing Sox11 could accelerate bone regeneration in necrotic femoral heads, as shown by the microCT and histological analysis.
MSCs have been considered as one of the most promising stem cells for cyto-therapy to treat many diseases, including fractures, cartilage defects, heart failure, and so on. The autologous bone-marrow mononuclear cells or MSCs have already been used to treat patients with the Association Research Circulation Osseous stage-I or II necrosis of the femoral head 28,29 . It is concluded that the intervention is safe and effective in delaying or avoiding femoral head collapse. Recent studies have found tendon-derived stem cells (TDSCs) might be a better cell source than BMSCs for musculoskeletal tissue regeneration, they showed better capacity to directly contribute to de novo formation in critical-sized bone defect 15 . Our previous study has also shown TDSCs could be used for tendon repair and regeneration 30 . However, the effect of TDSCs transplantation for ONFH treatment has not been studied.
ONFH is a leading cause of mobility impairment. Long-term administration of GCs (Glucocorticosteroids) is considered as one of the predominant factors resulting in ONFH 31 –33 . At present, no effective treatments have been found for ONFH 34,35 . Therefore, there is an urgent need for alternative interventions to treat ONFH and reduce patient morbidity. Although the etiology and pathology of ONFH are not completely clarified, impaired angiogenesis and osteogenesis are the two important factors involved in the process of ONFH 36 –39 . Several angiogenic and growth factors have shown promising effects on bone formation and regeneration in ONFH, including VEGF and FGF 40 –42 . In the present study, we used AAV to deliver Sox11 into MSCs. AAVs have been used in clinical studies with high efficacy and safety 43 . Recently, an intravenously dosed AAV vector delivering the SMN1 gene, Zolgensma, has been approved by FDA and successfully used in children with spinal muscular atrophy 44 . Our results showed that transplantation of Sox11-transgenic MSCs is a promising method for the treatment of ONFH. We found Sox11 could promote osteogenesis in TDSCs by activating the PI3K/Akt signaling pathway. By using the PI3K-specific inhibitor LY294002, we showed that osteogenic differentiation of TDSCs was significantly inhibited, as well as the expression levels of some specific markers such as Runx2. The PI3K/AKT pathway is involved in mediating many important cellular processes, including cell proliferation, differentiation, apoptosis, and so on. Recent studies have also found that PI3K/AKT can promote the proliferation of MSCs and inhibit their apoptosis 45,46 . Xue et al had found that Hif-1α overexpression in MSCs could improve cell viability and suppress apoptosis, in which the PI3K/AKT signaling pathway is implicated 46 . In our study, we proved that PI3K/AKT signaling pathway was also involved in the osteogenic differentiation of MSCs.
In addition, we also proved that Sox11 modified TDSCs could promote angiogenesis using tube formation assay with TDSCs and HUVEC co-culture. Sox11 has been demonstrated to be involved in neural development and organogenesis 47,48 . Some studies also showed that Sox11 was one of the most frequently upregulated genes in various cancers, including mantle cell lymphoma, epithelial ovarian cancer, breast cancer, and gastrointestinal tumors 49 –52 . The promotion of angiogenesis may be accounted for its role in tumors, as it has been shown to promote tumor angiogenesis through transcriptional regulation of PDGFA in mantle cell lymphoma 53 .
The results from histological and Micro-CT analysis clearly demonstrated that administration of TDSCs-Sox11 could restore the microstructure of the femoral head. The BMD and BV/TV of the femoral head were significantly increased by TDSCs-Sox11. And the rate of empty lacunae in ONFH was also decreased by TDSCs-Sox11. Taken together, our findings demonstrated that Sox11 could enhance angiogenesis, as well as osteogenesis via PI3K/Akt signaling pathway, and local administration of TDSCs overexpressing Sox11 could accelerate bone regeneration in necrotic femoral heads, and hopefully, the TDSCs overexpressing Sox11 might be clinically used to treat ONFH.
Supplemental Material
Supplemental Material, sj-docx-1-cll-10.1177_09636897211053870 - Sox11 Modified Tendon-Derived Stem Cells Promote the Repair of Osteonecrosis of Femoral Head
Supplemental Material, sj-docx-1-cll-10.1177_09636897211053870 for Sox11 Modified Tendon-Derived Stem Cells Promote the Repair of Osteonecrosis of Femoral Head by Ming Ni, Weiwei Sun, Yucong Li, Lingli Ding, Weiping Lin, Haiwen Peng, Qingyuan Zheng, Jingyang Sun, Juncheng Li, Hao Liu, Yi Yang, Liangliang Xu and Guoqiang Zhang in Cell Transplantation
Footnotes
Acknowledgments
The authors would like to thank Dr. Jing Zhang for the assistance of manuscript preparation and language editing. The work was supported by grants from the Beijing Municipal Science & Technology Commission (7192194), as well as National Natural Science Foundation of China (81472094).
Authors Note
Ming Ni and Guoqiang Zhang is also affiliated from Department of Orthopedics, the Fourth Medical Center, Chinese PLA General Hospital, Beijing, P.R. China.
Ethical approval
The study has been approved by the local ethics board of the Chinese People’s Liberation Army (PLA) General Hospital (Approval number S-2020-325).
Statement of Human and Animal Rights
All procedures in this study were conducted following the ‘Guide for the Care and Use of Laboratory Animals’ published by the National Academy of Science of the USA. The study has been carried out following the Animal Care and Use Committee guidelines of the Chinese Medical Association. And no human subjects were involved in this study.
Statement of Informed Consent
There are no subject in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.